
Abstract
Abstract
Low-pressure membranes provide high-quality water through physical removal of bacteria, cysts, and particles. Bacteria can attach to the membrane surface and form biofilms that reduce energy efficiency and increase the frequency of membrane replacement. Modifying membranes with polydopamine, a chemical mimic of the adhesive proteins that mussels release, has the potential to delay biofilm formation. Dopamine polymerizes under mild conditions, forming a coating that is of interest in membrane applications due to its hydrophilic and reactive nature. Compared with hydrophobic base membranes, hydrophilic surfaces are resistant to bacterial attachment. Additionally, when polydopamine is exposed to silver nitrate, a reduction reaction takes place that embeds antimicrobial silver nanoparticles into the polydopamine layer. This study paired short-term deposition experiments with longer-term filtration experiments to determine the efficacy of this modification when challenged with Pseudomonas aeruginosa. Modification with polydopamine resulted in an average 48% reduction in the initial deposition rate of P. aeruginosa. However, hydrophilicity of the polydopamine layer did not have a significant impact on membrane performance over 3 days of bacterial filtration. Only the combination of both polydopamine and silver nanoparticles improved the rate of water production, with an average flux 1.7 times greater than that of the unmodified membrane. Confocal microscopy revealed heterogeneous bacterial growth on all membrane types with a lack of association between membrane performance and biofilm structure. Modifying membranes with polydopamine and silver nanoparticles has the potential to ameliorate the effects of biofouling, thereby increasing the efficiency of membrane water treatment.
Introduction
A
Membrane modification can delay biofouling by altering the interactions between the membrane surface and the bacteria in the feed solution (Kochkodan and Hilal, 2015). Dopamine is a chemical used for membrane modification that mimics the behavior of the adhesive proteins released by mussels, a bivalve mollusk. Mussels are known for their ability to stick to a wide range of materials due to the presence of catechol and amine groups in their adhesive proteins (Lee et al., 2007). Dopamine is the simplest compound containing these same functional groups (Chen et al., 2017) and polymerizes under mild conditions, requiring only the presence of oxygen and a pH slightly above neutral (Lee et al., 2007). When dopamine polymerizes, it forms a thin coating (tens of nm) on any exposed surface (Lee et al., 2007). Several theories have been developed regarding the reactions involved in the polymerization process (Lee et al., 2007; Dreyer et al., 2012; Hong et al., 2012; Liebscher et al., 2013; Chen et al., 2017). Once polymerized, this coating, like its biological inspiration, adheres to a wide array of materials, including metals, semiconductors, ceramics, and polymers (Lee et al., 2007). Dopamine deposition is uniform and can be regulated using dopamine concentration, deposition time, pH, and oxygen availability (Kasemset et al., 2013; Yang et al., 2015).
Polydopamine modification is of interest in membrane applications as base polymeric membranes are hydrophobic, whereas the thin polydopamine coating is hydrophilic (Arena et al., 2014; Tang et al., 2015; Huang et al., 2016; Liu and Hu, 2016; Yang et al., 2016a, 2016b; Zhang et al., 2016). Hydrophilic materials are less prone to organic and biological fouling as they form hydrogen bonds with water molecules, tightly binding a layer of water to the membrane surface and resulting in reduced foulant adsorption (Kim et al., 2003; Kochkodan and Hilal, 2015). Additionally, the polydopamine coating is reactive and can be used as a glue to attach materials to the membrane surface. Upon exposure to polydopamine, metallic salts, such as silver nitrate and copper (II) chloride, are reduced and their associated metallic nanoparticles are embedded in the polydopamine layer (Lee et al., 2007; Sureshkumar et al., 2011). The hydrophilic/antibacterial pairing of polydopamine and silver nanoparticles has been of specific interest due to the high antimicrobial activity of silver ions. A modified membrane that has a hydrophilic, antimicrobial nature has the potential to both delay initial deposition of bacteria and inactivate bacteria that deposit, thereby preventing biofilm development. Methods of evaluating the antiadhesive and antibacterial effects of these membranes vary widely across groups with most studies focusing on short-term impacts.
Although previous publications have addressed the effects of membrane modification with polydopamine and silver nanoparticles on bacterial attachment and survival upon exposure (Tang et al., 2015; Huang et al., 2016; Liu and Hu, 2016; Yang et al., 2016a, 2016b; Zhang et al., 2016), to our knowledge, studies that bridge the gap to applied use in a constant pressure system have not been conducted. The purpose of this article is to provide an understanding of how modifying low-pressure membranes with polydopamine and silver nanoparticles affect membrane performance. Through direct observation experiments, we quantified the initial rate of bacterial deposition onto the membrane surface. Through biofilm formation experiments, we simulated the conditions of a working treatment system and determined how modification affected flux decline in a constant pressure system. Finally, we used confocal microscopy to characterize biofilms formed in the system to assess how modification affected biofilm development. By pairing short and longer-term experimental methods, we elucidated the effects of modifying polysulfone ultrafiltration membranes with polydopamine and silver nanoparticles on biofouling.
Materials and Methods
Membrane fabrication and modification
Polysulfone membranes were made using the wet-phase inversion method (Strathmann and Kock, 1977). Polysulfone beads (15 wt %, Udel P3500; Solvay Advanced Polymers) were dissolved with LiCl in 1-methyl-2-pyrrolidone (NMP) at 55°C. For the biofilm formation experiments, the percentage of LiCl was increased (3.5 wt %) to increase porosity and permeate flux. This increase in initial flux was sought to better demonstrate the flux decline over time. Once the polysulfone and LiCl dissolved, the solution was allowed to degas for at least 72 h. The solution was then spread over a glass plate with a casting knife (Elcometer 3530/5 Adjustable Baker Film Applicator, Elcometer Limited) at a gate height of 50 μm. The glass plate was submerged in deionized (DI) water, resulting in the formation of a membrane sheet ca. 50 μm thick.
