
Abstract
Abstract
Effects of size and concentration of aquatic particles collected from Lake Poyang, as well as initial algal cell density and organic matter concentration, on flocculation of Microcystis aeruginosa (cyanobacteria) and Nitzschia (diatom) by aquatic particles were investigated in the laboratory. Results showed that flocculation efficiency of cyanobacteria by particles was higher when compared to diatoms. Flocculation efficiency of algae increased with decrease of particle size. When particle concentration increased from 0.02 g/L to 1.28 g/L, flocculation efficiencies of M. aeruginosa and Nitzschia increased from 19.3% to 43.7% and from 2.95% to 29.0%, respectively. Moreover, the flocculation efficiency increased with an increase of initial algal cell density, and efficiency of M. aeruginosa was two to three times higher than Nitzschia. Organic matter (humic acid) in the water had a positive effect on flocculation efficiency of M. aeruginosa, and efficiency increased from 17.6% to 23.4% when humic acid concentration increased from 0 mg/L to 8 mg/L. An increase in flocculation efficiency of Nitzschia from 11.8% to 16.3% was observed with an increase in humic acid concentration from 0 mg/L to 4 mg/L. It was concluded that in natural water bodies, flocculation efficiency of algae by the aquatic particles was highly dependent on characteristics of aquatic particles and algae such as size, extracellular polysaccharide contents on algal cell surfaces, algal cell density, and particle concentration, as well as organic matter concentration in the water. Aquatic particles can attenuate the dominance of cyanobacteria and promote dominance of diatoms, consequentially reducing cyanobacteria bloom risks.
Introduction
E
Lake Poyang is a typical Yangtze-connected lake in China, with a strong hydrodynamic condition. The evolution mechanism of cyanobacteria and diatoms in Lake Poyang is different from Taihu and Chaohu. Although total nitrogen (TN) and total phosphorus (TP) concentrations of Lake Poyang in 2016 (TN 1.45 mg/L, TP 0.07 mg/L) had approached the level in Taihu (TN 1.74 mg/L, TP 0.06 mg/L) and Chaohu (TN 1.4 mg/L, TP 0.07 mg/L) (Cai and Kong, 2013; Huang et al., 2014), the chlorophyll concentration was still low (only 5.2 μg/L), and the phytoplankton community was still dominated by diatoms. Some researchers suggested that this may be related to the transparency and turbidity (Wu et al., 2013, 2014). But they have analyzed the impact of particles on algae from the perspective of light. The impact of particles on the algae from the perspective of flocculation was not considered.
Aquatic particles affect the growth of algae mainly through light blocking and flocculation of algal cells (Bilotta and Brazier, 2008). At present, the studies on aquatic particles influencing the light transmission are far more than those on the effects of the particles on the flocculation of algae. Aquatic particles usually refer to small particles less than 63 μm (Waters, 1995). It mainly contains minerals, clays, organic particles, inorganic particles surrounded by organics, and biological debris (Lu and Allen, 2006). The viscous components of aquatic particles are mainly minerals, clays, and clay minerals which are usually smaller than 2 μm (Walling and Moorehead, 1989). They have the flocculation effect on the algae. The content of the viscous components in Lake Poyang is about 35.95–85.76% (Dong et al., 2011). These viscous components may have different flocculation effects on different algae.
Flocculation mechanism of modified particles on algae, especially cyanobacteria, was studied (Pan et al., 2006a, 2006b; Zou et al., 2006; Pan et al., 2011; Tang et al., 2011). Ionic strength, pH, organic content, cell concentration, and algal growth phase had different effects on the flocculation efficiency of Microcystis cells using modified particles. Besides, the viscosity and other properties such as the gravity of the algal cells varied at different growth stages, which could affect the affinity between the cells and the clay particles (Chen et al., 2004). Nevertheless, the properties of natural aquatic particles and the modified particles are different. Moreover, previous studies mostly focused on the effects of the modified particles on the flocculation of cyanobacteria. Studies on the flocculation effects of natural aquatic particles on algae and the effect of natural aquatic particles on the evolution of cyanobacteria and diatoms were lacking. In our study, several factors such as the size and concentration of the aquatic particles and the organic matter content in the algal particle mixture were selected to investigate their effects on the flocculation of cyanobacteria (Microcystis aeruginosa) and diatom (Nitzschia) using aquatic particles from Lake Poyang. The flocculating results may reveal the reason that diatoms are still the dominant species in Lake Poyang although nutrients were high compared with other similar lakes.
