
Abstract
Abstract
Azo dyes represent the most important class of textile dyes and pose a serious environmental issue when released in wastewater. White rot fungi decolorize azo dyes efficiently; however, some difficulties arise when they are cultured as suspended pellets. This study analyzed decolorization performance of an azo dye by Trametes versicolor immobilized on polyurethane foam in a sequencing batch reactor (SBR). The SBR was operated for 27 days with nine cycles of 73.25 h each and dye decolorization of ∼96% ± 1.39%, irrespective of initial dye concentration and the number of cycles. In the SBR, free biomass barely accounted for 5% of total biomass, which did not significantly (p > 0.05) contribute to the decolorization process or biomass conglomerate formation. Along the SBR culture, laccase, manganese, and lignin peroxidase activities were produced with a sustained increase, related to the decolorization process, reaching maximal activities (15.9, 40.4, and 14.4 U/L, respectively) at the end of the culture. Changes observed in the absorption spectrum showed that dye decolorization was related to a change in dye chemical structure, attributed to the action of enzymatic activity. The kinetic study of decolorization and enzymatic activity, by means of a first-order model, suggested that decolorization rates could decrease at higher dye concentrations as well as by increasing the cycle's number in the SBR. On the other hand, higher dye concentration could produce an increase in the total enzyme production rate, probably as a physiological response of the immobilized fungus.
Introduction
E
Textile effluents are mainly treated by physicochemical methods, which are often expensive, showing low efficiency and generally not able to eliminate the dyes completely; with these techniques, accumulation of concentrated sludge can create a disposal problem. A detailed review of physicochemical and biological methods for decolorization and degradation of dyes has been published elsewhere (Singh and Arora, 2011). Among biological processes for degradation of dyes, those catalyzed with fungi have been investigated extensively because most basidiomycetes are tolerant to high concentrations of pollutants, eco-friendly, and cost-effective (Ellouze and Sayadi, 2016).
Research has focused on white-rot fungi (WRF) as a possible alternative to the present treatment technologies. WRF have a unique capacity to degrade recalcitrant organics and dyes mainly by the action of nonspecific extracellular enzymes such as laccase (Lcc), lignin peroxidase (LiP), and manganese peroxidase (MnP). Despite the large amount of information available on the use of WRF to decolorize wastewater containing different types of dyes (Si et al., 2013; Chenaux et al., 2014; Imran et al., 2015), its implementation as a technology remains a challenge. The application of WRF in large-scale processes has been limited by the lack of bioreactor systems that can operate for prolonged periods with sustained secretion of enzymes, together with controlled growth of fungi (Hossain et al., 2015).
The sequencing batch reactor (SBR) has been applied in textile wastewater treatment, especially for the removal of azo dyes because they allow the fungal activity to be maintained for a long period and may achieve better results in dye decolorization when compared with batch cultivation (Borchert and Libra, 2001; Sanghi et al., 2006, 2011). However, with suspended cell culture, excessive growth of mycelia gives rise to practical and technical difficulties (Sathian et al., 2014).
Immobilized fungal bioreactors have been found to exhibit good biological activities and abilities during long operational periods. Currently, immobilization of fungal cells resulted in high cell densities and extracellular enzyme production without the problems typically associated with dispersed pellets. In addition, this culture mode can enhance fungal mechanical strength and facilitate the reuse of biomass (Rodríguez-Couto, 2009), favoring the SBR operation. However, there is no information concerning the use of immobilized Trametes versicolor in SBRs for dye removal.
The objective of this work was to analyze the decolorization performance of an azo dye by T. versicolor immobilized on polyurethane foam (PUF) in an SBR. The kinetics of decolorization and production of ligninolytic enzyme activities were also studied.
Materials and Methods
Microorganism
White-rot fungus, T. versicolor CDBB-H1051, was grown and maintained on malt extract agar plates at 4°C with periodic transfer. Plugs of agar (diameter 1 cm) were taken from the periphery of the culture to serve as inoculum sources.
Dye
Vinyl sulfone azo dye RB5 (C26 H21 N5 O19 S6 Na4) was purchased from Química Mexibras, (Mexico City, Mexico). The main commercial applications are in dyed cotton, wool, nylon, silk, hair, and wood (Hadibarata et al., 2013).