For polydopamine (PD) modification, the membrane was secured in a polycarbonate flow cell (90 × 38 × 2 mm), active-side facing solution. A solution of 1 g/L dopamine hydrochloride (Sigma-Aldrich) dissolved in Trizma hydrochloride (Sigma-Aldrich) buffer (pH 8.5) was pumped through the flow cell and recirculated for 6 h at a rate of 10 cm/s. Dopamine polymerized and formed a thin PD coating on the membrane surface. Following PD modification, the membrane was removed from the flow cell and rinsed with DI water. To incorporate silver nanoparticles into the PD coating (PD/Ag), the PD modified membrane was exposed to 50 mM AgNO3 for 2 h and then rinsed with DI water. Membranes prepared according to these methods have previously been characterized (Tang et al., 2015). The PD/Ag coating was verified through elemental analysis with X-ray photoelectron spectroscopy and energy-dispersive X-ray spectroscopy. Furthermore, the distribution of silver nanoparticles on the membrane surface was shown using scanning electron microscopy.
Preparation of bacterial solution
The bacterial species Pseudomonas aeruginosa PA01 was chosen for its ubiquity in the environment and its tendency to form biofilms. The species used in this study also has carbenicillin resistance, which was used to prevent the growth of other organisms. The P. aeruginosa is labeled with green fluorescent protein. Bacterial solutions were prepared by streaking P. aeruginosa onto an agar plate and leaving the plate overnight in an incubator at 30°C. A single colony was taken from the agar plate and transferred to LB Broth, Miller (Fisher Scientific) with 150 mg/L carbenicillin. The LB media was then left overnight in an incubator at 30°C. The next day, an aliquot of this overnight solution was pipetted into fresh LB media with 150 mg/L carbenicillin. When this bacterial solution reached the stationary growth phase (optical density = 0.8), it was removed from the incubator and washed (Tang et al., 2015). The bacterial solution was centrifuged at 4,300 rpm for 10 min followed by resuspension in 154 mM NaCl. The solution was centrifuged and resuspended twice more. Finally, the solution was resuspended in 10 mM NaCl and adjusted to an optical density of 1.0 (Benjamin and Lawler, 2013).
Direct observation system
The short-term system (Fig. 1) was used to characterize the attraction between bacteria in the feed solution and the membrane surface (Tang et al., 2015). A 10 mM NaCl solution adjusted to pH 7 using NaHCO3 was pumped from a pressurized feed tank through a flow cell holding the membrane. The system recirculates both the crossflow and permeates streams back to the feed tank. The flow cell was secured on the stage of an epifluorescence microscope (Nikon Eclipse E600W). The washed bacterial solution was injected into the feed tank, resulting in a system cell concentration of ca. 105 cells/L. The crossflow rate, permeate flow rate, and pressure were maintained at 10 cm/s, 26 μm/s, and 20 psi, respectively. As the bacterial solution circulated through the system, cells deposited on the membrane surface. Pictures of the membrane surface were taken with IPLab software every 2–3 min for 20 min. The bacteria in each picture were counted using ImageJ software and the number of cells deposited were plotted as a function of time. The slope of the resulting line was normalized by the initial cell concentration in the feed and the membrane surface area under the microscope (870 × 729 μm) to calculate the initial deposition rate.
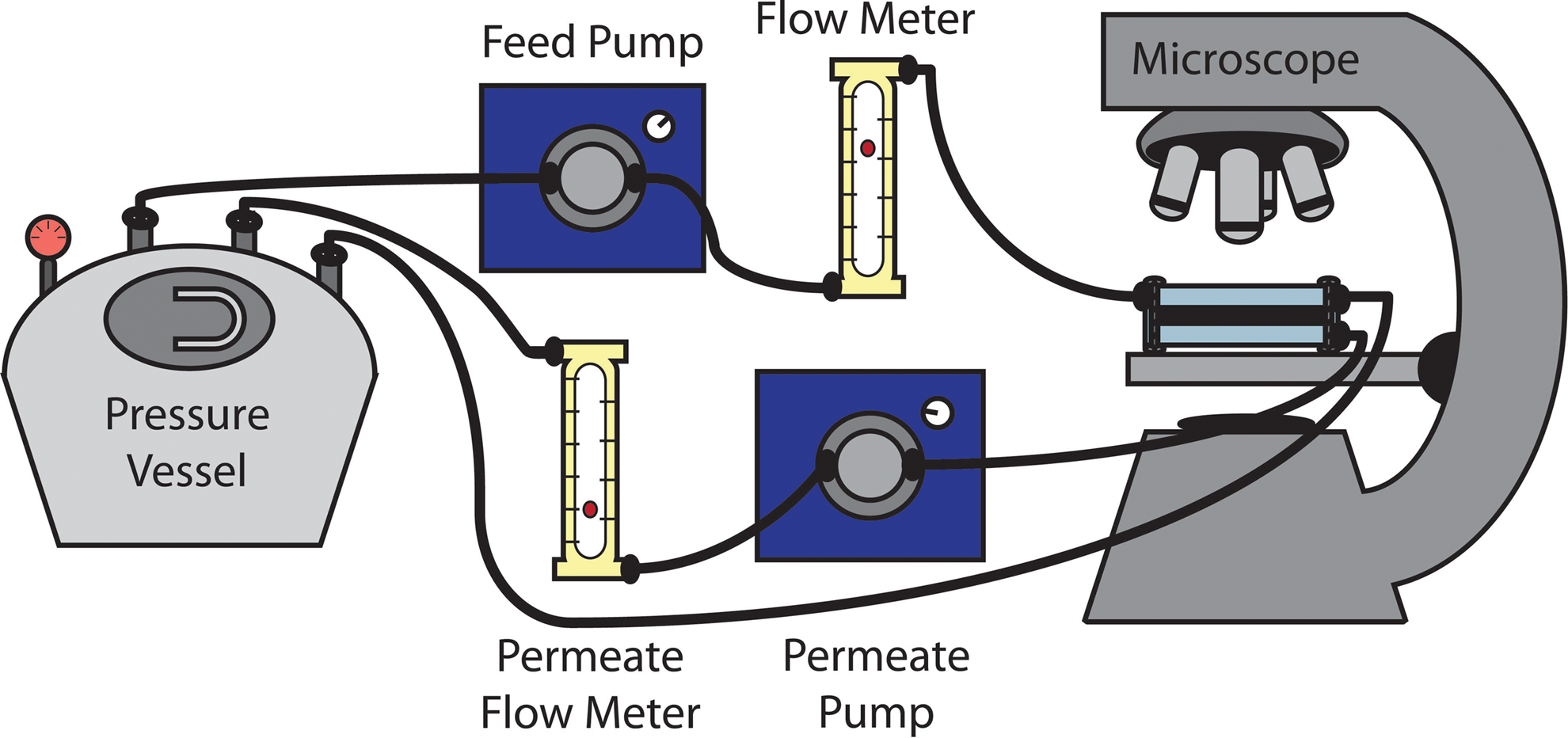
Direct observation system schematic. The system is pressurized to 20 psi. Bacterial feed solution in the pressure vessel is pumped through a flow meter and a polycarbonate flow cell set on the stage of a fluorescent microscope. The polycarbonate flow cell holds the membrane. The crossflow stream runs over the surface of the membrane, while the permeate stream passes through the membrane. Both the crossflow and permeate streams are recirculated to the pressure vessel.
