
Abstract
Abstract
This study investigates the influence of dosage of nanoscale zero-valent iron (nZVI) on an immobilized biological autotrophic denitrification reactor. Effects of hydraulic retention time (HRT; 11, 13, and 15 h), initial Fe(II) concentration (10, 20, and 30 mg/L), and dosage of nZVI (2, 4, and 6 mg/L) were analyzed by response surface methodology to determine the effect on denitrification. The highest nitrate removal efficiency of 95% occurred under the following conditions: HRT of 13 h, 21 mg/L Fe(II), and 4 mg/L nZVI. High-throughput sequencing was used to analyze the compositions and structures of the microbial community in the bioreactor at different stages. Addition of nZVI had a significant impact on microbial richness and diversity. In addition, strain CC76 and relevant bacteria were the dominant bacteria for the removal of nitrate in the biological reactor.
Introduction
High nitrate levels are mostly present in the discharge of municipal or industrial wastewater, and in agricultural run-off to water resources (Ghadiri et al., 2017). Once absorbed by the body, nitrate is reduced to nitrite that may combine with hemoglobin to form methemoglobin, seriously reducing the blood oxygen-carrying capacity that has been associated with ailments such as blue baby syndrome (Chiueh et al., 2011). There are many different processes and methodologies used for the removal of nitrates. Currently, bioremediation is considered one of the most economical, promising, efficient, and environmental technologies for nitrate removal. Moreover, adsorption and membrane processes, chemical and catalytic processes, membrane bioreactors, electroreduction, and electrocoagulation, as well as electrodialysis are often used for this purpose (He et al., 2017; Mautner et al., 2017).
In the past few years, nanoscale zero-valent iron (nZVI) particles, which are typically composed of a crystalline α-Fe0 core and an iron oxide/hydroxide shell, are extensively used for removing contaminants by precipitation and nitrates by reduction (Su et al., 2015; Honetschlägerová et al., 2016). Previous studies have shown that Fe(III) reduction to Fe(II) is dominated by iron-reducing bacteria (IRB) under anoxic conditions, which can use Fe(III) oxides as extracellular electron acceptors for their growth, coupled with the reduction of many other electron acceptors such as nitrate (Zhang et al., 2014). Several studies have shown that the combination of IRB and nZVI provided a way to prolong the long-term performance of nZVI by coupling the reduction of Fe(III) to Fe(II) (Yang et al., 2017). The combination of IRB and nZVI provided a way to promote the efficiency of nitrate reduction.
Recently, various immobilization techniques are available for keeping microbes in an appropriate environment that is neither nutrient- nor oxygen-limited (Liu et al., 2016). Alginate is one of the most useful supporting materials for entrapping microbial cells due to its low cost, nontoxicity, biocompatibility, and biodegradability (Kim et al., 2017).
Previous studies in a laboratory environment have determined the nitrate removal efficiency of IRB (Su et al., 2016a). However, individual microbial communities are limited in the application of groundwater treatment. The main objective of this article is to develop an immobilized autotrophic nitrate removal system that couples the IRB with nZVI. In this study, bacterial compositions in an immobilized autotrophic nitrate removal system were analyzed by high-throughput sequencing technology. The purposes of this study were (i) to evaluate the effect of hydraulic retention time (HRT), initial Fe(II) concentration, and dosage of nZVI on nitrate removal efficiency and (ii) to analyze the abundance and diversity of bacteria and to determine the dominant microbial communities on denitrification and iron reduction under varying initial nZVI concentrations.
Materials and Methods
Microorganism and simulated conditions
IRB strain CC76 was isolated from Tang Yu oligotrophic reservoir (Shanxi Province, China). IRB strain CC76 was stocked in a 0.5 glycerol solution at −20°C.
