
Abstract
Abstract
A halotolerant yeast was recently isolated and identified through 26S rDNA and internal transcribed spacer sequencing analysis. Its capacity of decolorizing various azo dyes was investigated, and the dye that was decolorized the fastest was chosen as the target compound for further study. Effect of parameters, including concentrations of nutrients (external carbon, nitrogen, and vitamin mixture), salinity (NaCl concentration), environmental factors (rotation speed, temperature, and pH), and the concentration of the target dye, on decolorization and cell growth of the yeast was investigated. A possible decolorization pathway of the target dye was proposed depending on detection of breakdown byproducts and related literatures. In addition, detoxification effectiveness of the target dye was also estimated through the Microtox test. Results showed that the yeast named SYF-1 was identified as Candida tropicalis and could decolorize six different azo dyes, among which Acid Red B (ARB) was decolorized the fastest, mainly through biodegradation. Growing cells of the yeast SYF-1 displayed the highest decolorization efficiency, with 4.0 g/L glucose, 0.6 g/L ammonium sulfate, 0.08 g/L yeast extract, ≤30 g/L NaCl, rotation speed ≥160 revolutions per minute, temperature 30°C, and pH 7.0–8.0. Both decolorization and cell growth processes were inhibited by higher concentrations of ARB. A degradation pathway of ARB by the yeast SYF-1 was proposed as that consisting of reduction, monooxygenase-oxidization, reductive/oxidative deamination, oxidative desulfurization, and ring-cleavage processes. Moreover, the acute toxicity level of 50 mg/L ARB decreased from original high toxicity to nontoxicity after treatment for 12 h.
Introduction
Nowadays, a large amounts of textile dyes are lost during dyeing processes and then are discharged into industrial effluents every year (Chengalroyen and Dabbs, 2013). Release of these industrial wastewaters to the environment without purification is undesirable due to the potential harms of textile dyes and their breakdown byproducts to humans and other organisms (Ozdemir et al., 2013). Azo dyes, which contain one or more azo groups (–N =.dbnd; N–) in their chemical structures, are the largest class of commercial synthetic dyes representing more than 70% of the total textile dyestuffs (Lang et al., 2013).
In addition to textile processes, azo dyes are also widely used in many other industries, including paper, pharmaceutical, food, cosmetic, and leather (Saratale et al., 2013). These aromatic or heterocyclic compounds are recalcitrant to biodegradation by many microorganisms and are even carcinogenic to humans (Saratale et al., 2011). Therefore, they should be effectively removed from industrial influents before being discharged into the environment.
Many physicochemical and biological treatments are used for treating industrial wastewaters containing azo dyes. For instance, physicochemical methods such as activated carbon adsorption and advanced oxidation processes are potentially efficient for decomposing or even mineralizing azo dyes in wastewaters (Li et al., 2017; Quadrado and Fajardo, 2017). However, these methods are always costly and may cause secondary pollution due to the utilization of reagents (Zhang et al., 2014). By contrast, biological treatments are extensively used due to their advantages, including ecological friendliness, high efficiency, and low cost (Sen et al., 2016). Bacteria and fungi are the most widely researched and used microorganisms for treating azo dyes.
Bacteria, which are characterized by extensive distribution and strong adaptability, are generally efficient for decolorizing azo dyes (Pearce et al., 2003; Dos Santos et al., 2007). However, breakdown byproducts of azo dyes (e.g., aromatic amines) can inhibit the metabolic activity of many bacteria (Tan et al., 2013). By contrast, fungi can further degrade the toxic intermediates of azo dyes into nontoxic compounds depending on their nonspecific ligninolytic enzymes, including lignin peroxidase (LiP), manganese peroxidase (MnP), and laccase (Lac) (Gomi et al., 2011).
Fungi belonging to Basidiomycetes such as Pleurotus sp. and Penicillium sp. have been confirmed as efficient degraders of organic pollutants (Gou et al., 2009; Pozdnyakova et al., 2018). Meanwhile, as a group of single-celled fungi affiliated to Ascomycota, yeasts also display their important roles in treatment of wastewaters from oil manufacturing, fermentation, paper-making, printing, and dyeing industries (Zheng et al., 2001; Yang et al., 2003, 2011, 2012). Further, it has been confirmed that yeasts can degrade and detoxify azo dyes (Tan et al., 2013, 2014). Though fungi have been attracting more and more attention for treating organic pollutants, more efficient and stable fungi are still needed to cope with treatment of novel xenobiotic compounds in industrial effluents.
Wastewaters from textile, petroleum, agro-food, and leather industries generally contain high load of salts aside from organic pollutants (Castillo-Carvajal et al., 2014). High salinity (at least 2% NaCl) can inhibit many microorganisms and result in failure of biological treatments (Oren et al., 1992). However, some microorganisms including Archaea, Bacteria, and Eucarya can survive and keep relatively high metabolic activity under hypersaline conditions, which are identified as halotolerant or halophilic microbes (Oren, 2002). Among them, some halotolerant yeasts belonging to Candida sp., Scheffersomyces sp., and Pichia sp. have been confirmed as efficient degraders of organics, such as azo dyes and phenol under hypersaline conditions (Jiang et al., 2015; Tan et al., 2016; Song et al., 2017).
Though isolation and systematic characterization of halotolerant fungi have been attracting much attention for coping with hypersaline wastewaters, their species are still limited. In addition, specialized microbial communities that contain numerous effective microbes are generally required for field application due to their higher efficiency, stability and environmental adaptability than single strains. Meanwhile, construction of highly effective microbial communities strongly depends on selection of efficient and stable pure cultures. Therefore, more halotolerant fungi (yeasts) are still necessarily explored and studied for this purpose.