Biofilm formation system
The biofilm formation system (Fig. 2) contains a feed tank, a pump, a flow cell containing the membrane, and crossflow and permeate lines recirculating solution back into the feed tank. The system was cleaned before and after each experiment using the method described by Kwan et al. (2015). Fifteen percent bleach was circulated for 30 min followed by two 10-min rinses with DI water. Then, 5 mM EDTA adjusted to pH 11 was circulated for 30 min followed by two 10-min rinses with DI water. Finally, 95% ethanol was circulated for 60 min followed by three 10-min rinses with DI water. Following cleaning, the membrane was secured in the flow cell, and the pressure and crossflow velocity were set at 25 psi and 8.5 cm/s, respectively. The membrane was subjected to compaction for at least 19 h using 3 L of 200 mg/L NaN3. After compaction, the solution was replaced with 5 L of high nutrient synthetic wastewater to promote bacterial growth. The synthetic wastewater consisted of 0.6 mM Na3C6H5O7, 0.4 mM NH4Cl, 0.2 mM KH2PO4, 0.2 mM CaCl2, 0.5 mM NaHCO3, 8 mM NaCl, and 0.15 mM MgSO4 (Kwan et al., 2015). The system was equilibrated for at least 5 h before bacterial addition. The pressure was then decreased to 5 psi, resulting in an initial flux of 37 L/(m2·h) ±9.7 L/(m·h) (1.2 mL/min ±0.3 mL/min). Initial flux was uncorrelated with membrane type and membrane performance. The washed bacteria were injected into the system resulting in a cell concentration of ca. 105 cells/mL. To determine permeate flux at a time point, the permeate stream was collected for 3 min and weighed. Then, the weight was converted into a volume and divided by the filtration area (membrane area) and the filtration time (3 min). The same experiment was run without the addition of bacteria to isolate the effects of biofouling on membrane flux.
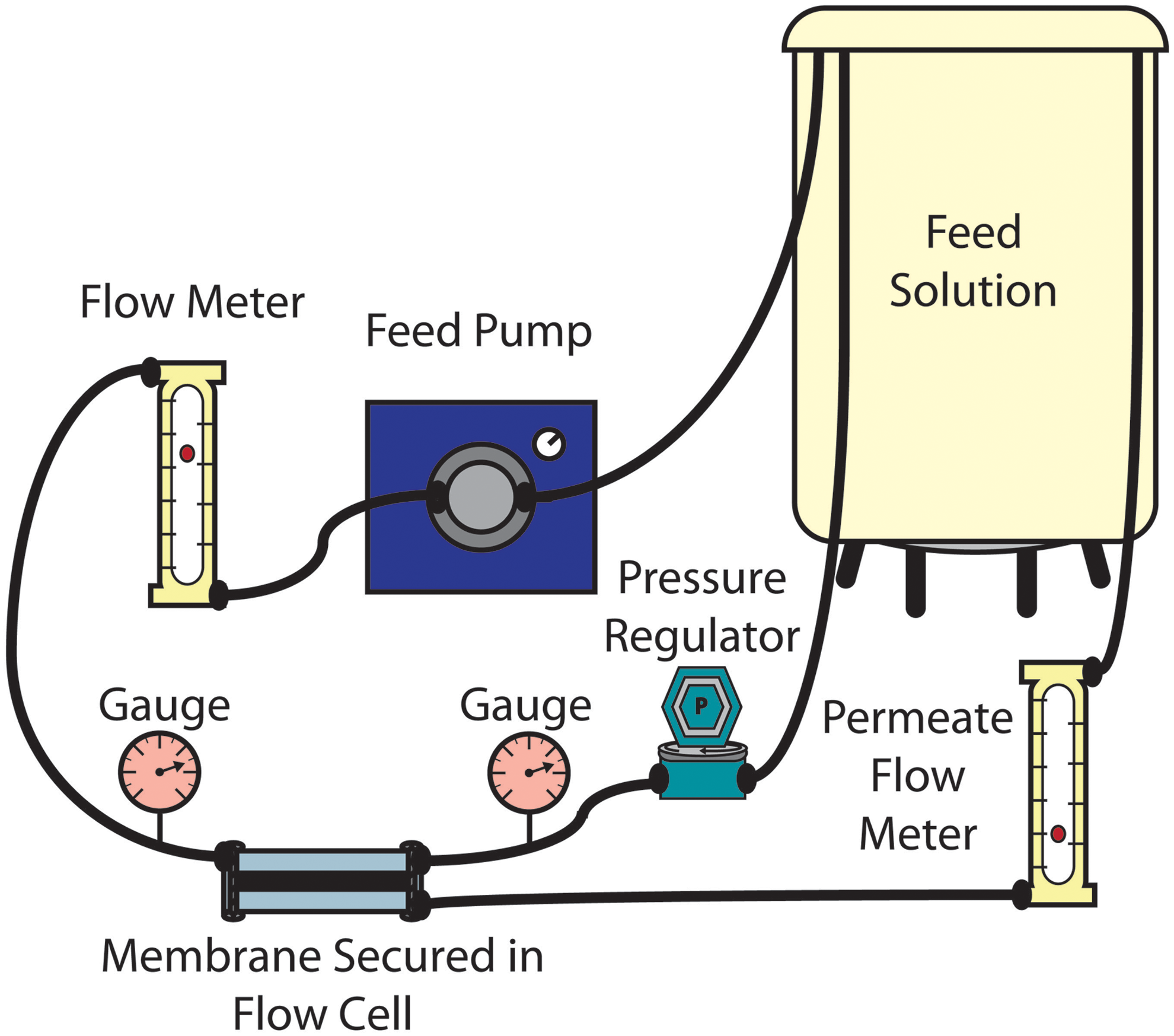
Biofilm formation system schematic. The system is pressurized to 5 psi, maintained using a pressure regulator and two pressure gauges. Bacterial feed solution is pumped through a flow meter and a polycarbonate flow cell which holds the membrane. The crossflow stream runs over the surface of the membrane, while the permeate stream passes through the membrane. Both the crossflow and permeate streams are recirculated to the feed solution container.