Bacteria were grown in basal medium (Su et al., 2018) as follows (per liter): 0.5 g NaHCO3, 0.1 g NaNO3, 0.1 g KH2PO4, 0.05 g MgSO4·7H2O, 0.05 g CaCl2, and 2 mL trace element (TE). The TE included the reagents (per liter) as follows: 1 g EDTA, 0.5 g MgSO4·7H2O, 0.5 g CuSO4·5H2O, 0.5 g FeSO4·7H2O, 0.2 g ZnSO4, 0.2 g CoCl2·6H2O, and 0.1 g MnCl2·4H2O. In this study, strain CC76 was grown in 500 mL bottles and the pH of the medium was adjusted to 7 with 1 mol/L NaOH or HCl solution. The temperature of the reactor was maintained at 30°C.
Cell immobilization
Sodium alginate (SA) is the major material of the bacteria immobilization system and has to be made at a final concentration of 2%, that is, composition SA (4% m/v) mixed with cell suspension alone, with the cell suspension and nZVI, or with deionized water in 1:1 (v/v) ratio. The resulting solution was then added drop-wise into 2% (m/v) CaCl2 solution to form homogeneous pellets. The formed immobilized granules were immersed in CaCl2 solution at 30°C until use.
Reactor operation
A laboratory-scale experimental setup of an immobilized system with 13 stages (Table 1) was operated for ∼100 days. The experiment contained three parallel systems: (a) control group: immobilized SA pellets with deionized water; (b) experimental group: immobilized SA pellets with strain CC76; and (c) experimental group: immobilized SA pellets with strain CC76 and nZVI powder. At different stages of the experiment, the concentrations of NO3−-N, Fe(II), and NO2−-N in the influent and effluent were measured.
Operational Conditions of Biological Reactor
HRT, hydraulic retention time; nZVI, nanoscale zero-valent iron.
Experimental design for evaluation of optimum conditions
To determine the best conditions of the reactor, the effects of HRT (11, 13, and 15 h), initial Fe(II) concentration (10, 20, and 30 mg/L), and dosage of nZVI (2, 4, and 6 mg/L) on anaerobic denitrification were analyzed. Seventeen experiments were carried out according to the Box–Behnken design. Design-Expert (version 8.06) software was used for statistical design and data analysis.
Analyses of high-throughput sequencing
Samples (LT2, LT3, LT4, and LT5) were collected from the reactor at the influent nZVI concentration of 0 mg/L (stage 7), 2 mg/L (stage 8), 4 mg/L (stage 7), and 6 mg/L (stage 9) among the steady operation period. Respectively, sample LT1 was collected from the control reactor under the same condition. The 16S rRNA gene of strain CC76 was polymerase chain reaction-amplified using the universal bacteria primers F27 (5′-AGAGTTTGATCMTGGCTCAG-3′) and R1492 (5′-TTGGYTACCTTGTTACGACT-3′). The pyrosequencing procedure and the analysis method of sequence were followed according to Hao et al. (2013). In addition, the National Center for Biotechnology Information BLAST and Ribosomal Database Project (RDP)-II Classifier of the RDP were applied to the taxonomy-based analysis.
Analytical methods
Concentrations of nitrate, nitrite, and dissolved Fe (II) were measured using ultraviolet spectrophotometric methods, N-(1-naphthalene)-diaminoethane spectrophotometry, and spectrophotometrically with phenanthroline (DR5000, HACH, USA). The nitrate removal efficiency was determined by the initial concentration of NO3−-N and the final concentration of NO3−-N. pH in this experiment was monitored using a pH meter (HQ11d, HACH, USA).
Results and Discussion
Overall performance of bacteria immobilization system
Overall performance of the bacteria immobilization system with 13 stages is shown in Fig. 1. As shown in Fig. 1a, the concentration of nitrate in group c was significantly lower than that in group b. The maximum removal rate of nitrate in experimental groups b and c was 76% and 93%, respectively, with 4 mg/L nZVI and 20 mg/L Fe(II), and the HRT was 13 h. The concentration of nitrate in groups b and c was 3.7 and 0.74 mg/L, respectively. Nitrate removal was effected by the addition of nZVI due to the reaction between nitrate and nZVI on the metal surface (Zhang et al., 2010). Meanwhile, there was also a certain removal rate of nitrate in control group a, which may be because nitrate acted as a nitrogen source for the growth of other bacteria (Chandra et al., 2016).