The aim of this study was isolation of a halotolerant yeast from a sea mud sample that was capable of degrading and detoxifying azo dyes under hypersaline conditions. The pure culture possessing the highest decolorization efficiency was selected as a research target and first identified through 26S rDNA and internal transcribed spacer (ITS) sequencing analysis. Decolorization effectiveness of six azo dyes by growing cells of the yeast SYF-1 was investigated, and the fastest decolorized one was chosen as target dye. Then, the effects of different parameters on decolorizing the target azo dye by the growing cells of this strain were investigated for selecting the culture medium with the highest efficiency for further testing.
A possible metabolic pathway of the target dye by this strain was proposed based on byproduct identification through high-performance liquid chromatography combined with mass spectrometry (HPLC-MS) technique and related literatures. In addition, acute toxicity to Vibrio fischeri (NRRL B-11177) of the target dye (under hypersaline conditions) before and after treatment by the selected strain was estimated through the Microtox test. It is expected that an efficient and stable halotolerant yeast for treating hypersaline wastewaters would contribute to field application in the near future.
Materials and Methods
Reagents
Azo dyes used in this study were Acid Red B (ARB), Acid Scarlet 3R (3R), Reactive Green KE-4BD (KE-4BD), Reactive Brilliant Red K-2G (K-2G), Reactive Yellow 3RS (3RS), and Reactive Brilliant Blue K-R (K-R), with the characteristic absorption wavelengths of 516, 507, 627, 509, 421, and 565 nm, respectively. They were purchased from Dye Synthesize Laboratory of Dalian University of Technology (Dalian, China) with a purity of more than 99%. Other chemical reagents used in the experiments were purchased from J&K China Chemical Ltd. (Shanghai, China) with analytical-grade purity. In addition, biochemical reagents were purchased from Sangon Biotech Co., Ltd. (Shanghai, China).
Microbial source and culture medium
The sea mud sample for isolation of the azo-decolorizing halotolerant yeast was obtained from the beach of a harbor industrial zone (Dalian, China). The medium for acclimatization of the sea mud microbial community contains (g/L): glucose 4.0, (NH4)2SO4 1.0, K2HPO4 1.0, MgSO4 · 7H2O 0.5, NaCl 30.0, and azo dye 0.02 (initial pH was adjusted to 7.0), and that for pure culture isolation (on solid medium plate) contains 2% (w/v) agar in addition to the components described earlier. Before inoculation, the medium was autoclaved at 115°C for 15 min. After acclimatization for a period, the dye that was decolorized the fastest was first chosen as the target compound for pure culture isolation.
Isolation and identification of halotolerant yeast
The sea mud microbial community was first acclimatized in 250-mL shaking flasks under the conditions of temperature 30°C and rotation speed 160 revolutions per minute (rpm) for about 2 weeks. Then, pure yeast strains were isolated from the acclimatized microbial community by using the dilution-plate method, as previously described (Tan et al., 2016). The pure strain with the highest decolorization efficiency was chosen for identification and further investigation.
Genomic DNA of the chosen strain was extracted and purified; then, the 26S rDNA and ITS sequences were amplified through polymerase chain reaction (PCR) following the previous reports (Tan et al., 2013, 2014). The PCR products were sequenced by Sangon Biotech Co., Ltd. Sequencing results were analyzed through the BLAST program on NCBI database (https://blast.ncbi.nlm.nih.gov/Blast.cgi), and phylogenetic tree was constructed based on the 26S rDNA sequence according to a previous method (Tan et al., 2013).
Azo dyes decolorzation by growing cells of pure yeast
Decolorization of different azo dyes by growing cells of the pure yeast strain was operated in 250-mL shaking flasks with 100-mL volume. Fresh cell suspension of the yeast in the exponential growth phase (cell concentration represented by optical density at 600 nm (OD600) was about 0.375 after 10 times dilution) was inoculated in 100-mL sterilized medium containing 20 mg/L different azo dyes with the inoculation size of 6% (v/v); then, it was incubated at 30°C and 160 rpm for 24 h. To quantify the color removal percentages through adsorption by the cells of the yeast SYF-1, the test on decolorization of 20 mg/L corresponding azo dyes by dead cells (the initial OD600 was the same with the inoculum, pretreated through autoclaved at 121°C for 20 min) was performed.
The azo dye that was decolorized the fastest by the pure culture through biodegradation was chosen as the target compound for further study. The effect of different parameters, including concentrations of glucose (0–6.0 g/L), (NH4)2SO4 (0–1.2 g/L), yeast extract (0–0.1 g/L), and NaCl (0–110.0 g/L), rotation speed (0–200 rpm), temperature (20–40°C), pH (3.0–11.0), and the target dye concentration (50–300 mg/L), on decolorization efficiency and cell growth of the pure yeast were investigated through single-factor tests. When investigating one of these parameters, others were as follows: 4.0 g/L glucose, 0.6 g/L (NH4)2SO4, 0.08 g/L yeast extract, 50 mg/L the target dye, 30.0 g/L NaCl, rotation speed 160 rpm, temperature 30°C, and pH 7.0.
Analytical methods
Concentrations of azo dyes were analyzed by spectrophotometry (JASCO V-560; Japan) after pretreatment through centrifugation (10,000 g for 10 min) for separation of suspended solid. Decolorization ratio was defined as (A0 − A1)/A0 × 100, where A0 and A1 were the absorbance of azo dyes at the corresponding characteristic absorption wavelength before and after decolorization, respectively. Cell density of the yeast was represented by OD600, which was also analyzed by spectrophotometry. To eliminate the influence of possible residual absorbance on analysis of OD600, the supernatant after centrifugation (10,000 g for 10 min) at the same time was used as the reference solution.
To propose the decolorization pathway of the target dye by the pure yeast, the HPLC-MS method was used to identify possible intermediates. Operation conditions of HPLC-MS were the same as those in previous research (Tan et al., 2016). All the analysis was performed in triplicate.
Acute toxicity assessment
Acute toxicity of the target dye and its decolorization byproducts (with the same salinity of 30 g/L NaCl) were assessed by using the Microtox test with luminescent bacterium V. fischeri (NRRL B-11177) according to Tan et al. (2016). Acute toxicity was represented by the inhibition ratio (IR) of luminescence by: IR (%) = (S0 × Cf − St)/(S0 × Cf), where S0 and St were the luminescence intensities of the samples at 0 and t min, respectively; Cf was the relative luminescence intensity variation of the negative sample at 0 to t min.