Silver leaching
To determine the extent of silver leaching from the PD/Ag-modified membranes over the period of compaction (ca. 19 h of recirculating 3 L of 200 mg/L NaN3) and the period of bacterial filtration (72 h of recirculating 5 L of high nutrient synthetic wastewater), ca. 40 mL of the feed solution was collected and tested for dissolved silver using inductively coupled mass spectrometry (ICP-MS, PerkinElmer ELAN DRC II). The samples were filtered through a 0.2 μm polysulfone membrane (PALL Corporation) and acidified to a 2% HNO3 concentration before ICP-MS analysis. Silver dissolution rate was calculated by dividing the mass of silver dissolved by the total membrane area and the time of exposure.
Biofilm imaging
Following membrane performance evaluation in the biofilm formation system, membranes were removed from the flow cells. The membranes were stained with SYTO 9, Propidium Iodide (Live/Dead Biofilm Viability Kit; Invitrogen), and Concanavalin A (Con A, Alexa Fluor 633; Invitrogen) to visualize live cells, dead cells, and extracellular polysaccharides, respectively. The biofilm was set in place using VECTASHIELD Mounting Medium (Vector Laboratories). Additionally, imaging spacers (Grace Bio-labs) were set between the glass slide and cover slip to protect the sample from compression. These samples were then imaged with a Zeiss LSM 510 Multiphoton ConfoCor 3 laser scanning microscope under a 40 × objective. Image stacks (between three and nine per membrane sample) were taken at an interval of 2 μm and analyzed using COMSTAT 2 to quantify biomass accumulation (Heydorn et al., 2000).
Results and Discussion
Direct observation system
Figure 3a shows the number of bacteria depositing on base, PD-modified, and PD/Ag-modified membranes over a period of 20 min. PD modification decreased the initial rate of bacterial deposition by 48%, whereas the PD/Ag modification decreased the rate by 38% (Fig. 3b). The deposition rates associated with the two modifications were not significantly different at the 95% confidence level, but both modifications resulted in a substantial decline in initial bacterial adhesion compared with the unmodified membrane. The lack of distinction between the PD modification and the PD/Ag modification shows that the hydrophilic PD layer is likely responsible for delaying initial bacterial attachment and is not significantly impacted by the additional incorporation of silver nanoparticles.

This finding is consistent with the literature that has investigated the antiadhesive nature of PD/Ag-modified membranes by quantifying the attraction between the membrane surface and proteins or bacteria. Huang et al. (2016) modified polysulfone ultrafiltration membranes with PD/Ag and characterized their performance during a 70-min laboratory-scale filtration of bovine serum albumin (BSA) solution. Modification increased water flux and demonstrated a small increase in BSA rejection. Our previous work also showed that PD/Ag modification decreased initial deposition of Escherichia coli over a period of 20 min (Tang et al., 2015). Consequently, over short-term exposures, membranes modified with PD/Ag have demonstrated increased resistance to foulant adhesion.
Biofilm formation system
Figure 4 shows the decline in membrane permeate flux over 3 days of P. aeruginosa filtration. The rate of water filtering through the base, PD-modified, and PD/Ag-modified membranes decreased to 18%, 28%, and 33% of the initial values, respectively. These flux losses can be attributed to biofouling as the base membrane used in the control experiment retained 98% of the initial flux (data not shown). The flux of the base and PD/Ag-modified membranes were consistently significantly different throughout filtration. From hours 19 to 72 of filtration, the flux of the PD/Ag-modified membrane remained between 1.77 and 1.98 times greater than that of the unmodified membrane. PD modification by itself, however, did not result in significantly improved membrane performance compared with base membranes at any point during filtration. The antiadhesive nature of PD, therefore, was not enough to significantly alter the effects of biofouling on the membrane surface. The additional modification with antimicrobial silver nanoparticles was required to see an improvement in membrane performance.