The changes in nitrite concentration are shown in Fig. 1b. The accumulation of nitrite-N in group c was significantly higher than that in group b. The maximum nitrite concentration in test group c was 3.8 mg/L, which was due to nZVI reacting with nitrates in the reactor (Ryu et al., 2011). As shown in Fig. 1c, with the addition of nZVI, the concentration of Fe(II) in group c was higher than that in group b and reaches a maximum of 20.39 mg/L in the short term (stage 4). This was because the reaction between nZVI and nitrate resulted in the production of Fe(II). Furthermore, Fe(II) acts as an electron donor for the denitrification process and would be converted to Fe(III) and, after that, part of the Fe(III) would be reduced to Fe(II) by the action of strain CC76. As a result, the concentration of Fe(II) fluctuates, which is in accordance with our previous study on IRB (Su et al., 2016b).
A Box–Behnken design with 17 experiments was conducted to investigate the interactive effects of the three important parameters that significantly affect the removal of nitrate, that is, HRT, dosage of nZVI, and Fe(II) concentration. Figure 2 shows the relationship among HRT, Fe(II) concentration, and dosage of nZVI on the nitrate removal rate. The results indicate that the highest nitrate removal efficiency (95%) occurs with an HRT of 13 h, 21 mg/L Fe(II), and 4 mg/L nZVI, which was consistent with the study mentioned.

Figure 2a shows that the nitrate removal efficiency increased gradually when the HRT increased from 11 to 15 h. A longer HRT may promote the viability of the bacteria and allow them to adapt to the living environment and remove nitrate more effectively. HRT as an external factor had a significant effect on the growth of bacteria. As Xu et al. (2016) have suggested, the extension of HRT would promote the nitrogen removal rate. The response surface methodology (RSM) analysis demonstrated that the nitrate removal rate had a gradual upward trend when the Fe(II) concentration increased from 10 to 21 mg/L, and then decreased with increasing concentration of Fe(II). This indicates that the highest nitrate removal rate was obtained when Fe(II) concentration was 21 mg/L.
Fe(II) as an electron donor at an appropriate concentration can assist in denitrification to enhance nitrogen removal (Zhang et al., 2018). However, excess Fe(II) enters the microbial cells and limits the microbial activity and so the nitrogen removal rate will decrease (Song et al., 2016).
As shown in Fig. 2b, Fe(II) and nZVI concentrations were evaluated to determine their effects on the nitrate removal efficiency. The reactor showed an increasing denitrification efficiency at concentration of Fe(II) from 10 to 21 mg/L and concentration of nZVI from 2 to 4 mg/L. The maximum nitrate removal rate of 95% was obtained at 21 and 4 mg/L nZVI. Fe(II) and nZVI were used as electron donors to remove nitrate (Fig. 3). The removal of nitrate was achieved simultaneously by the reaction of nZVI (Park et al., 2016) and biological denitrification (Campos et al., 2008). Table 1 shows a higher nitrate removal efficiency upon addition of nZVI at all stages, which indicates that nZVI plays a major role in the efficiency of nitrate removal (Oprckal et al., 2017).

Potential mechanisms of the stimulative effects of nZVI and Fe(II) in the biological reactor.