Materials and Methods
Algae species
M. aeruginosa (FACHB 1328) and Nitzschia (FACHB 510) were obtained from the laboratory of Institute of Hydrobiology, Chinese Academy of Sciences. They were incubated in sterilized 10 L glass flasks containing 9 L aqueous medium at 25°C ± 1°C under fluorescent light (3000 l × , 12-h light/12-h dark cycle). The algae were expanding cultured for about 1 week to reach the exponential phase. The cultivation media were M11 for M. aeruginosa and M11 plus 100 mg/L Na2SiO3·9H2O for Nitzschia. M11 medium was composed of 100 mg/L NaNO3, 10 mg/L K2HPO4, 75 mg/L MgSO4·7H2O, 40 mg/L CaCl2·2H2O, 20 mg/L Na2CO3, 6 mg/L ferric citrate, and 1 mg/L Na2EDTA ·2H2O.
Collection and pretreatment of aquatic particles
Aquatic particles were sieved from the sediments collected from Lake Poyang (E 116°32′37′′, N 29°09′55′′). The sediment samples were kept cold and transported to the laboratory. They were dried by natural wind and crushed and then sieved through a 300 mesh screen (63 μm). Finally, the aquatic particles were used to prepare a 5 g/L suspension with water.
Flocculation experiment
Flocculation experiments were conducted in 800 mL glass beakers stirred with a six-head stirrer (MY 3000-6; Qianjiang Meiyu instrument Co., Ltd., China). Five hundred milliliters of algae culture (M. aeruginosa in M11 or Nitzschia in M11 + 100 mg/L Na2SiO3·9H2O) was transferred into the 800 mL beaker, stirred at 21 s−1 [equivalent to 0.4 m/s velocity of Lake Poyang (Du et al., 2015), calculation formula was shown in Eq. (1)] for 30 min. Different volumes of the particle suspensions were added to the algae culture rapidly at the beginning of the stirring. The control culture was run without adding any particle suspensions. The initial algal cell density was controlled at 106 cells/mL (equivalent to the concentration of algae blooms occurrence), and the pH of the medium was adjusted to 8.0 with NaOH and/or HCl solution. After a subsequent sedimentation of 30 min, the supernatants were collected from 2 cm below the water surface in the experimental beaker. The algal cell concentrations in the supernatants were analyzed. The size and concentration of the aquatic particles, cell concentration, pH and organic matter of algal particle in the algal particle mixture were separately analyzed. The concentration of particulate matter in Lake Poyang was 0.016–0.15 g/L, with a mean value of 0.06 g/L (Cao et al., 2016). The pH was 7.0–8.0, and the chemical oxygen demand (COD) was between 9 mg/L and 16 mg/L. A measure of 0 mg/L–32 mg/L humid acid was equivalent to 0–22 mg/L CODCr (Pan et al., 2006b). Therefore, the detailed experimental parameters were chosen in Table 1.
where V is the velocity, m/s; π ≈ 3.14; n is the stirring speed in the stirring system, r/s; r is the radius of stirring paddles, m; n = 80 r/min, r = 5 cm.
Data analyses
The algal cell concentration was enumerated in a counting chamber (Minato TATAI) under an optical microscope (Olympus CX31) after being fixed by Lugol solution. The images of the aquatic particles and algae were recorded using the microscope (Olympus CX31) and image acquisition software (Algacount-Mx, Shineso). Eq. (2) was used to calculate the flocculation efficiency.
where C0 (cells/mL) is the initial algal cell density of the experimental group, C1 (cells/mL) is the algal cell density of the experimental group after sedimentation; C0′ (cells/mL) is the initial algal cell density of the control group, C1′ (cells/mL) is the algal cell density of the control group after sedimentation.
TN and dissolved TN (filtered by 0.45 μm membrane) in the suspension of aquatic particles were measured with the alkaline potassium persulfate digestion–UV spectrophotometric method. TP and dissolved TP were measured using the ammonium molybdate method (SEPA 2012). The result is shown in Table 2. Zeta potentials of the aquatic particles and algae were measured by Zetasizer Nano (Zetasizer 2000; Malvern). Size distributions of them were measured by Laser Particle Size Analyzers (Mastersizer 2000). Extracellular polysaccharides (EPSs) of the algae were stained with Alcian Blue according to Crayton (1982). To be specific, two drops of the cell suspension were mixed with five drops of stain (0.1% w/v Alcian Blue 8GX in 0.5 N acetic acid, pH 2.5) in a small test tube, mixed and allowed to stand at the room temperature for 5 min. Samples were observed and photographed using a microscope under bright field illumination at a magnification of 200 × . The concentrations of EPS were determined by the phenol-H2SO4 method (Dubois et al., 1956; Yang et al., 2008).