Immobilization of fungi on PUF
T. versicolor immobilization on PUF cubes was done in Sabouraud dextrose broth using the technique described by Martínez-Sanchez et al. (2018). The dry weight of the fungal biomass immobilized into PUF cubes was determined by drying and then weighing the PUF cubes before and after fungal growth.
Medium
All decolorization assays were performed in mineral medium (Radha, 2005), containing per liter: glucose, 2 g; NH4Cl, 0.05 g; KH2PO4, 2 g; MgSO4, 0.5 g; CaCl2, 0.1 g; and 10 mL of trace element. The trace element solution contained (g/L) MnSO4·H2O, 0.5; NaCl, 1; FeSO4·7H2O, 0.1; CoCl2·6H2O, 0.185; ZnSO4·7H2O, 0.11; Na2MO4·2H2O, 0.011; and H3BO3, 0.011. In all assays, pH was adjusted to 5. All reagents were purchased from JT Baker® Mexico.
Sequencing batch reactor
Decolorization assays of RB5 were carried out in a glass vessel of 1-L capacity. The reactor, with a length of 0.19 m and width of 0.11 m, was operated with a maximum working volume of 750 mL. A tube was inserted into the reactor top to ensure filling and discharge of the medium using peristaltic pumps. The reactor was stirred with a Teflon-coated magnetic stirring bar. Air was supplied aseptically to the bioreactor, during the decolonization step, by using a sterilized cotton fiber and an air stone diffuser connected to a room-aquarium aeration pump. At the beginning, the SBR was inoculated with the fungal-colonized PUF equivalent to 0.27 g of immobilized T. versicolor per liter, whose weight was determined previously by means of dry cell mass.
The SBR was operated during a period of 648 h with cycles of 73.25 h on average. For each cycle, the reactor was filled, during the first 0.5 h, with sterilized culture medium containing RB5 and then inoculated with the fungal-colonized PUF. The reaction step, 72 h on average, consisted of an aerated operation in which fungi grew until achieving 96 ± 1.39% decolorization of RB5. Afterward, aeration was shut down and settling of free cells was carried out for 0.5 h. Finally, 0.35 L of medium was discharged from the reactor within 0.25 h to start a new cycle. The reactor was filled with sterilized culture medium containing 2 g/L glucose, 0.05 g/L NH4Cl, and the corresponding RB5 dye concentration (100, 150, or 200 ppm), three cycles were carried out for each dye concentration. Samples were taken at the beginning and along the cycles to measure dye decolorization, residual glucose, and ligninolytic enzyme activities.
Kinetic study
A kinetic study was done for each operation cycle to analyze (1) the decolorization rate of RB5 and (2) production of summative enzymatic activity of Lcc, MnP, and LiP. These enzymes can act simultaneously or synergistically; for this reason, we evaluated the kinetics for enzyme production considering the summative activity of the three enzymes.
(1) A modified first-order model (Sathian et al., 2014) can describe the RB5 decolorization rate
where A corresponds to the RB5 absorbance at 595 nm, A0 is absorbance at the beginning of every cycle, A∞ is absorbance at the end of the cycle, t is the cycle time (h), and kd is the pseudo-first-order decolorization rate constant (d−1). By integrating, we get
RB5 decolorization rate during the SBR operation decreased gradually and became nearly constant after a certain period of operation, so A∞ was considered as the minimal final RB5 absorbance obtained in the three assays from each dye concentration.
(2) Furthermore, we propose a first-order kinetic expression to describe the production of total enzyme activity:
In Equation (3), E represents the sum of enzymatic activities (Lcc, MnP, and LiP) at t and kE is the first-order total enzyme production rate constant (d−1). It is possible to obtain kE by integrating the above equation
where Eo was the amount of enzymatic activity at the beginning of every cycle.
Analytical methods
Decolorization studies and glucose consumption
The samples were centrifuged at 4,000 rpm for 5 min. The supernatants were collected for further analysis (RB5 decolorization, residual glucose concentration, and enzyme activities). RB5 decolorization was measured spectrophotometrically at 597 nm with a Genesys UV-VIS spectrophotometer; changes in spectral absorption were monitored in the liquid medium by scanning the spectrum between 200 and 700 nm. Dye decolorization was expressed as a percentage of the removed dye, comparing the concentration at any time with that corresponding to the start of each cycle. The residual glucose concentration was quantified by the dinitrosalicylic acid method (Blum et al., 1960).