Diversity of microbial communities
Operational taxonomic units (OTUs) is an artificial taxon for analysis in population genetic, and takes the possible sequencing reads based on their sequence and result in clusters of reads (Mysara et al., 2017). The microbial diversity indices listed in Table 2 show the variations in the microbial population in samples LT1–LT5. A total of 34,580, 34,674, 33,134, 32,818, and 28,811 effective sequences were retrieved from the reactors, which were assigned to OTUs at 0.97 similarity. Samples LT1, LT2, LT3, LT4, and LT5 were obtained from different stages of the reactor, with LT1 being sampled for making a contrast. Reads represent the number of sequences in each sample and reflect the community when the read number is large enough (Cheng et al., 2016). The number of OTUs of LT2 and LT3 showed that the number of microorganisms species decreased with the addition of nZVI in the immobilized system, indicating that nZVI was less toxic to strain CC76. Moreover, the changes of additives, especially with more nZVI, led to an increased number of OTUs, indicating that nZVI at higher concentrations can provide a more suitable environment for the bacteria, particularly nZVI can enhance the relative abundance of the bacterial community at low concentration (Li et al., 2010).
Microbial Diversity Indices in Biological System
OTU, operational taxonomic unit.
The values of Ace and Chao revealed a tendency similar to the OTUs with a decrease in community richness (Pazinato et al., 2010). Sequence coverage is a coefficient of concordance of the sequencing depth, which reflects the reliability of the sequencing result (Tang et al., 2016). The coverage given in Table 2 indicates that the collected samples can be a good representation of the experimental situation. The Shannon and Simpson indices are also used to evaluate the microbial community diversity (Schloss et al., 2011). The relevant sample has a higher diversity when the Shannon index is higher, or when the Simpson index is lower. These indices provided relevant information for the related investigation.