TN, total nitrogen; TP, total phosphorus; DTN, dissolved total nitrogen; DTP, dissolved total phosphorus.
Results
Characteristics of aquatic particles and algae
Figure 1 shows the microscopic morphologies of aquatic particles and algal cells. M. aeruginosa grew in single cells (size 4–6 μm, with round or oval shape) or small colonies (size 10–80 μm). Nitzschia was wedge shaped with gradually narrowed ends. The single cell of Nitzschia (15 μm in length and 4.5 μm in width) was bigger than M. aeruginosa. The volume and mass of Nitzschia were thus larger than M. aeruginosa. As indicated in Fig. 2, EPS concentrations of M. aeruginosa and Nitzschia cell wall were 3.64 × 10−2 mg/mg and 2.86 × 10−2 mg/mg, respectively.

Microscopic morphologies of particles and algal cells.
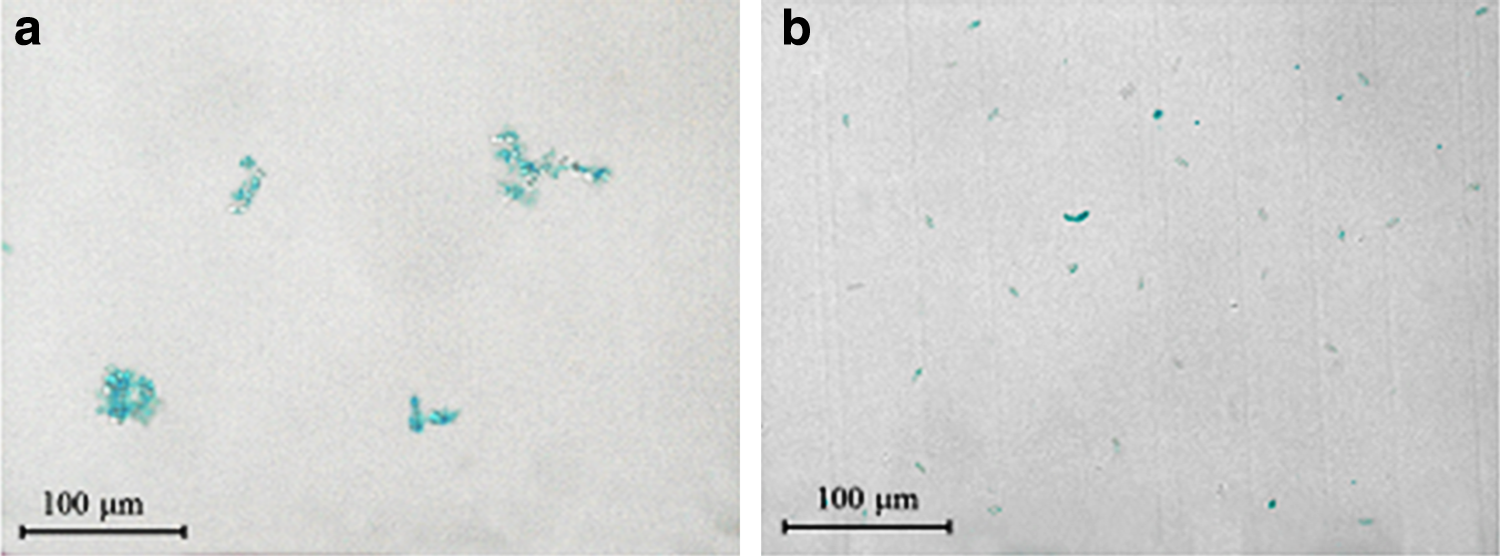
Micrographs of Microcystis aeruginosa and Nitzschia stained with Alcian Blue
Table 3 presented the size distribution of aquatic particles. The median diameters (D50) of 60, 100, 200, 230, and 300 mesh were 22.92, 20.59, 14.32, 12.47, and 10.72, respectively. As indicated in Table 3 and Fig. 3, the volume percentage of 1–10 μm particles in the sieved particles gradually increased with the reduction of D50, which implied that the corresponding viscous component in the sieved particles gradually increased. Specifically, the volume percentage of particles with the diameter of 1–5 μm was 12.58% when D50 was 22.92 μm (60 mesh) and it increased to 19.42% when D50 reduced to 10.72 μm (300 mesh).