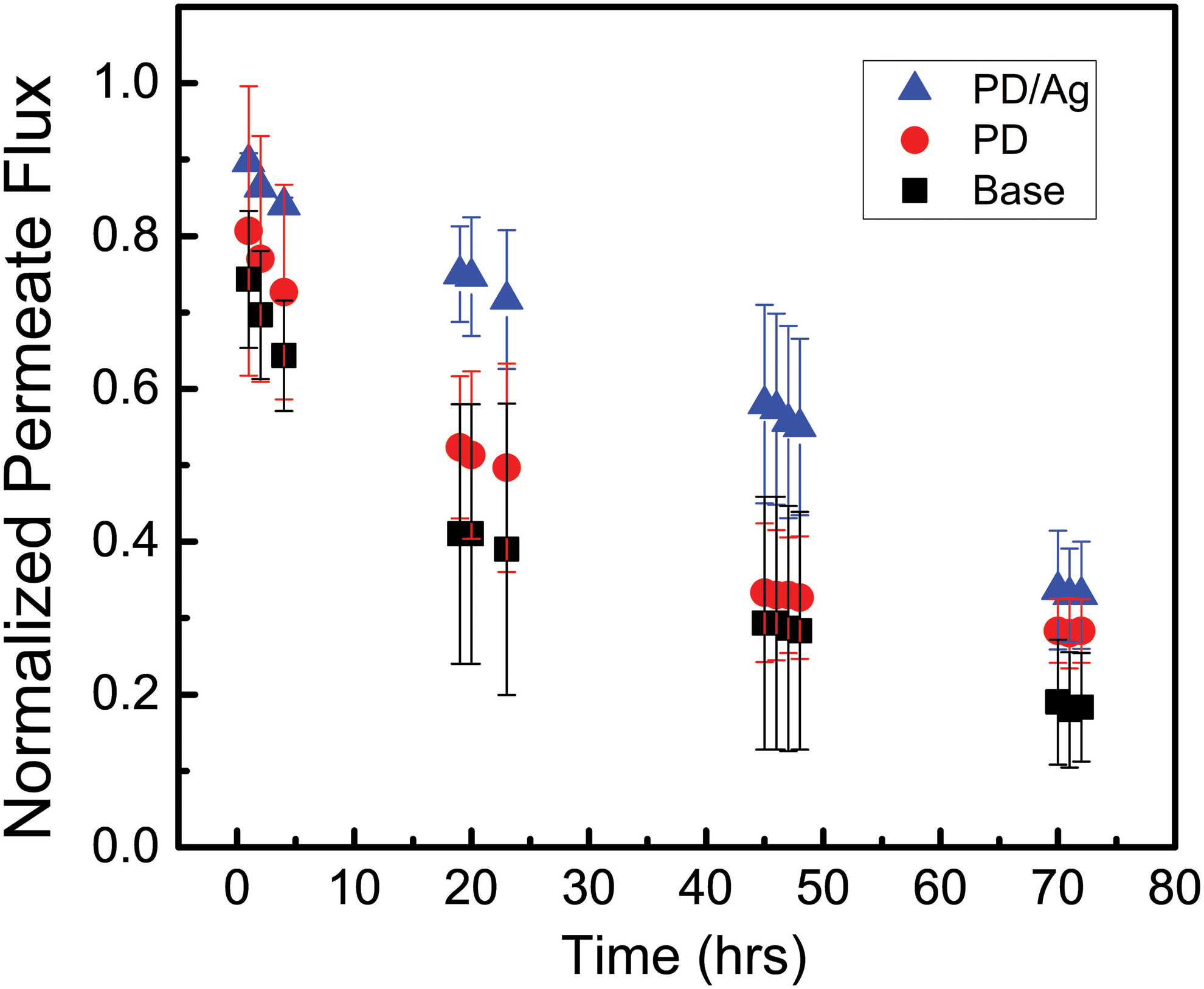
Flux decline over 3 days of P. aeruginosa filtration in the biofilm formation system through base, PD-modified, and PD/Ag-modified membranes. Flux measured by weighing the output of the permeate stream collected over a period of 3 min and converting to a flow rate. Data represent average and standard deviation of triplicate experiments.
This inconsistency in the results of the short and long-term tests supports previous assertions that while hydrophilic membrane modifications can delay initial fouling, they do not provide sufficient protection over longer filtration periods (Xu et al., 2010; Miller et al., 2012). Throughout the literature, hydrophilicity is shown to be one of the primary surface characteristics influencing bacterial adhesion (Pasmore et al., 2001; Geise et al., 2010; Habimana et al., 2014). However, Xu et al. (2010) showed that the improvement in temperature corrected specific flux provided by a hydrophilic, neutral surface was only exhibited during the initial stages of fouling (<48 h). After the initial stages, all membrane types were subject to flux decline due to bacterial attachment. Miller et al. (2012) also found a discrepancy in the impact of hydrophilic modifications on membrane efficiency over short-term and long-term periods of filtration. While the PD and poly (ethylene glycol) modification delayed adhesion after 1 h of exposure, it did not lessen the pressure increase the system experienced during longer-term fouling runs that lasted up to 10 days. Our findings reinforce the conclusion that Miller et al. (2012) drew that accurate assessments of antibiofouling coatings require longer-term filtration studies.
The improvement in membrane performance seen in this study was due to the antibacterial character of PD/Ag-modified membranes. The antibacterial effects of the membrane modification have generally been measured by tests that put bacteria in direct contact with the membrane following modification. The studies that included disk diffusion assays found that silver modification resulted in zones of inhibition surrounding the membrane surface (Sureshkumar et al., 2011; Yang et al., 2012, 2016b; Huang et al., 2016; Zhang et al., 2016). Tang et al. (2015) demonstrated the antimicrobial character of the modification by quantifying the colony-forming units that grew on the membrane surface following filtration. This antibacterial effect was also shown in various studies that used the spread plate method to determine the concentration of bacteria able to survive 2–24 h of incubation with the modified membranes (Huang et al., 2016; Liu and Hu, 2016; Yang et al., 2016b; Zhang et al., 2016). Additionally, Huang et al. (2016) measured the water flux through the membrane before and after incubation with bacterial solution for 48 h. The modification resulted in a higher flux following incubation. All of these studies show that membrane modification results in inactivation of bacteria, but the impact of these findings was unclear as the studies were not conducted in realistic membrane filtration systems, where bacteria is continually pushed to the membrane surface by applied pressure, building up layers over time. This study demonstrates the translation of the antibacterial character of PD/Ag-modified membranes to an improvement in membrane performance in a constant pressure filtration system.