Statistical analysis
Statistic analysis of experimental data was performed through one-way analysis of variance by using Microsoft Excel 2010 software. The p-value of <0.05 suggests that the data were significant.
Results and Discussion
Identification and basic characteristics of halotolerant yeast
A halotolerant yeast strain named SYF-1, which was capable of decolorizing various azo dyes under aerobic and hypersaline (30 g/L NaCl) conditions, was isolated. Colonies of the yeast SYF-1 were white, smooth on the surface, and regular on the edge, which could also decolorize 20 mg/L ARB on a solid agar plate (Fig. 1C). Meanwhile, it was observed through a scanning electron microscope that the cell shape of this yeast was a short rod (Fig. 1A). A 585 bp partial 26S rDNA sequence of the yeast SYF-1 was yielded and then deposited in GenBank database with an accession number MG751324 (Fig. 1D).
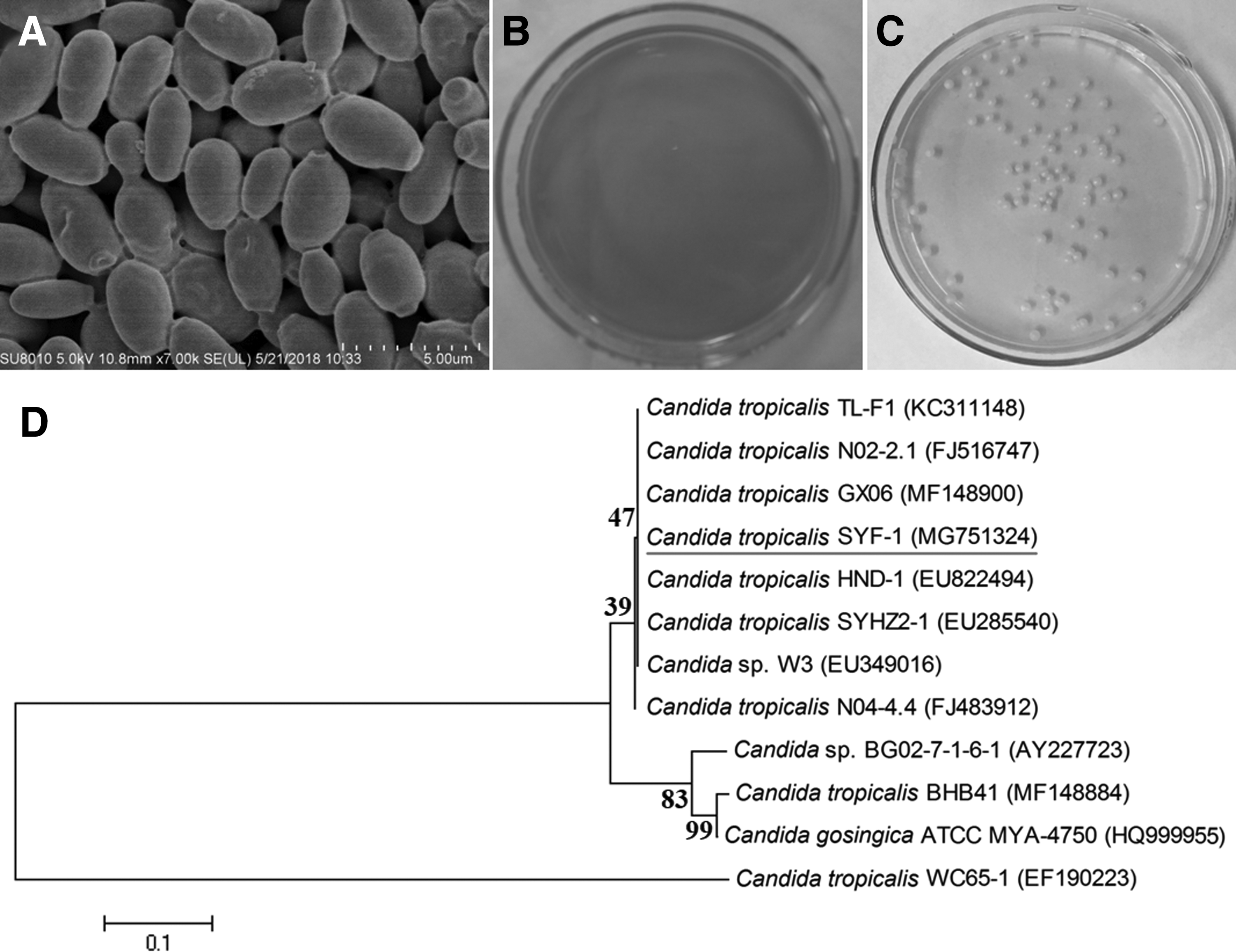
According to the phylogenetic tree, the yeast SYF-1 exhibited 100% homology to several Candida tropicalis strains such as GX06 (MF148900), HND-1 (EU822494), and SYHZ2-1 (EU285540). In addition, the yeast strain SYF-1 was also identified by ITS sequencing and the result (Supplementary Data S1) showed that it also exhibited 100% homology to the C. tropicalis strain CTR1146 (KX664627). Therefore, it was identified as C. tropicalis.
Under aerobic and hypersaline (30 g/L NaCl) conditions, growing cells of the yeast SYF-1 could decolorize 20 mg/L of six different azo dyes with higher than 80% color removal percentages within 12–24 h (shown in Fig. 2). Among the tested azo dyes, KE-4BD was the fastest decolorized dye; however, some of the dye might be removed from the aqueous phase through adsorption on cells of the yeast SYF-1, which could be judged from the color of suspension and cells after decolorization (Fig. 3). Meanwhile, 3R, K-2G, 3RS, and K-R were also decolorized, at least partially through adsorption.