Enzyme assays
Lcc activity was evaluated spectrophotometrically using 10 mM guaiacol (2-metoxyphenol) in 100 mM acetate buffer (pH 5) and 0.05 mL of culture-centrifuged supernatant. Oxidation of guaiacol at 30°C ± 3°C was monitored through the increase in absorbance at 470 nm (
Toxicity assays
Assays were performed using a modified method with serial dilutions in yeast mannitol broth (YMB) of the reference strains, Escherichia coli and Pseudomonas aeruginosa (Enayatzamir et al., 2011). RB5 stock solution and samples obtained from SBR decolorization assays were centrifuged (15 min at 4,000 rpm and room temperature) and then successively filtered through fast filter paper. Test tubes containing YMB and RB5 samples, in a final volume of 3 mL, were inoculated with 0.5 mL of bacterial culture and incubated (35°C ± 3°C, 130 rpm). Serial dilutions were prepared with 0.9% NaCl solution after 24 h, 0.1 mL of each solution was spread onto yeast mannitol agar plates, and viable colonies were counted after 24 h. The antimicrobial activities of RB5 samples were estimated after comparison with the bacterial growth obtained in the same conditions using 1 g/L glucose in test tubes.
Results and Discussion
RB5 Decolorization in the SBR
T. versicolor has been recognized for its high capacity of decolorizing various dyes commonly used in textile industries (Yang et al., 2017); however, to the best of our knowledge, there are no reports in which this fungus has been used as immobilized cells in an SBR. With this method of culturing, T. versicolor was capable of decolorizing RB5 for several SBR cycles over an extended period of 27 days. All cycles started with a blue color, whose intensity depended on the RB5 initial concentration; the color at the end of the cycles changed from a pink tone (first cycle) to a colorless medium at the end of the ninth cycle. In all cycles, fast decolorization occurred in the first 24 h and, subsequently, RB5 concentration changed slowly. Thus, an average decolorization of 96% ± 1.39% was obtained at 72 h in each cycle, regardless of the initial dye concentration (Fig. 1).

RB5 decolorization in the SBR at average cycles of 72 h, after which a percentage of the volume was removed and fresh medium was added. Vertical dotted lines on the graph indicate the RB5 concentration used in the cycles; for each concentration, three different trials were carried out. RB5, Reactive Black 5; SBR, sequencing batch reactor.
These results were better than those obtained in the reactor operated in batch mode by Fernández et al. (2009), who reported 83% RB5 decolorization in 24 h by T. versicolor immobilized on Luffa sponge with 300 ppm RB5. Enayatzamir et al. (2009) found decolorization of 74%, 13%, 43%, and 52% in 24 h during four successive batches with 60 ppm RB5 using cultures of Trametes pubescens immobilized on stainless steel sponges. Even the decolorization, reported by our research group (Martínez-Sanchez et al., 2018), using T. versicolor immobilized in PUF was improved by 9%. Likewise, the results were better than those reported for the SBR; Sanghi et al. (2011) showed that free cells of Coriolus versicolor decolorized sulfonic azo dyes from 70% to 90%. Sathian et al. (2014) found maximum decolorization of 71.3% in about 3 days with the combination of mycelial pellets of Pleurotus floridanus, Ganoderma lucidum, and T. pubescens.
Despite the importance of relating the decolorization percentage with biomass growth to estimate decolorization efficiency, only a few studies addressed the effect of the inoculum size of free cells on dye decolorization by fungal strains (Vaithanomsat et al., 2010; Khan and Fulekar, 2017). Due to the difficulty in immobilized biomass quantification, there are no reports about immobilized biomass growth during dye decolorization. Thus, in this work, it was only possible to estimate the decolorization efficiency by comparing decolorization based on the initial biomass. In this regard, the results obtained in this work were better than those obtained by Martínez-Sanchez et al. (2018) who used 0.33 g/L as the inoculum, which confirms the greater efficiency of RB5 decolorization in the SBR.