The rank–abundance curve is another method that can be used to analyze species abundance and distribution evenness. According to Fig. 4a, the range of species abundance from small to large in the horizontal direction was in the order LT3, LT2, LT1, LT4, and LT5. There was higher species abundance at a high concentration of nZVI (>4 mg/L) and lower species abundance from 0 to 2 mg/L. This indicates that the concentration of nZVI contributed to population diversity in the reactor, and thus, the rank–abundance distribution and the community diversity analysis mentioned interrelate closely.
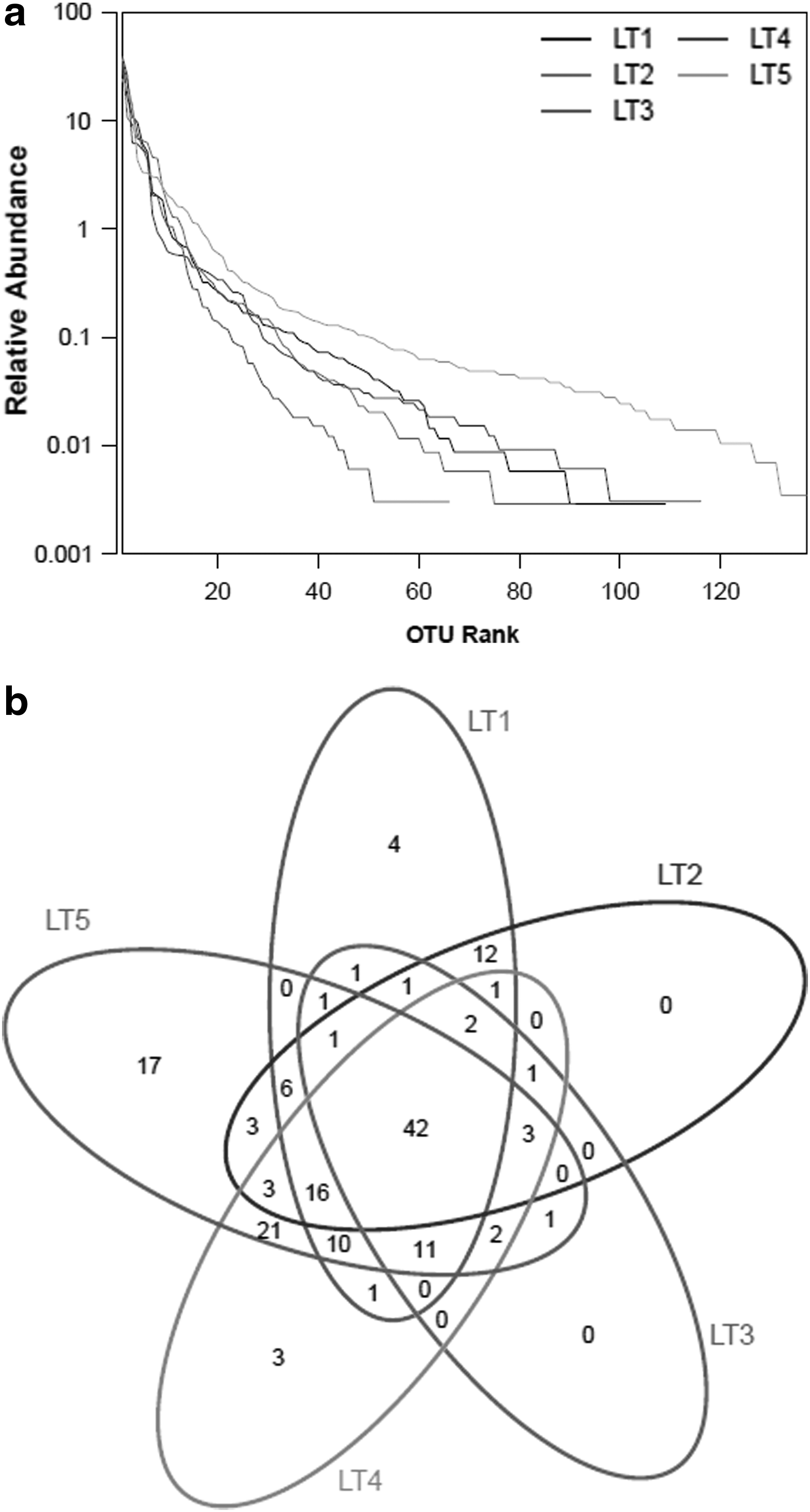
Diversity of bacterial communities in the reactor.
As shown in Fig. 4b, a Venn diagram was adopted to show the similarity and difference of each sample (Kestler et al., 2005). The ratios of unique OTUs accounting for the total OTUs in LT2, LT3, LT4, and LT5 were 0, 0, 0.026, and 0.12, respectively; the common number of OTUs in all these four samples was 17 at 0.03 distance, wherein samples with nZVI of 6 mg/L provided the most common OTUs. Moreover, the common OTUs shared between LT5 and LT4 were 108, significantly higher than the number of OTUs among other samples, showing that some OTUs were obviously rising when nZVI concentration was increased. The conclusion revealed the changes in community composition of the reactor.
Community structure analysis
A taxonomic analysis was performed to identify the bacterial structures and relative abundances of immobilized granule at different concentrations of nZVI at the phylum, class, and genus levels (Fig. 5). Figure 5a shows the abundance of microorganisms at the phylum level, and that in the microbial community of five samples, the dominant phyla were Proteobacteria, Bacteroidetes, Firmicutes, and Actinobacteria. Proteobacteria was the dominant phyla in all experimental samples (LT2–LT5), accounting for 52%, 59%, 47%, and 52%. Kern and Simon (2009) have reported that Proteobacteria plays a vital role in the process of both nitrification and denitrification. Meanwhile, other abundant phyla, such as Bacteroidetes and Firmicutes, also play a role in the denitrification process (Heylen et al., 2006; Wang et al., 2009).