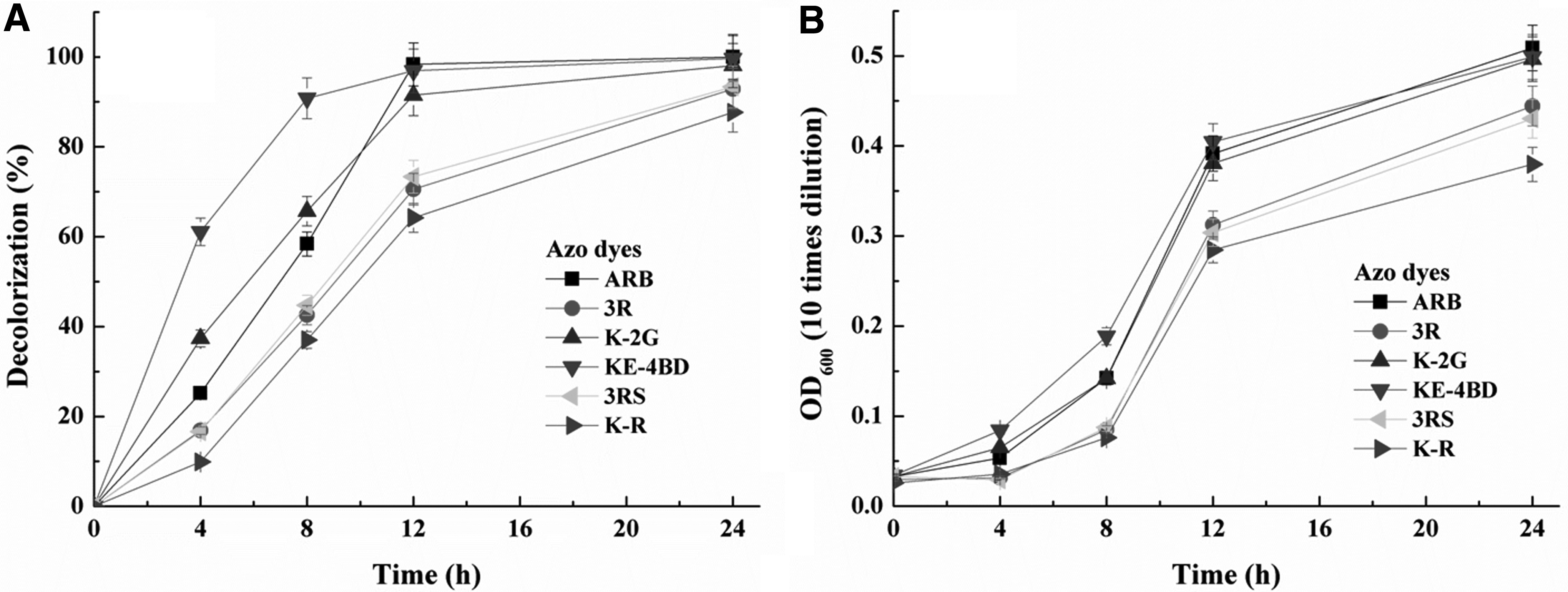
Decolorization of six different azo dyes (20 mg/L) by growing cells of the yeast SYF-1.

Photos of original 20 mg/L dye solution
The result of the adsorption test (Supplementary Fig. S1) showed that 20 mg/L of six azo dyes were adsorbed on the dead cells of the yeast SYF-1, with the maximum (almost saturated) decolorization percentages of 49.1–99.7% within 1 h. Among them, ARB and 3R were decolorized by the growing cells of the yeast SYF-1, with the maximum decolorization percentages of 99.9% and 92.9% within 24 h, which were higher than those by the dead cells (85.8% and 49.1%), respectively.
Meanwhile, the color of the dead cells after adsorbing ARB and 3R (Supplementary Fig. S2) was obviously deeper than that of growing cells shown in Fig. 3. It was suggested that ARB and 3R were decolorized mainly through biodegradation. On the other hand, the maximum decolorization percentages of the other four dyes by growing cells of the yeast SYF-1 were almost the same with the corresponding ones by dead cells, suggesting that the other four dyes, including K-2G, 3RS, K-R, and KE-4BD, were decolorized mainly through adsorption. In comparison, the decolorization (through biodegradation) percentage of ARB within 12 h (98.3%) was the highest among all the six tested dyes; thus, ARB was selected as the target dye for further study.
According to the existing literatures, many strains belonging to C. tropicalis had been identified as efficient degraders of organics such as phenol, aniline, toluene, and azo dyes (Wang et al., 2011; Song et al., 2012; Tan et al., 2013; Basak et al., 2014). However, none of them was halophilic or halotolerant. In addition, some yeasts belonging to genus Candida were confirmed as halotolerant yeasts (Crognale et al., 2012; Jiang et al., 2015; Li et al., 2018). As far as it is known, this might be the first report on the halotolerant C. tropicalis strain that was capable of decolorizing azo dyes. As discussed earlier, exploration of more halotolerant or halophilic microorganisms was important for improving biological treatments of hypersaline wastewaters due to the higher efficiency and stability under hypersaline conditions than nonhalotolerant microbes.
Therefore, the significance of this study was to provide an alternative halotolerant yeast for further field applications.
Effect of parameters on ARB decolorization and growth of the yeast SYF-1
In general, external carbon, nitrogen, and vitamin mixture were essential factors for metabolism and growth of yeasts (Tan et al., 2016). Thus, the effects of their concentrations on ARB decolorization by growing cells of the yeast SYF-1 were performed and the results are shown in Figs. 4A–C and 5A–C. It was suggested from Figs. 4A and 5B that higher than 95% of decolorization percentages (50 mg/L ARB) within 12 h were achieved with at least 4.0 g/L glucose and 0.6 g/L (NH4)2SO4 in the medium. When concentrations of glucose and (NH4)2SO4 further increased, the decolorization efficiency almost no longer increased. However, decolorization percentages were all <85% when concentrations of glucose and (NH4)2SO4 were lower than 4.0 and 0.6 g/L, respectively.