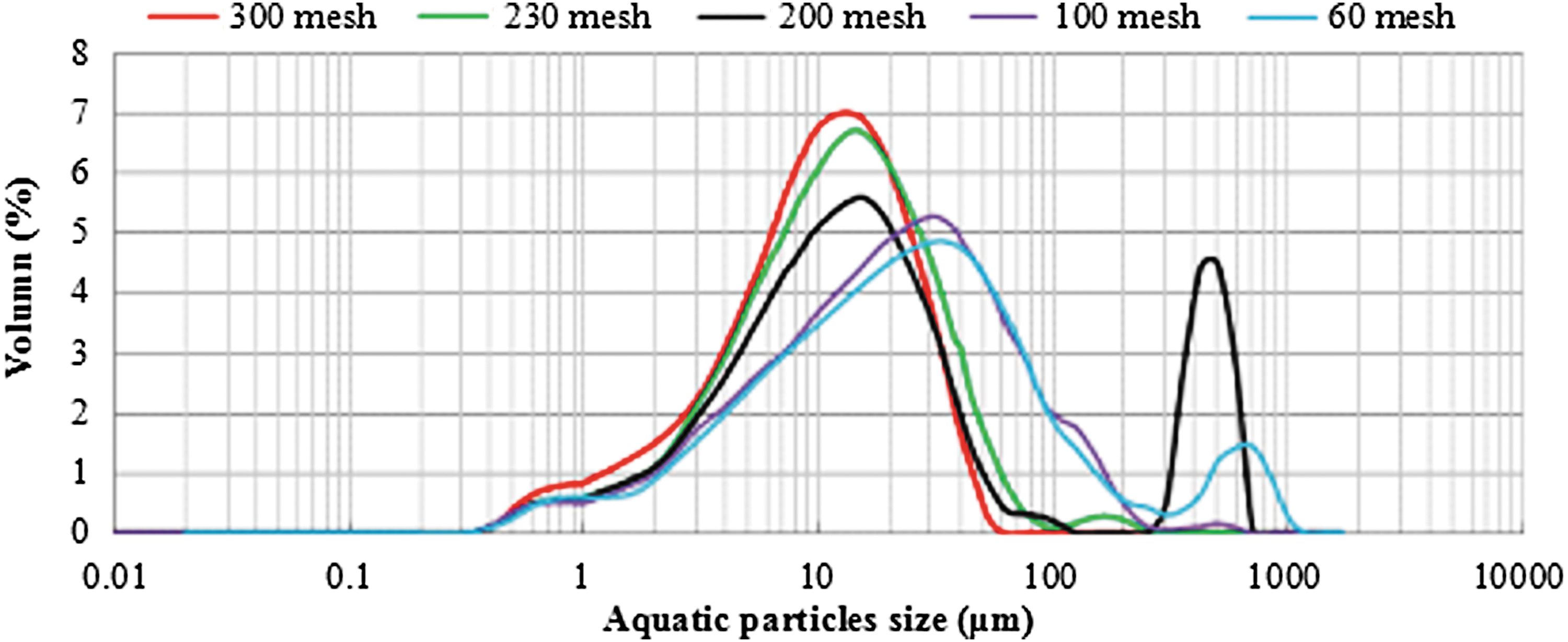
Size distributions of aquatic particles.
Effect of particle size on flocculation efficiency
Figure 4 presented the flocculation efficiency of algae with different particle sizes. As the particle size increased from 10.72 μm to 22.92 μm, the flocculation efficiencies of M. aeruginosa and Nitzschia decreased accordingly. The corresponding variations were 30.1% to 11.4% for the former and 17.0% to 5.1% for the latter. These results indicated that the effect of aquatic particle size on the flocculation of M. aeruginosa was greater than Nitzschia.

Flocculation efficiency of algae and zeta potential of particles under different particle sizes. Error bars in
Figure 4b showed the zeta potential of the particles of different sizes. The surface charge of the aquatic particles decreased as their size decreased. In natural lakes, aquatic particles and algae were negatively charged. The DLVO theory explains the aggregation of aqueous dispersions quantitatively and describes the force between charged surfaces interacting through a liquid medium. When the flocculation occurred between aquatic particles and algae cells, the electrostatic repulsion between them should increase with a decrease of the particle size according to DLVO theory. DLVO theory suggested that the interparticle forces (van der Waals and electrostatic repulsion forces) play key roles in flocculation, along with the hydrodynamics and other conditions of the suspensions (the size, shape, density, surface charge etc.). The speculation is different from the result represented in Fig. 4a. It seems that the electrostatic neutralization didn't dominate particles–algae flocculation process.