Analysis of UV-Vis spectrum
Modification in absorption spectra of the decolorized medium is shown in Fig. 2. Only spectra of the third cycle of each RB5 concentration are presented because they represent global changes that occurred when increasing the initial dye concentration. It is important to note that RB5 absorbance, measured at the beginning of each cycle, is lower than the planned concentration (around 15.79% ± 4.38%) because the dye was decolorized upon contact with the residual medium and the immobilized microorganism contained in the reactor during the filling step.
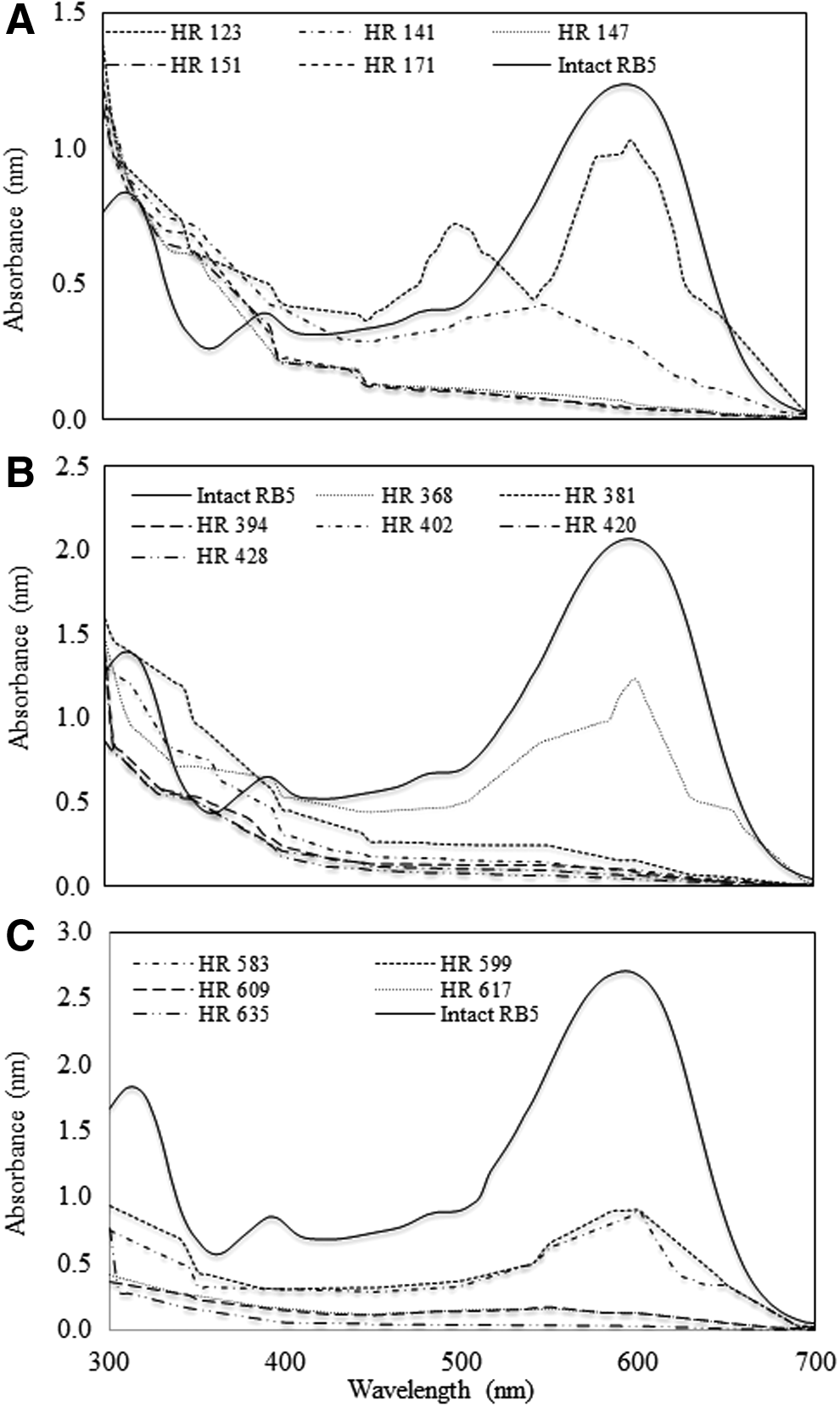
Absorption spectrum of RB5 decolorization in the SBR by Trametes versicolor at the third cycle of
With 100 ppm RB5 (Fig. 2A), from 123 h of culture, the intensity of absorbance decreased in the range from 400 to 650 nm, specifically the decrease at 595 nm suggested azo bond degradation (Hadibarata et al., 2013), whereas in the UV region (300–400 nm), the increased absorbance indicated cleavage of aromatic rings (Borchert and Libra, 2001; Kumar et al., 2012) and formation of some by-products (Castillo-Carvajal et al., 2012). The same changes in the UV-Vis spectrum also occurred during the following operation cycles of the SBR, but in shorter times, with a trend showing lower absorbance in the range from 300 to 700 nm toward the end of every cycle.