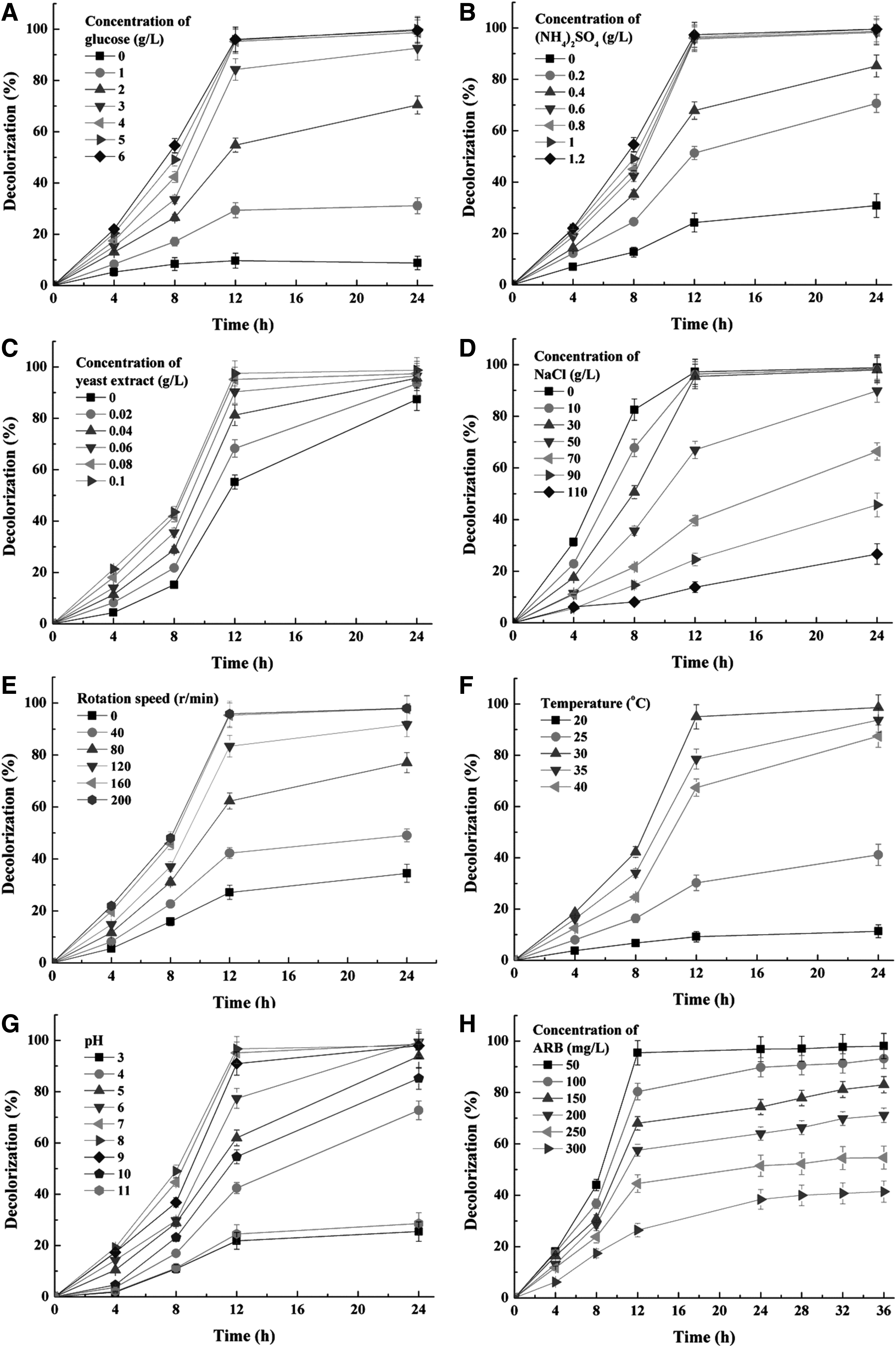
Effect of different parameters on decolorizing 50 mg/L [except for

Effect of different parameters on cell growth of yeast SYF-1.
In addition, at least 0.08 g/L yeast extract was also required for higher than 95% of decolorization percentages within 12 h; in comparison, <91% of the color was removed with 0–0.06 g/L yeast extract (Fig. 4C). On the other hand, higher concentrations of glucose, (NH4)2SO4, and yeast extract also resulted in faster growth of the yeast SYF-1, as shown in Figs. 5A–C. However, the growth curves with 4.0–6.0 g/L glucose [or 0.6–1.2 g/L (NH4)2SO4 or 0.08–0.10 g/L yeast extract] were very close to each other, which were all higher than those with less glucose [or (NH4)2SO4 or yeast extract]. Therefore, the optimal concentrations of glucose, (NH4)2SO4, and yeast extract for biodecolorization were determined as 4.0, 0.6, and 0.08 g/L, respectively.
Effects of salinity (NaCl concentration) on decolorization and cell growth are displayed in Figs. 4D and 5D. It was shown that higher than 95% of decolorization percentages within 12 h were achieved with 0–30.0 g/L NaCl. When NaCl concentration was 50.0 g/L, the 12-h decolorization percentage decreased to 89.9%. When NaCl concentration further increased to 70.0–110.0 g/L, the 12-h decolorization percentages sharply decreased to 26.7–66.4%. It was suggested that metabolic activity of the yeast SYF-1 was inhibited by high concentrations of salt. Meanwhile, high salinity also inhibited the growth of the yeast SYF-1, which was reflected in Fig. 5D. When NaCl concentration was 0–30.0 g/L, the 12-h OD600 was 0.387–0.449.