Effect of aquatic particle concentration on flocculation efficiency
Flocculation efficiencies of the algae under different particle concentrations are presented in Fig. 5a. The flocculation efficiency of M. aeruginosa and Nitzschia sharply increased from 19.3% to 43.7% and 2.95% to 29.0%, respectively, when the concentration of aquatic particles increased from 0.02 g/L to 1.28 g/L. The effect of particle concentration on the flocculating efficiency of both M. aeruginosa and Nitzschia was significant. However, as the aquatic particle concentration increased from 0.02 g/L to 1.28 g/L, the surface charge of particles varied insignificantly between −18.9 mV and −19.6 mV (Fig. 5b).
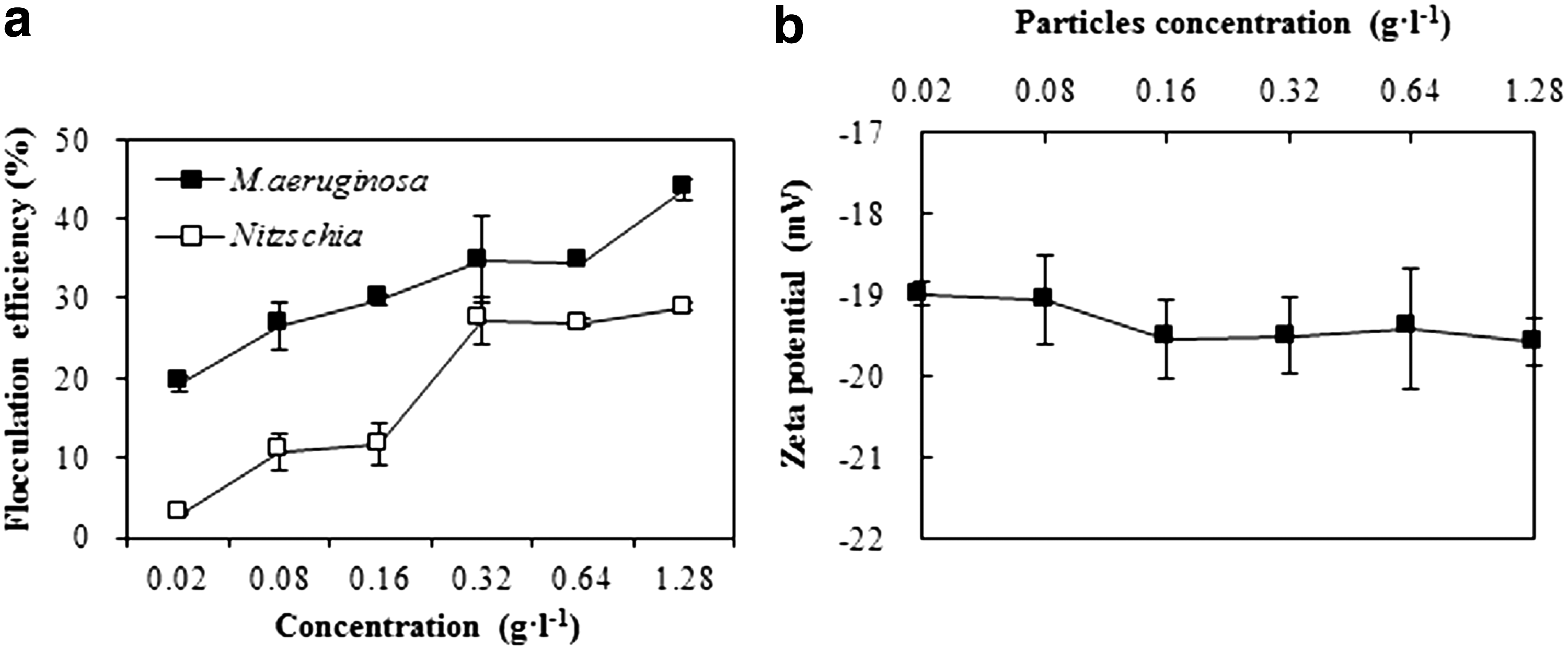
Flocculation efficiency of algae and zeta potential of particles under different particle concentrations. Error bars in
Effect of initial algal cell density on flocculation efficiency
Flocculation efficiency of the algae and zeta potential of M. aeruginosa and Nitzschia under different algal cell densities is presented in Fig. 6a and b. The flocculation efficiency of M. aeruginosa and Nitzschia increased with the increasing cell density until it reached a plateau when the concentration of aquatic particles was the same. When the initial cell density was 1.8 × 108 cells/mL, the flocculation efficiency of M. aeruginosa and Nitzschia reached the maximum. At this cell density, the increase of the flocculation efficiency of M. aeruginosa was two to three times higher compared with Nitzschia, indicating that the algal cell density displayed a more significant effect on the flocculation of M. aeruginosa than Nitzschia. The surface charges of M. aeruginosa and Nitzschia cells decreased with the increase of the initial cell density until 1.0 × 106 cells/mL. The decreasing trend stabilized or reversed at higher cell densities.