Silver leaching
The total load of silver on membranes modified using this method is ca. 9.6 μg/cm2 (Tang et al., 2015). Triplicate compaction runs yielded silver dissolution rates of 0.22, 0.15, and 0.13 μg/(cm2·d), resulting in an average silver dissolution rate of 0.17 μg/(cm2·d) and a standard deviation of 0.05 μg/(cm2·d). Silver dissolution rates of 0.04, 0.02, and 0.05 μg/(cm2·d) were measured following triplicate bacterial filtration runs, resulting in an average silver dissolution rate of 0.04 μg/(cm2·d) with a standard deviation of 0.02 μg/(cm2·d). The concentrations of silver in the recirculated synthetic wastewater were two orders of magnitude below the silver drinking water standard of 100 μg/L (Ratte, 1999). These dissolution rates, including the overall trend of the rate of dissolution decreasing after the initial silver release, are consistent with those reported in similar studies (Liu and Hu, 2016; Yang et al., 2016b; Zhang et al., 2016). The membranes lost about 1.8% of their silver over 1 day of compaction and 1.3% over the following 3 days of bacterial filtration. Liu and Hu (2016) also demonstrated that reintroduction of silver nitrate results in a recharge of silver nanoparticles on the membrane surface. Given the mild conditions required (ambient temperature and pressure), this modification technique has the potential to be reapplied in situ.
Biofilm imaging
Representative images showing the wide range of bacterial growth that developed on each type of membrane surface are shown in Fig. 5. No clear pattern emerged in these images. The live cell, dead cell, and extracellular polysaccharide biovolumes quantified through analysis of these image stacks using COMSTAT 2 were too varied to be conclusive (Fig. 6). Although extracellular polysaccharide biomass, an indicator of total EPS, is too wide ranging for differences to be statistically significant, the average accumulation on PD/Ag-modified membranes is three orders of magnitude lower than that on base and PD-modified membranes. This disparity suggests that EPS biomass accumulation is most indicative of membrane performance, supported by previous literature, which asserts that it is the viscoelastic property of EPS, rather than the presence of deposited bacteria, which drastically increases the hydraulic resistance of the membrane during biofouling (Baker and Dudley, 1998; Herzberg and Elimelech, 2007). Overall, the biofilm imaging conducted in this study did not reveal a meaningful relationship between membrane performance and biomass accumulation. Literature studies using similar biofilm imaging techniques were examined in an attempt to identify factors contributing to the inconclusive observations reported in this study.