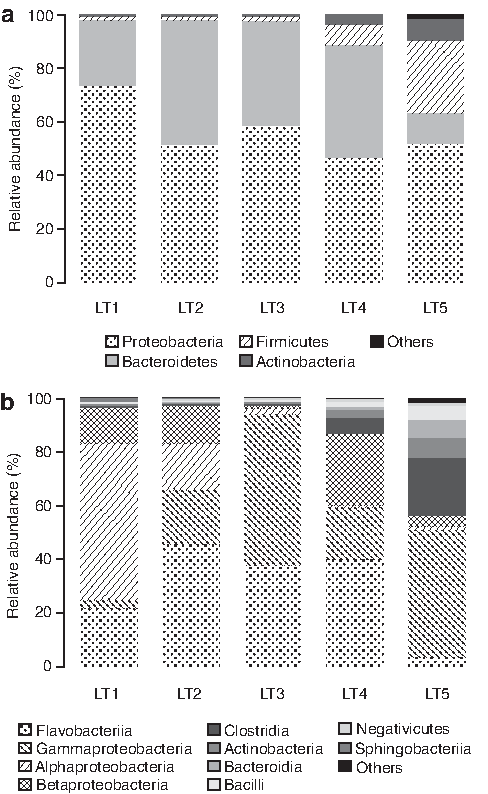
Significant differences were also observed at the class level within samples LT1–LT5 (Fig. 5b). The main classes within LT2 were Flavobacteriia (46%), Gammaproteobacteria (20%), Alphaproteobacteria (18%), and Betaproteobacteria (14%); in sample LT3, Gammaproteobacteria (56%) and Flavobacteiia (38%) were dominant; in sample LT4, Flavobacteriia (40%), Betaproteobacteria (27%), and Gammaproteobacteria (20%) were the prominent groups; whereas in LT5, Gammaproteobacteria (47%) and Clostridia (21%) occupied quite a large proportion among all the bacterial groups; and for the control reactor of sample LT1, Alphaproteobacteria (58%), Flavobacteriia (22%), and Betaproteobacteria (13%) were the major classes. In general, Alphaproteobacteria, Gammaproteobacteria, and Betaproteobacteria belong to the phylum of Proteobacteria; Flavobacteriia pertains to Bacteroidetes; Clostridia is part of the phylum Firmicutes, whereas strain CC76 belongs to Gammaproteobacteria. Many studies have reported that Gammaproteobacteria plays a prominent role in the denitrification process (Gao et al., 2011). As shown in Fig. 5b, Gammaproteobacteria was found to be present in all the samples and accounts for a relatively high proportion.
A heat map categorized at the genus level to analyze bacterial community structures is shown in Fig. 5c. At the genus level, Rhizobium, Flavobacterium, and Sphingomonas were the primary species in control sample LT1; Cloacibacterium and Klebsiella were dominant in sample LT2; Acinetobacter, Klebsiella, and Cloacibacterium were the dominant species in sample LT3, in samples LT4 and LT5, although Cupriavidus and Cloacibacterium were significant in samples LT4 and LT5, the dominant microbial species were Acinetobacter and Klebsiella. The microbial genera Rhizobium, Klebsiella, Acinetobacter, Cupriavidus, and Sphingomonas belong to α-, β-, and γ-Proteobacteria; Cloacibacterium belongs to Bacteroidetes. As Lee et al. (2013) have reported, the enrichment of Bacteroidetes and Proteobacteria plays an important role in the denitrification process.
Conclusions
Addition of an appropriate dosage of nZVI improved the removal of nitrate, but excess nZVI was toxic to the bacteria and had a slight inhibitory effect on the reduction of nitrate.
RSM analysis indicated that the maximum nitrate removal rate (95%) was achieved with an HRT of 13 h, 21 mg/L Fe(II), and 4 mg/L nZVI.
Analysis of microbial distribution and rank–abundance curve indicate that microbial diversity in the reactor was influenced by the addition of nZVI.
Community structure analysis indicate that strain CC76 and relevant bacteria were dominant in the biological reactor.
Footnotes
Acknowledgments
This research work was partly supported by the National Natural Science Foundation of China (No. 51678471), the National Key Research and Development Program of China (2016YFC0400706), Key Scientific Technological Innovation Team Plan of Shaanxi Province (No. 2017KCT-19-02), and Key Research and Development Program in Shaanxi (2018ZDXM-SF-029).
Author Disclosure Statement
No competing financial interests exist.