Flocculation efficiency of algae and zeta potential of M. aeruginosa and Nitzschia under different algal cell densities. Error bars in
Effect of organic matter concentrations on flocculation efficiency
As shown in Fig. 7a, the flocculation efficiency of M. aeruginosa increased from 17.6% to 23.4% when the humic acid concentration rose from 0 mg/L to 8 mg/L, with a slight reduction at 2 mg/L. With a further increase of the humic acid concentration, the flocculation efficiency did not change. Similar to M. aeruginosa, the flocculation efficiency of Nitzschia decreased slightly from 11.8% to 10.7% when 2 mg/L humic acid was added in the medium. With a further increase of the humic acid concentration, the flocculation efficiency of Nitzschia increased to 16.3% at 4 mg/L and decreased to 8.3% at 32 mg/L. For the two kinds of algae, the increase of the humic acid concentration did not change their surface charges significantly (Fig. 7b). A measure of 2 mg/L humic acid increased the surface charges of the two kinds of algae slightly. The surface charge of aquatic particles decreased largely when the humic acid concentration increased from 0 mg/L to 4 mg/L.
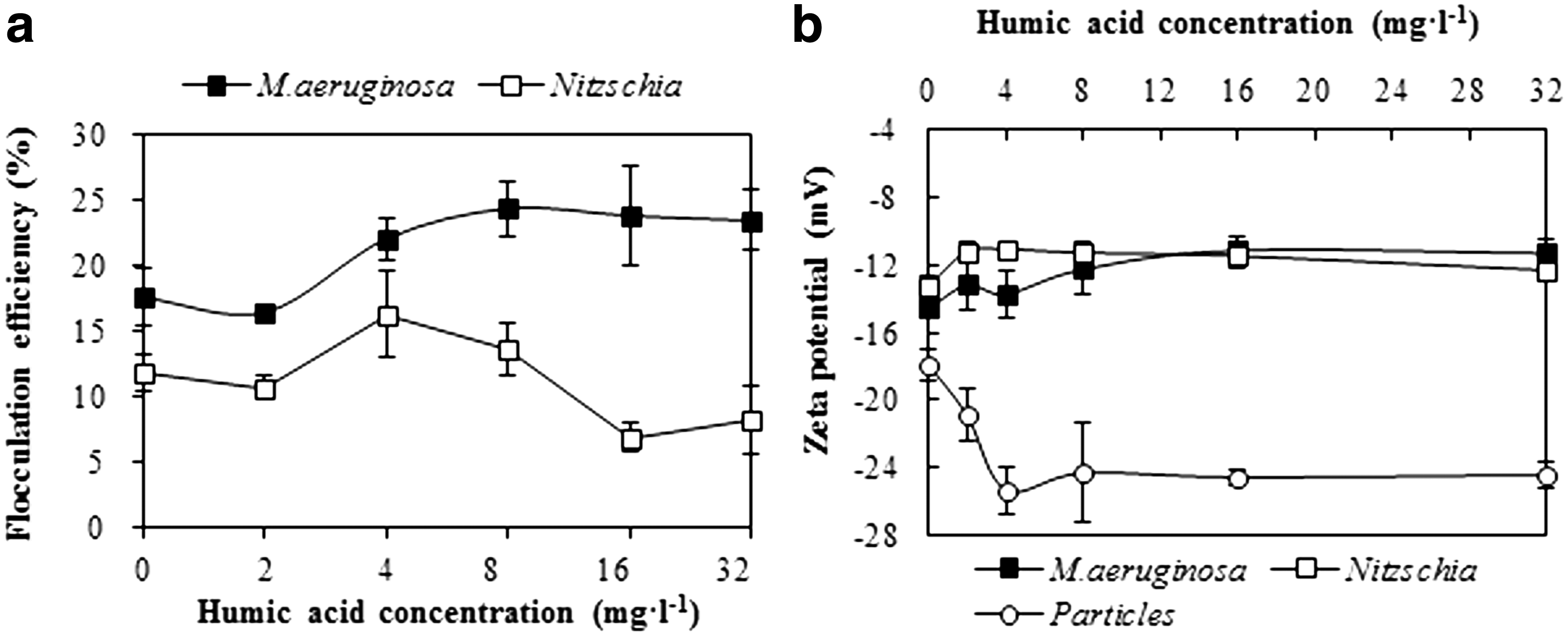
Flocculation efficiency of algae and zeta potential of algae and particles under different humic acid concentrations. Error bars in
Discussion
Results of Figs. 4 to 6 suggested that the electrostatic repulsion effect of the surface charge was not the key factors affecting the flocculation between aquatic particles and algal cells. With the decrease of aquatic particle sizes, the flocculation efficiency of M. aeruginosa increased, and the efficiency of M. aeruginosa was much higher than Nitzschia. The flocculation efficiency of M. aeruginosa was always higher compared with Nitzschia. One of the reasons may be that the size of M. aeruginosa was smaller than Nitzschia. Li (2005) showed that the smaller the algal cells were, the larger the specific surface area was, and the stronger the flocculation efficiency was. According to the shape of the algae, we know that M. aeruginosa cells with their round shape have bigger specific surface area than Nitzschia, giving rise to the higher flocculation efficiency of M. aeruginosa than Nitzschia. It has been proposed that the highest collision efficiency occurred when the size of clay particles was similar to the size of algal cells (Han and Kim, 2001). According to Table 3 and Fig. 3, the proportion of particles with the size of 1–5 μm increased when the meshes that particles were sieved increased from 60 mesh to 300 mesh. Moreover, the particles in the size range of 1–5 μm were the viscous component (Walling and Moorehead, 1989). In our study, the size of single M. aeruginosa cells (5 μm) was smaller than Nitzschia (4.5 μm in width, 15 μm in length) and was closer to the size of the adhesive component of aquatic particles (<5 μm), resulting in the higher flocculation efficiency of M. aeruginosa than Nitzschia.