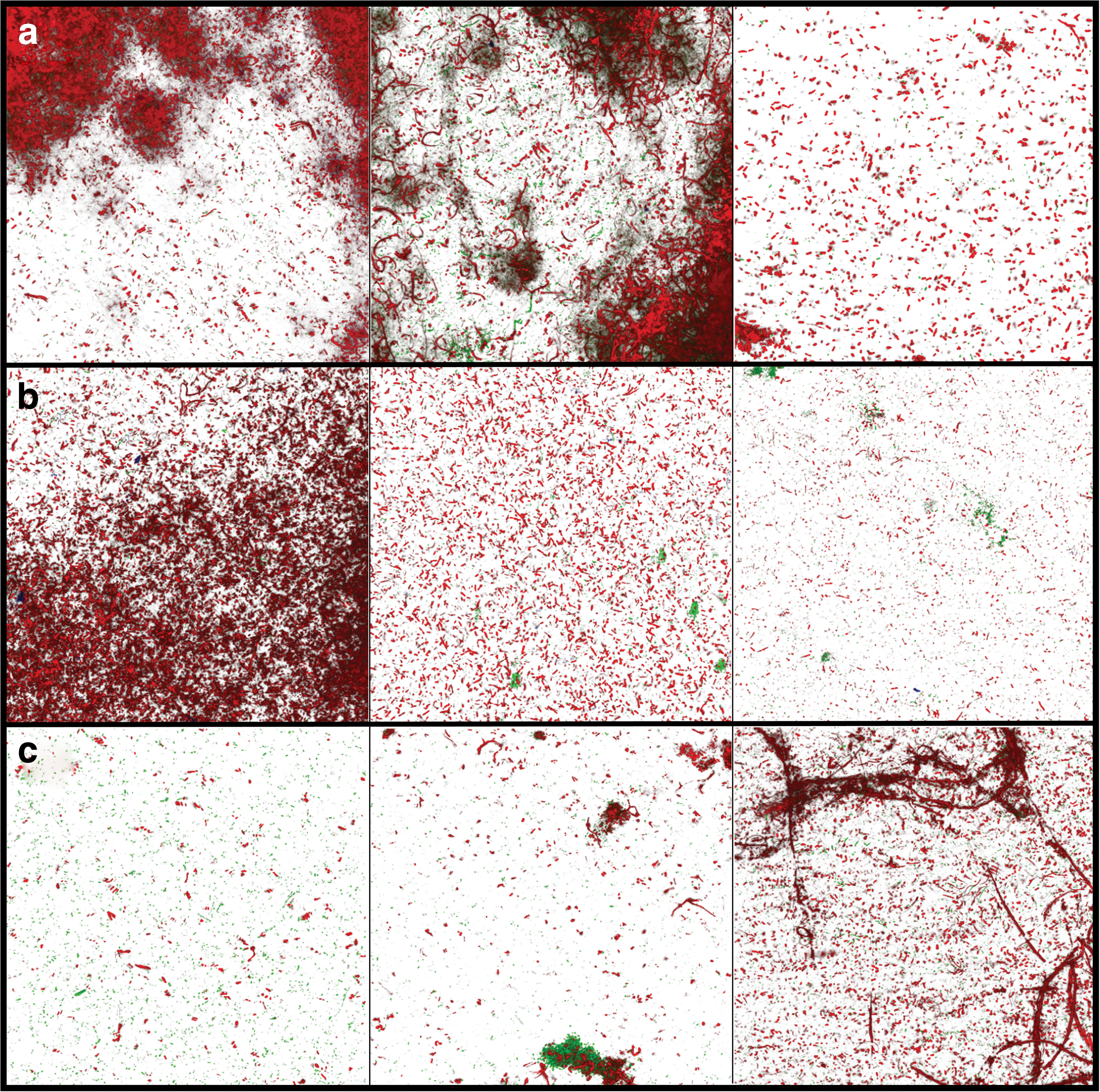
Representative top view confocal images of the wide range of bacterial growth developed on the membrane surfaces over 3 days of bacterial filtration in the biofilm formation system. Live cells (green) and dead cells (red) are shown on
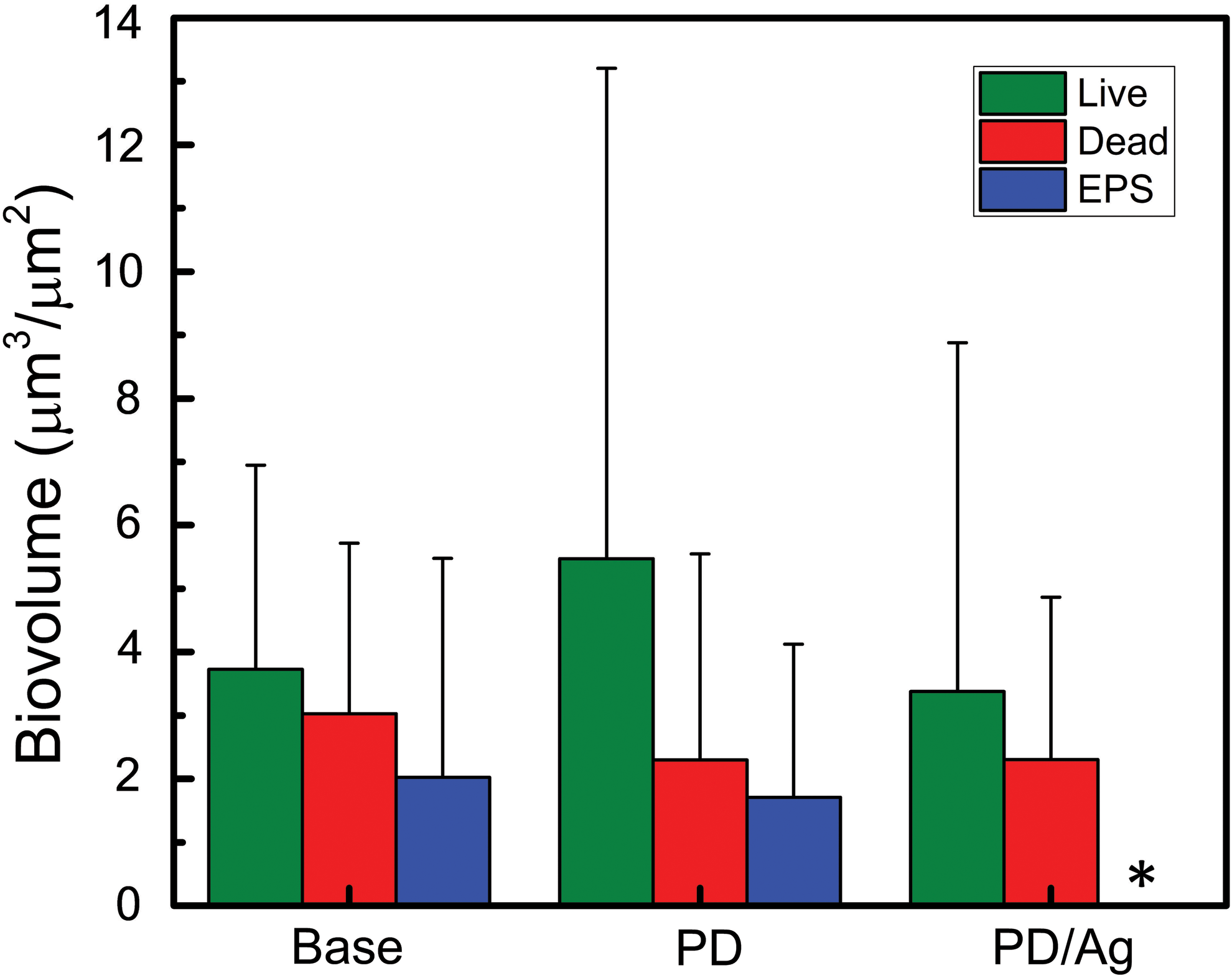
Live cell, dead cell, and extracellular polysaccharide (used as an indicator of EPS) COMSTAT biovolumes measured on base, PD-modified, and PD/Ag-modified membranes. The extracellular polysaccharide biovolume associated with PD/Ag-modified membranes, indicated by an asterisk (*), cannot be seen as the average value is 0.0025 ± 0.003 μm. Data represent average and standard deviation of triplicate experiments.
The biofilm heterogeneity observed in this experiment was not reported in some previous studies that used confocal microscopy to quantify bacterial accumulation on membrane surfaces (Herzberg and Elimelech, 2007; Bar-Zeev et al., 2014; Ben-Sasson et al., 2014; Bernstein et al., 2014; Kwan et al., 2015; Liu et al., 2017). While these studies used Pseudomonas sp. in the feed, other pertinent factors, including membrane type, nutrient conditions, cell concentration, and sample preparation, varied. The active layer of the membranes used in previous studies was polyamide, whereas the membranes in this study are polysulfone. This material change is not likely to account for the differences in bacterial coverage as polyamide membranes are more hydrophilic than polysulfone membranes and thus should be less prone to fouling (Benjamin and Lawler, 2013). Furthermore, Flemming and Schaule (1988) demonstrated that 2–4 h of contact with various bacterial suspensions (Pseudomonas vesicularis, Acinetobacter calcoaceticus, Staphylococcus warneri, and mixed culture) did not result in significantly different attachment to polyamide and polysulfone membranes. Additionally, modifying the membranes with positively charged, negatively charged, or zwitterionic hydrophilic polymers and/or silver nanoparticles (Ben-Sasson et al., 2014; Bernstein et al., 2014; Liu et al., 2017) did not significantly affect the variance in previous studies. The heterogeneity in our study was also consistent between the hydrophobic base and hydrophilic modified membranes. Therefore, it seems unlikely that the membrane surfaces can account for the discrepancy in results.