In comparison, the 12-h OD600 was 0.238 when NaCl concentration increased to 50.0 g/L, which was about one half of that with 0 g/L NaCl. Further, when NaCl concentration further increased to 70.0 g/L, the 12-h OD600 sharply decreased to 0.112, about one quarter of that with 0 g/L NaCl. As mentioned earlier, both halophilic and halotolerant microorganisms could keep relatively higher metabolic activity and growth rate than nonhalotolerant ones under hypersaline conditions. It was indicated that halotolerant microorganisms could tolerate hypersaline conditions; however, they were generally inhibited by high salinity (Tan et al., 2016). In comparison, halophilic ones always grew and metabolized obligately with high salinities (Oren, 2010).
The results described earlier suggested that the yeast SYF-1 could tolerate high salinity of 30.0 g/L NaCl; however, it did not only keep higher metabolic activity under hypersaline conditions. Thus, it could be concluded that the yeast SYF-1 was a halotolerant yeast rather than a halophilic one.
The effect of some common environmental factors, including rotation speed (related to dissolved oxygen), temperature, and pH on the yeast SYF-1, is displayed in Figs. 4E–G and 5E–G. It was shown that higher than 95% of decolorization percentages within 12 h were achieved only with ≥160 rpm rotation speed, 30°C, and pH 7.0–8.0. Meanwhile, cell growth rates under these conditions were also higher than those with the corresponding values out of these ranges. Thus, the optimal rotation speed, temperature, and pH were determined as 160 rpm, 30°C, and 7.0–8.0, respectively.
Finally, the effect of initial ARB concentration was investigated and the results are shown in Figs. 4H and 5H. When the initial ARB concentration increased from 50 to 100 mg/L, the 12-h decolorization percentage decreased from about 95.5% to 80.4%. Meanwhile, the 12-h decolorization percentages of 150, 200, 250, and 300 mg/L ARB were about 68.0%, 57.5%, 44.5%, and 26.4%, respectively. It seems that the metabolic activity of the yeast SYF-1 was inhibited by a higher concentration of ARB. However, the total 12-h decolorization amount of ARB increased from 47.45 to about 115.03 mg/L when the initial ARB concentration increased from 50 to 200 mg/L; then, it started to decrease when the initial ARB concentration further increased to 250 and 300 mg/L.
Thus, it was suggested that ARB of higher than 200 mg/L could inhibit decolorization processes by growing cells of the yeast SYF-1, possibly due to the lack of a carbon source and possible accumulation of toxic byproducts (Pearce et al., 2003). On the other hand, it was suggested from the result in Fig. 5H that cell growth processes were inhibited by ARB of higher concentrations. Further, when time was prolonged from 12 to 36 h, decolorization percentages increased from 26.4–68.0% to 41.4–83.0% corresponding to initial ARB concentrations of 150–300 mg/L. Meanwhile, growth of the yeast SYF-1 also sustained. These results suggested that metabolic and growth activities were not completely inhibited.
Proposed decolorization pathway of ARB by yeast SYF-1
To propose the possible decolorization pathway of ARB by growing cells of the yeast SYF-1, byproducts after treatment were first analyzed through UV-Vis scanning for preliminarily monitoring the change of major functional groups. As shown in Fig. 6A, the intensities at 516 and 323 nm obviously decreased after treatment by the yeast SYF-1 for 12 h, which indicated the destruction of the azo bond and naphthyl ring, respectively. This process might be catalyzed by NADH-dichloroindophenol oxidoreductase and/or Lac (Tan et al., 2014, 2016). Meanwhile, the intensity at 254 nm, which was the characteristic absorption of the benzene ring, first increased after treatment for 6 h, and it then conversely decreased during the next 6 h.

It was suggested that some aromatic compounds were produced and accumulated at the beginning, and then some of them were further degraded by the yeast SYF-1 mainly depending on the catalysis of LiP and/or MnP (Tan et al., 2016; Song et al., 2017). To further reveal the decolorization pathway of ARB, possible byproducts were detected by the HPLC-MS method. The result in Fig. 6B showed that five possible decolorization byproducts were determined as 4-hydrazinylnaphthalene-1-sulfonic acid (compound I), 4-hydroxynaphthalene-1-sulfonic acid (compound II), 4-aminonaphthalene-1-sulfonic acid (compound III), 3,4-dihydroxynaphthalene-1-sulfonic acid (compound IV), and 4-hydroxynaphthalene-1,2-dione (compound VII), with the m/z ion peaks of 237.1, 223.1, 222.1, 239.1, and 173.2, respectively (Arun Prasad et al., 2013; Adnan et al., 2015; Tan et al., 2016).
Based on the analysis results described earlier, possible degradation pathways of ARB by the yeast SYF-1 are proposed, as shown in Fig. 7. Reductive cleavage of azo bond (–N =.dbnd; N–) was generally considered the first step of azo dyes biodecolorization, during which corresponding amines would be produced (Dos Santos et al., 2007). However, neither of the two possible amines, including 4-amino-naphthalene-1-sulfonic acid and 3-amino-4-hydroxy-naphthalene-1-sulfonic acid, was detected in this study, which suggested that the yeast SYF-1 might decolorize ARB through other pathways. Two other possible secondary decolorization byproducts of ARB, 4-hydrazinylnaphthalene-1-sulfonic acid (compound I) and 4-hydroxynaphthalene-1-sulfonic acid (compound II), were detected according to the research by Adnan et al. (2015).