Algal cells can secrete EPS for sand fixation and good adhesion. M. aeruginosa synthesized and secreted much more EPS than Nitzschia. Thus, M. aeruginosa cells had higher viscosity, which improved the flocculation efficiency of M. aeruginosa. In addition, motility was an important factor affecting flocculation process of particles on algae cells. Algae with fast moving speed could escape from the flocs preferentially and resume their growth (Pan et al., 2011). The self-sedimentation movement of Nitzschia cells was more significant because of its larger density and volume, which weakened the flocculation efficiency by aquatic particles. Self-sedimentation rate of diatoms was much higher than cyanobacteria both in light or darkness. The rate of diatoms and cyanobacteria was 0.240 m/day and 0.020 m/day that experienced 12 h of darkness, respectively, and was 0.288 m/day and 0.030 m/day that experienced 12 h of light, respectively (Wang et al., 2015).
Flocculation mainly involves two discrete steps as follows: transport and attachment. The transport step leads to the collision of two particles, which is caused by the random Brownian motion of the particles and/or the difference in the settling velocities of the individual particles. Attachment of the colliding particles is induced by van der Waals force and electrostatic attractions, which are largely dependent on the nature of the particle surfaces (Gerde et al., 2014). When the concentration of the aquatic particles or the algal cell density increased, the distance between single aquatic particles and algal cells shortened. The interaction force between them increased (Tuna et al., 2003), and the collision caused by the Brownian motion became more frequent, leading to the enhanced flocculation effect of the particles on the algal cells. Moreover, the increase in the particle–particle interaction decreased the sedimentation velocity of the particles (Kourki and Famili, 2012), which may increase the effective collision probability of aquatic particles and algae (Gerde et al., 2014). Besides, the increase of the particles concentration and the density of algae cells in unit volume increased the number of small size particles and the content of EPS on algae surface. This strengthened the effective contact between aquatic particles and algae cells and enhanced the viscous fixation (Wyatt et al., 2012), promoting the flocculation effect.