Nutrient conditions are known to affect the development of the biofilm structure. Flemming and Schaule (1988) showed that starved cells tend to form islands of bacterial growth rather than a smooth homogeneous biofilm. The nutrient composition of the synthetic wastewater used in this study was consistent with those used in Bar-Zeev et al. (2014) and Kwan et al. (2015) Although the initial nutrient conditions were the same, the nutrient availability varied due to differences in the initial cell concentrations and the duration of filtration. The filtration in this study ran 2 days longer than the previous studies, whereas the initial cell concentration was 1–2 orders of magnitude smaller. Even with these considerations, given the high carbon content of the synthetic wastewater, we consider it unlikely that the nutrient concentration would be so depleted as to significantly alter biofilm development.
Initial cell concentration may affect biofilm development independent of nutrient limitations. Previous studies (Ridgway et al., 1984; Flemming and Schaule, 1988) have attempted to determine a range in which there is a linear relationship between the concentration of cells in suspension and the number of cells attached to the membrane surface. Ridgway et al. (1984) found this linear range to exist between 0 cells/mL and 106 cells/mL for Mycobacterium sp. in contact with a cellulose diacetate membrane for 3 h. Flemming and Schaule (1988) reported a higher range, between 106 cells/mL and 108 cells/mL, for Pseudomonas sp. exposed to a polyethersulfone membrane for 4 h. A sharp drop in bacterial adhesion below detection limits was associated with a suspended cell concentration less than 106 cells/mL. Given the comparable bacterial species and membrane type used in our study, the initial concentration of 105 cells/mL (1–4 orders of magnitude below similar studies that saw the formation of homogeneous biofilms (Herzberg and Elimelech, 2007; Bar-Zeev et al., 2014; Ben-Sasson et al., 2014; Bernstein et al., 2014; Kwan et al., 2015; Liu et al., 2017)) may fall below a concentration threshold, resulting in significantly different biofilm growth. However, in our constant pressure system, permeate drag will bring bacteria in contact with the membrane surface, limiting the impact of cell concentration as a factor in bacterial attachment.
Bar-Zeev et al. (2014) demonstrated that confinement of the biofilm results in a compacted biofilm structure. Additionally, they show that a more accurate evaluation of the biofilm can be achieved by utilizing a custom-made chamber to avoid this confinement (Bar-Zeev et al., 2014; Kwan et al., 2015; Liu et al., 2017). In lieu of a custom-made chamber, we placed spacers between the glass slide and the cover slip to avoid compaction of the samples. Although some effects may still be present, we have not seen any evidence to suggest that confinement results in increased biofilm heterogeneity.
The magnification used in this study (40 ×) was higher than those used in the previous studies (20 ×) (Herzberg and Elimelech, 2007; Bar-Zeev et al., 2014; Ben-Sasson et al., 2014; Bernstein et al., 2014; Kwan et al., 2015; Liu et al., 2017). However, Bernstein et al. (2014) included an evaluation of images taken at 40 × magnification in the Supporting Information. By comparing the analysis done on image stacks taken at 40 × and 20 × magnification, Bernstein et al. (2014) found that both image sets demonstrated the same overall trend in biomass attachment. Additionally, increasing the magnification did not result in increased variance in the analysis of the image stacks.
We were unable to use confocal microscopy to distinguish between the mass of live cells, dead cells, and extracellular polysaccharides that accumulated on the different membrane surfaces. The heterogeneity of the biofilms that developed was so great that biovolume could not be related to membrane performance. However, as modifying the membrane with PD/Ag resulted in demonstrably higher flux over the course of P. aeruginosa filtration, we must assume that there are differences in bacterial attachment and growth that we were unsuccessful in capturing through confocal microscopy. Comparison with literature on biofilm imaging did not reveal factors contributing to the inconclusive results obtained in this study.
Conclusion
This study demonstrated the need for researchers investigating membrane modification to use long-term fouling systems (i.e., several days) instead of short-term tests (i.e., minutes to hours). During short-term adhesion experiments, the hydrophilic nature of the polydopamine coating was able to decrease the initial deposition rate of P. aeruginosa by almost 50%. However, this antiadhesive effect did not translate to an improvement in membrane flux over 3 days of bacterial filtration. Instead, it was the antibacterial character of the silver that significantly improved membrane performance in a more realistic system. Future studies should probe the effects of the PD/Ag modification on cleaning efficiency, as the antiadhesive character of the PD coating is likely to improve flux recovery (Pasmore et al., 2002; Huang et al., 2016). Overall, PD/Ag membrane modification shows potential for application in membrane treatment systems. While surface modifications lose their efficacy over time, PD/Ag modification has potential for reapplication in situ due to mild modification conditions and proven recharge ability. The cost/benefit analysis will vary depending on the circumstances of the specific treatment site (scale, water quality, bacterial community) and should be considered in future studies. For water sources, where biofouling is a strong limiting factor in clean water production, such as feed waters with high nutrient conditions, decreased energy costs and frequency of membrane replacement will result in a significant reduction in the overall cost of membrane treatment, making high-quality water more accessible.
Footnotes
Acknowledgments
This work was supported by The National Science Foundation Integrative Graduate Education and Research Traineeship Grant 1069213. The authors thank Michael McCaffery and Erin Pryce (Johns Hopkins University Integrated Imaging Center) for providing training on the confocal microscope. They also acknowledge Dr. Moshe Herzberg (Ben-Gurion University of the Negev) for providing the bacterial species Pseudomonas aeruginosa PA01. The authors are grateful for the laboratory assistance provided by Ayella Maile-Moskowitz and Isabela Sampaio.
Author Disclosure Statement
No competing financial interests exist.