Glucose consumption
Glucose degradation in the SBR is shown in Fig. 3. The highest glucose concentration measured at the end of the first cycle suggested that addition of RB5 altered the activity of T. versicolor; however, starting with the second cycle, the microorganism became acclimated and, consequently, the consumption of glucose was almost total in subsequent cycles. Furthermore, the continuous glucose consumption observed appears to be related to RB5 decolorization, similar to that reported by Borchert and Libra (2001) and in agreement with the 87% RB5 decolorization reported by Castillo-Carvajal et al. (2012) with the addition of 2 g/L of glucose.
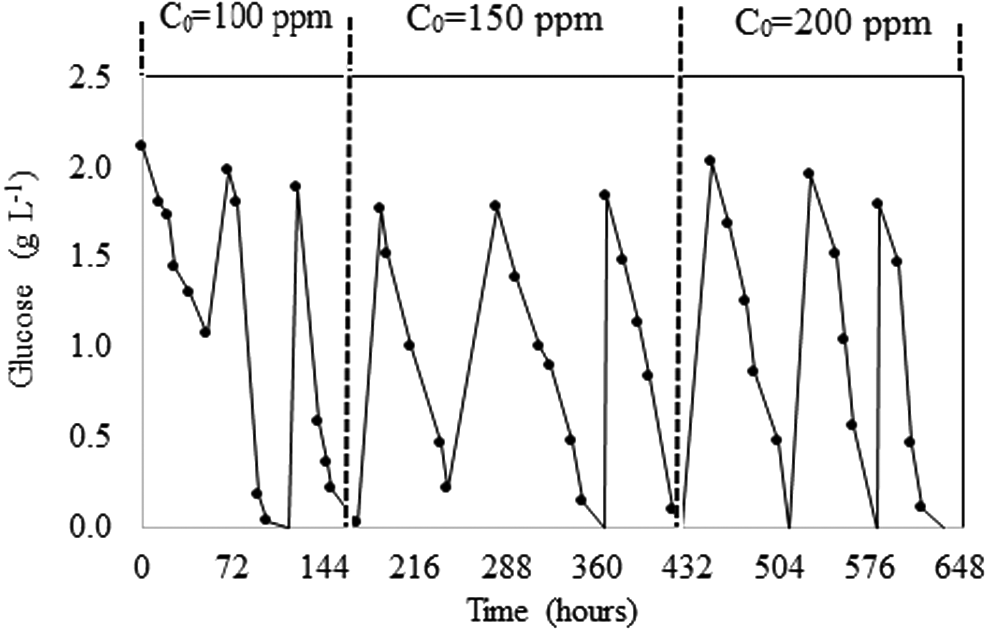
Residual glucose during RB5 decolorization in the SBR at average cycles of 72 h, after which a percentage of the volume was removed and fresh medium was added. Vertical dotted lines on the graph indicate the RB5 concentration used in the cycles; for each concentration, three different trials were carried out.
Enzymatic activity production on the SBR
Enzymatic activity produced during RB5 decolorization by T. versicolor (Fig. 4) showed a gradual increase as well as a change in the proportion in each cycle. At the end of the first cycle, the predominant activity was LiP, followed by Lcc and MnP, whereas in the ninth cycle, MnP was predominant, followed by Lcc and LiP. At the end of the last cycle, Lcc, MnP, and LiP activities increased 7.7, 8.6, and 2.3 times, respectively, over the activity detected at the end of the first cycle. This behavior is congruent with that reported by Enayatzamir et al. (2009) during RB5 decolorization, although the presence of the three enzymes differs from previous studies. Castillo-Carvajal et al. (2012) and Ottoni et al. (2013) reported the production of Lcc and LiP, and Lee et al. (2006) reported the production of Lcc and MnP by T. versicolor during RB5 decolorization. This discrepancy is explained considering that the relative contribution of each enzyme to dye decolorization might be different for each fungal strain (Vaithanomsat et al., 2010).