Organic matter concentration decreased the flocculation efficiency of M. aeruginosa and Nitzschia with the humic acid concentration increased from 0 mg/L to 2 mg/L. The flocculation efficiency of M. aeruginosa and Nitzschia was, respectively, promoted by 2–8 mg/L and 2–4 mg/L humic acid, but the adverse effect was observed for Nitzschia when the concentration exceeded 4 mg/L. Humic acid is acidic and negatively charged due to the presence of oxygen-containing functional groups, and it will produce the electrostatic repulsion contacting with the aquatic particles which are also negatively charged (Pallier et al., 2010). Electrostatic repulsion and van der Waals force between aquatic particles and algal cells would be changed as a result of the adsorption of humic acid on aquatic particles and algal cells (Gu et al., 1994; Avena et al., 1999). According to the formula of electrostatic repulsion (Park et al., 2001; Shang et al., 2015), the concentration of 0–2 mg/L humic acid enhanced the electrostatic repulsion of aquatic particles and algae which was bad for the flocculation. However, the decrease of flocculation efficiency of M. aeruginosa and Nitzschia with the humic acid concentration increasing from 0 mg/L to 2 mg/L was in the range of experimental errors. Therefore, it was difficult to determine the accurate effect of 2 mg/L humic acid. When the concentration of humic acid exceeds 4 mg/L, zeta potential of algae and aquatic particles changed insignificantly. However, the flocculation efficiency of M. aeruginosa increased, and the flocculation efficiency of Nitzschia decreased. Because M. aeruginosa surface secretes the viscous EPS, it may be covered by much more humic acid than Nitzschia. This increased the volume of M. aeruginosa cells, van der Waals, and the zeta potential of M. aeruginosa cell surface, promoting the flocculation sedimentation of particles on M. aeruginosa. When the humic acid concentration was greater than 4 mg/L, zeta potential of Nitzschia showed little changes. It may be that the surface of Nitzschia cells was totally covered with humic acid, and there will be excess humic acid with negative charge in the system, reducing the flocculation settlement effect.
Lake Poyang is a big Yangtze-connected lake. The suspended solid concentration in Lake Poyang (mean value 84.5 mg/L) was much higher than other lakes in the subtropical area such as Lake Taihu (35.8 mg/L in Taihu Meiliang Bay) and Lake Chaohu (20–50 mg/L) (Chen et al., 2003; Jin et al., 2010). In April, the five tributaries of Lake Erhai enter the flood season, carrying a large number of aquatic particles into the lake. During July to September, the aquatic particles are deposited in the lake (Ma et al., 2003). After the operation of Three Gorges Dam, the velocity in Lake Poyang increased due to the increase of the discharge volume, resulting in an increase of the aquatic particle concentration in the lake. The result that the increase of the flocculation efficiency of M. aeruginosa due to the increase of the aquatic particle concentration was much more outstanding than Nitzschia favored the flocculation of cyanobacteria and thus not beneficial for cyanobacteria to gain the predominance. The humic acid also contributed to the preferential flocculation of aquatic particles on cyanobacteria and helped to reduce the risk of cyanobacteria blooms in Lake Poyang. The higher the organic matter content, the greater the flocculation sedimentation efficiency of cyanobacteria. Low concentration of organic matter (0–4 mg/L) facilitated the flocculation sedimentation efficiency of particles on diatoms and inhibited the flocculation efficiency of diatoms when the concentration was more than 4 mg/L. In conclusion, aquatic particles in natural lakes such as Lake Poyang had a significant effect on the evolution of cyanobacteria and diatoms. Aquatic particles decreased the dominance of cyanobacteria and increased the dominance of diatoms, subsequently reducing the cyanobacteria bloom risk of Lake Poyang.
Conclusions
In natural water bodies, the flocculation of algae by aquatic particles was highly dependent on the characteristics of aquatic particles and algae. The flocculating efficiency of the algae increased with the decrease of aquatic particle size (D50 decreased from 22.92 μm to 10.72 μm), the increase of aquatic particle concentration (from 0.02 g/L to 1.28 g/L), and the increase of initial algal cell densities (from 4.5 × 105 to 1.2 × 108 cells/mL). The increase of organic matter concentration (humic acid increased from 2 mg/L to 8 mg/L) also had a positive effect on the flocculation of the cyanobacteria. The flocculation efficiency of aquatic particles on cyanobacteria was higher than that on diatoms, which was not conducive to cyanobacteria blooms in Lake Poyang. In conclusion, algae size and the EPS contents on algae cell surface, algal cell density, the concentration of particles, and organic matter were the main factors affecting the flocculation efficiency of aquatic particles on algae in natural water bodies like Lake Poyang.
Footnotes
Acknowledgments
This work is jointly supported by the National Basic Research Program of China (973 Program) (No. 2012CB417004) and the National Natural Science Foundation of China (No. 51078341, No. 50938007-01).
Author Disclosure Statement
No competing financial interests exist.