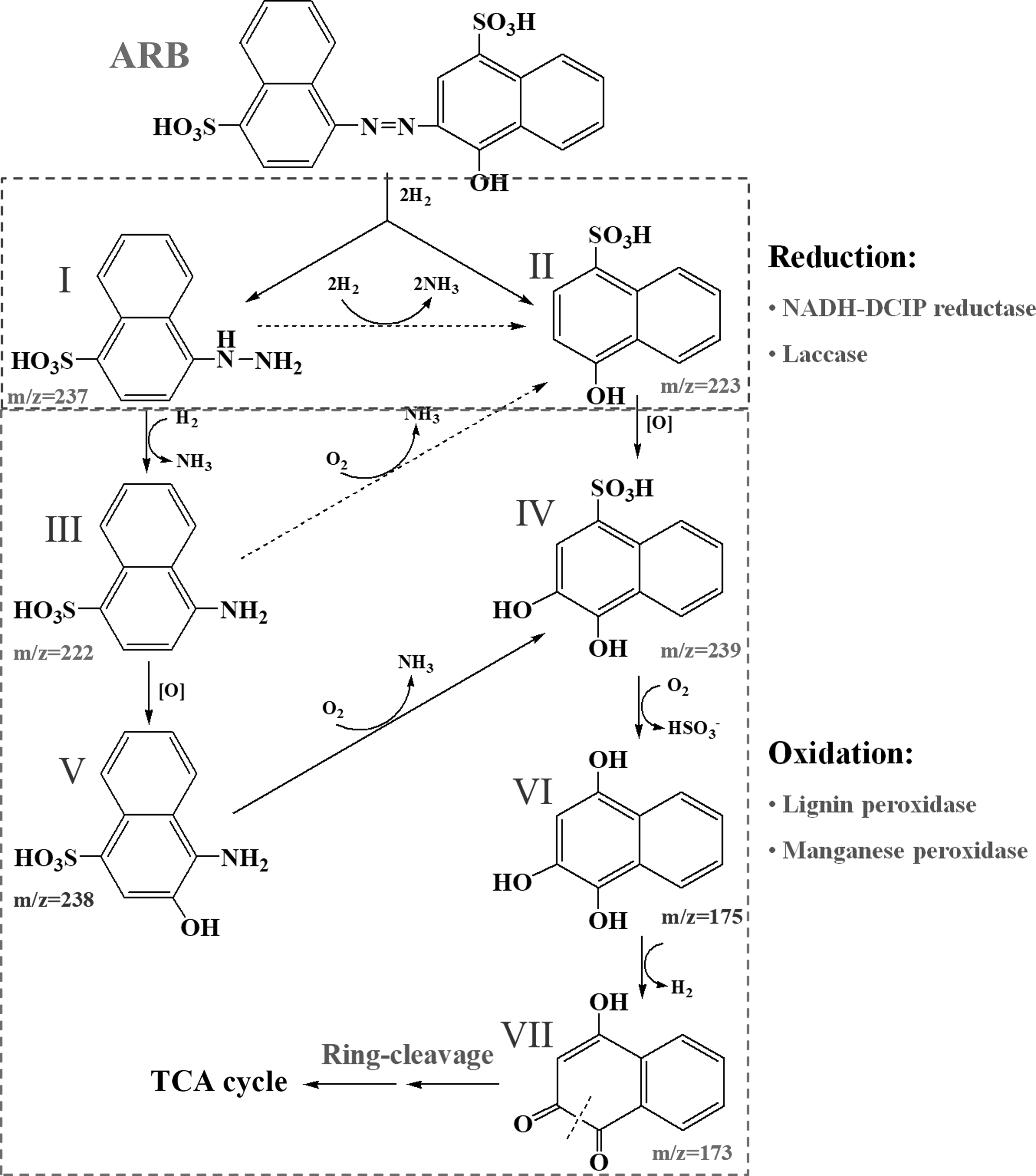
Proposed partial pathways for ARB degradation by growing cells of yeast SYF-1. I, 4-hydrazinylnaphthalene-1-sulfonic acid; II, 4-hydroxynaphthalene-1-sulfonic acid; III, 4-aminonaphthalene-1-sulfonic acid; IV, 3,4-dihydroxynaphthalene-1-sulfonic acid; V, 4-amino-3-hydroxynaphthalene-1-sulfonic acid; VI, naphthalene-1,2,4-triol; VII, 4-hydroxynaphthalene-1,2-dione.
Then, compound I might be successively degraded into compounds III (also detected) and II through reductive and oxidative deamination processes, respectively (Saratale et al., 2013). Meanwhile, compound III might also be transformed into compound V through a oxidative bioprocess catalyzed by monooxygenase (Song et al., 2017). Then, compounds II and V might be degraded into the same product (compound IV, detected) through monooxygenase-oxidative and oxidative deamination processes, respectively (Song et al., 2017). Subsequently, compound IV might be successively degraded into compounds VI and VII through oxidative desulfurization and reduction processes, respectively, which were proposed according to the detection of compound VII.
Finally, compound VII might be further degraded into small-molecular products through a series of ring-cleavage processes and tricarboxylic acid cycle (TCA) cycle. The proposal of decolorization pathways depended on the determination of possible byproducts and related reports, which might be not exact in some details. Thus, exact decolorization mechanisms should be further systematically investigated in future research.
Assessment of acute toxicity
To evaluate the detoxification effect of ARB and its toxic decolorization byproducts by the yeast SYF-1, the acute toxicity of 50 mg/L ARB (along with 30 g/L NaCl) before and after treatment (for 6 and 12 h) under the optimal conditions was estimated and compared by the Microtox test. The IR of about 75.4% against V. fischeri after exposure for 5 min suggested that the original 50 mg/L ARB possessed high acute toxicity (Fig. 8). After treatment for 6 h, the dye solution also displayed the 5-min IR of about 67.2%, just slightly lower than that before treatment. However, the 5-min IR of the dye solution sharply decreased to <10% (9.4%) after complete decolorization for 12 h, suggesting a nontoxic level. This result suggested that the yeast SYF-1 could detoxify ARB.