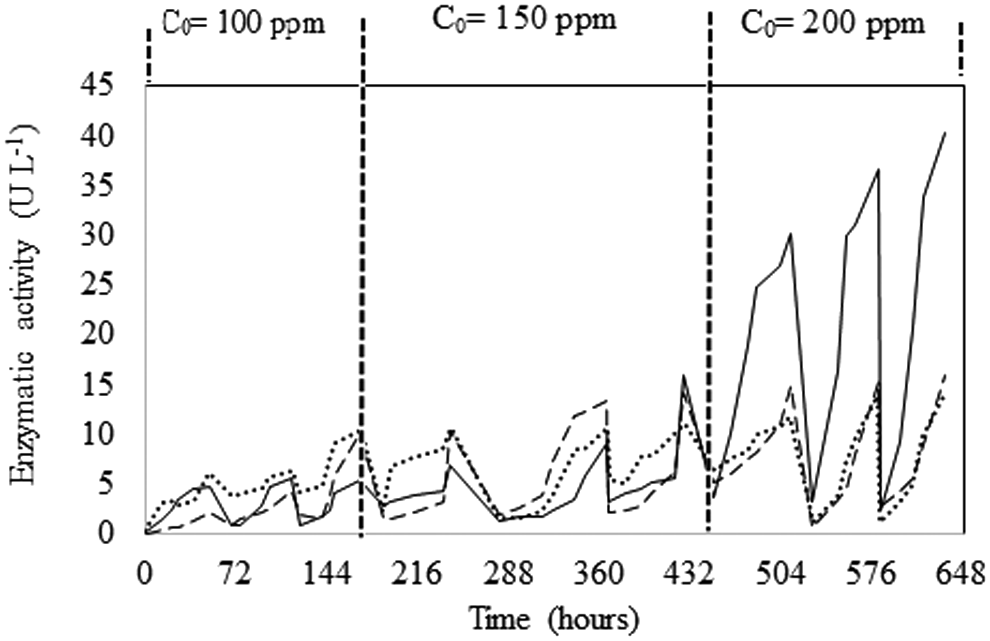
Time course of enzymatic activities Lcc (----), MnP (___), and LiP (•••) produced by T. versicolor in the SBR during RB decolorization. Vertical dotted lines on the graph indicate the RB5 concentration added in the cycles; for each concentration, three different trials were carried out. Lcc, laccase; LiP, lignin peroxidase; MnP, manganese peroxidase.
The enzymatic activities detected seemed to be related to RB5 decolorization, unlike that observed by Castillo-Carvajal et al. (2012), who found that the LiP activity was induced by Luffa cylindrica sponge used as an immobilization support. However, in the present work, PUF only acts as a physical matrix for hyphal growth, so LiP activity could be associated with RB5 decolorization. Therefore, production of the three oxidases can be dependent on conditions established during dye decolorization and could explain the high percentage of decolorization, regardless of RB5 concentration. It is known that decolorization occurs through fungal adsorption by mycelium, followed by degradation (Parshetti et al., 2007). Considering the results, it can be stated that the decolorization carried out by T. versicolor immobilized in the PUF could be related mainly to its enzymatic activity and in low proportion to the adsorptive capacity of the biomass and PUF. This was shown by a decrease in RB5 absorbance at the wavelength of maximum absorption, which occurred without changes in the profile absorption spectrum, obtained in the first decolorization assays in the SBR (data not shown).
Toxicity assays
Metabolites produced from any dye decolorization process vary in their toxicity according to the organism, the presence of different isoenzymes, enzymatic specificities, and chemical structure of the dye (Kandelbauer and Guebitz, 2005). Therefore, it was necessary to evaluate the toxicity of the obtained dye derivatives from the decolorization process to determine whether they were environmentally safe. In this study, products generated during RB5 decolorization, as well as the original dye, were not toxic to E. coli and P. aeruginosa. These results are congruent with the lack of phytotoxicity of biodegradation products from RB5 produced by Phanerochaete sordida (Permpornsakul et al., 2016) as well as the capacity of Phanerochaete chrysosporium to convert RB5 into a nontoxic metabolite to soil fungi (Enayatzamir et al., 2011). Therefore, neither the original dye structure nor the transformation products are toxic; however, it would be interesting to characterize the metabolites produced during RB5 decolorization by T. versicolor in the SBR.
Kinetic study
Experimental data of RB5 decolorization and summative enzymatic activity obtained during the SBR operation are shown in Fig. 5. Both profiles showed the same trend, regardless of the initial dye concentration. RB5 decolorization showed a first stage of fast decolorization rate and the subsequent second stage of a rather slow decolorization rate, which became nearly constant after 20 h of culture, while the total enzymatic activity was maintained with a constant increase during all assays.

Absorbance (—) for cycles 1 (◊), 2 (◯), and 3 (Δ) and summative enzymatic activity (___) for cycles 1 (●), 2 (■), and 3 (▲) during RB decolorization by T. versicolor in the SBR.
Application of a modified first-order model [Eq. (2)] and a first-order model [Eq. (4)] to the experimental data obtained in the three concentrations of RB5 analyzed indicated that both models have a relatively good fit (in most cycles, R2 > 0.90). Estimated constants for the models are shown in Table 1. At low concentrations of RB5 (100 and 150 ppm), kd constants had a similar behavior throughout the process: their value increased with the number of cycles and this increase was greater for the lower dye concentration. On the contrary, at higher levels of dye, the specific decolorization rate decreased with the number of cycles. Our results are consistent with Sanghi et al. (2011), who observed that an increase in the initial dye concentration tends to decrease the decolorization efficiency. Regarding kE, its value remains more or less constant along the process for each dye concentration, around 0.025 ± 0.003 and 0.023 ± 0.004 d−1 for 100 and 150 ppm, respectively; furthermore, its value was close to 0.021 d−1 in the first cycle of higher dye concentration. However, when the concentration of dye was high and the decolorization rate notably diminished, kE values increased; probably as a physiological response of the immobilized fungus.
Kinetic Constant Values for Reactive Black 5 Decolorization in Sequencing Batch Reactor During Three Cycles for Each Initial Dye Concentration
Summaries
The SBR operated with immobilized cells can solve problems associated with mycelial growth and, for this reason, RB5 decolorization was studied in the SBR using T. versicolor immobilized on PUF. In the SBR, no problems related to biomass conglomerates were produced because the free cell biomass was 5% of the total biomass produced. Dye decolorization was mainly attributed to enzymatic activity; accordingly, at the end of the culture, maximal activity was 15.9, 40.4, and 14.4 U/L for Lcc, MnP, and LiP, respectively. Changes observed in the UV-VIS spectrum suggested that the RB5 chemical structure was modified by enzymatic activity of T. versicolor, thus it was considered as the main mechanism of dye removal. Furthermore, changes in the spectra were consistent with color changes occurring in the reactor during the SBR operation; consequently, at the end of the last cycle, a colorless medium was obtained. It was also demonstrated that residual medium was not toxic to E. coli and P. aeruginosa. Kinetic studies showed that RB5 decolorization followed a first-order model and the efficiency of the SBR increases with culture time.
Finally, the use of T. versicolor to remove textile dyes from wastewater has shown to be a good alternative to traditional, wastewater treatment technologies. In this regard, the SBR increases the potential of T. versicolor because it was possible to maintain continuous ligninolytic enzyme production and, consequently, RB5 decolorization for a long time. Moreover, using T. versicolor as immobilized cells allows solving many of the practical and technical difficulties that are usually encountered with dispersed mycelium. However, some aspects require additional studies such as to determine changes that occur in the biomass during culture and to associate them with both decolorization and enzymatic activity. For scaling up the process, further study of the SBR behavior under nonsterile conditions is required, as well as knowledge about the performance of the fungus for longer operating times. This kinetic study provides a general idea of how the behavior of the culture would be at higher dye concentrations, for which it could be expected to achieve decolorization at lower rates, but with an increase in total production of enzymes involved.
Footnotes
Acknowledgments
This work was financially supported by The “Tecnológico Nacional de Mexico.” L.E.L.-G. thanks “Consejo Nacional de Ciencia y Tecnología” for the scholarship, No. 539526.
Author Disclosure Statement
No competing financial interests exist.