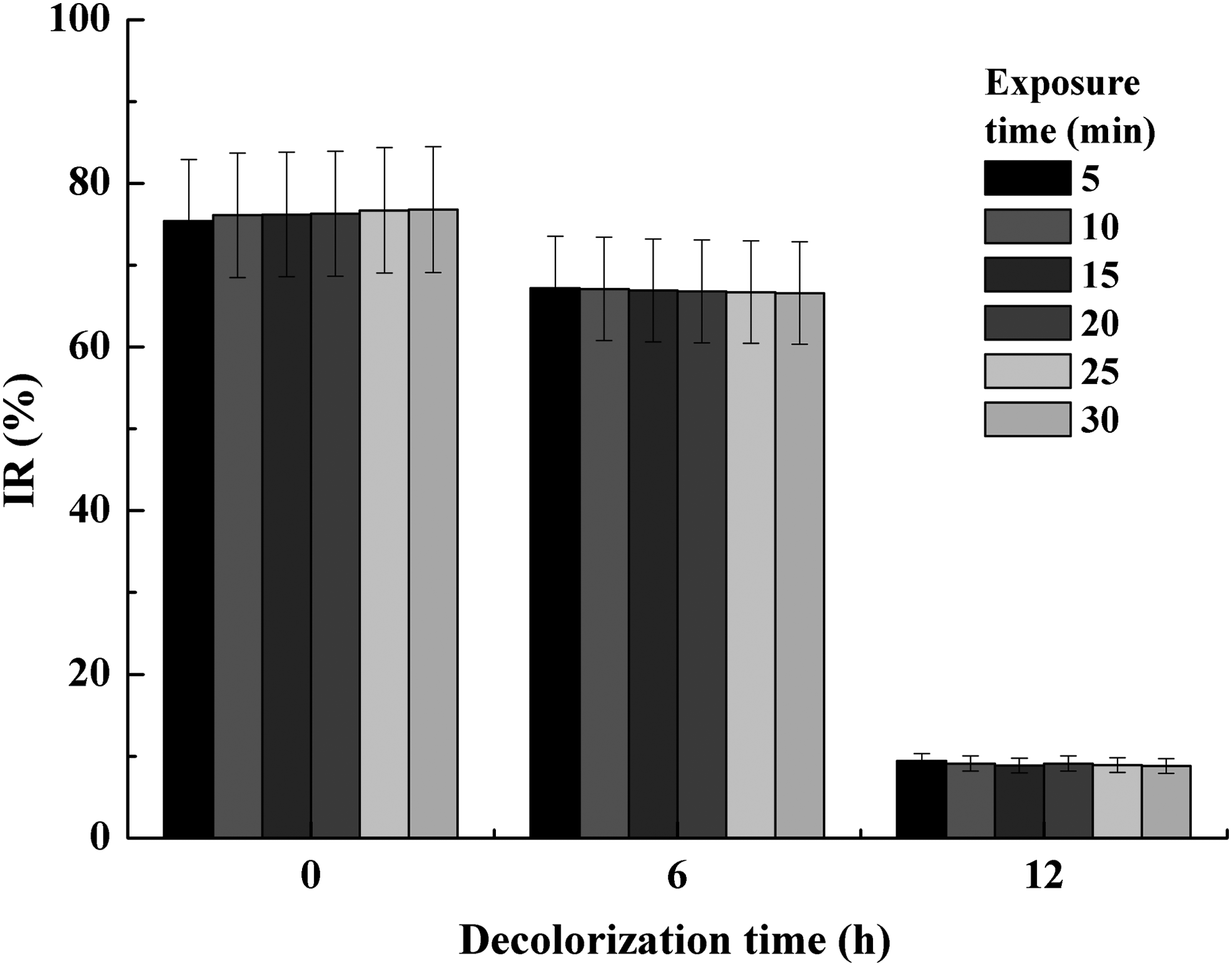
Acute toxicity of 50 mg/L ARB (with 30 g/L NaCl) before and after treatment by growing cells of yeast SYF-1 for 6 and 12 h, which were assessed by the Microtox test. 70 ≤ IR (%) < 100, high toxicity; 50 ≤ IR (%) < 70, moderate toxicity; 20 ≤ IR (%) < 50, low toxicity; 10 ≤ IR (%) < 20, micro toxicity; 0 ≤ IR (%) < 10, nontoxicity. IR, inhibition ratio.
It was reported that some biodecolorization intermediates of azo dye (e.g., aromatic amines) were more toxic than the dyes themselves and were difficult to be further degraded by plenty of bacteria (Qu et al., 2012). In comparison, fungi could degrade and detoxify many types of recalcitrant organic compounds depending on their nonspecific and nonstereoselective ligninolytic enzyme system (Kües, 2015). More and more fungi such as yeasts and white rot fungi had been explored and studied for treating azo dyes due to their detoxification effects (Hadibarata et al., 2013; Tan et al., 2016). It was also confirmed that the yeast SYF-1 could detoxify azo dye in this study.
The proposed decolorization pathway also displayed that some sulfonated and ammoniated byproducts with high acute toxicity were probably further degraded by the yeast SYF-1, resulting in a sharp decrease of acute toxicity. In addition, the yeast SYF-1 was a halotolerant type; thus, it might be potentially effective for the treatment of hypersaline industrial wastewaters in field application.
Conclusions
A halotolerant yeast SYF-1 that was capable of aerobically decolorizing azo dyes under hypersaline conditions was isolated, identified, and characterized in this study. The yeast SYF-1 was identified as C. tropicalis through 26S rDNA and ITS sequencing analysis and showed the highest ARB decolorization activity under the conditions as follows: 4.0 g/L glucose, 0.6 g/L ammonium sulfate, 0.08 g/L yeast extract, ≤30 g/L NaCl, rotation speed ≥160 rpm, temperature 30°C, and pH 7.0–8.0. The target azo dye ARB was possibly degraded by the yeast SYF-1 through a possible pathway consisting of reduction, monooxygenase-oxidization, reductive/oxidative deamination, oxidative desulfurization, ring-cleavage, and final TCA cycle, which might be different from that by most other microbes. Further, ARB was detoxified after being decolorized by the yeast SYF-1.
In conclusion, the halotolerant yeast SYF-1 may be potentially useful for the treatment of azo dye-contaminated hypersaline wastewaters.
Footnotes
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (Nos. 51878324, 51108223, and 51508259).
Author Disclosure Statement
Neither the entire article nor any part of its content has been published or has been accepted elsewhere. Meanwhile, the article is not under consideration by another journal at the same time as EES. All the authors have approved the submission to EES.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.