
Abstract
Toluene biodegradation from synthetic waste gas by aerobic denitrification using nitrate as an electron acceptor in a biotrickling filter (BTF) was investigated. Toluene removal efficiency achieved 92.3% in 105 days of operation. Pseudomonas, Paracoccus, Flavobacterium, SM1A02, Acinetobacter, Blastocatella, Moheibacter, Chlamydiales, Rhodobacter, and Ottowia were dominant toluene-degrading genus. Bradyrhizobium, Comamonas, Cupriavidus, Pseudomonas, Pseuoxanthomonas, and Ralstonia simultaneously had the ability to participate the process of toluene biodegradation and denitrification. BTF used nitrate or oxygen as electron acceptor, toluene as electron donor, and thereby redox between toluene and nitrate was formed, to realize toluene oxidation. BTF was capable of toluene oxidation by denitrification. These results show that the aerobic denitrifying BTF is achievable and open new possibilities for applying the BTF to the removal of volatile organic compounds.
Introduction
Volatile organic compounds (VOCs) are important precursors of ozone and secondary organic aerosol, which are emitted from gasoline evaporation, solvents and paint use, vehicular exhaust, and industrial processes (Wang and Qin, 2020). VOCs emitted to the environment highly probably result in ecological and health risks including cancer, neurotoxicity, and some other health impacts, whereas some hydrocarbons can lead to photochemical smog with other gases (NO x ) and produce ozone (Cheng et al., 2016a). Toluene is a symbolic gaseous contaminant of the typical aromatic VOCs, which could be emitted in petrochemical production, architectural coating, organic synthesis, packaging, and printing (Yao and Chen, 2020). In addition, it is capable of creating hazardous effects on environment and disadvantageous health problems (Win-Shwe and Fujimaki, 2010).
Biotechnology has proven to be a cost-effective method for the elimination of harmful VOCs and odor (El-Naas et al., 2014). The anoxic biodegradation of benzene, toluene, ethylbenzene, and xylene (BTEX), present as single compounds or in dual or quaternary mixtures by an acclimated bacterial consortium, was studied (Carvajal et al., 2018). The fungi-based biotrickling filter had high toluene elimination capacity (EC), the dominant fungi were Paramicrosporidium saccamoebae (Zhang et al., 2019). The compost/ceramic bead biofilter displayed good removal performance of gas-phase toluene and xylene mixture simultaneously (Rene et al., 2018). The biofilter packed with perlite showed better performance than the biofilter packed with tezontle in terms of the removal efficiency of toluene and bioaerosol emissions (Esquivel-Gonzalez et al., 2017). A gas-phase mixture of benzene and toluene was treated in a compost biofilter inoculated with mixed microbial consortium (Rene et al., 2015). Rhodococcus sp. ZJUT312 could finish efficient degradation of BTEX (You et al., 2018). The compost biofilter achieved waste gas treatment from high concentration of toluene and hexane (Zamir et al., 2012). A fungal–bacterial biofilter showed better removal efficiency than fungal or bacterial one alone in treating toluene, the combination of bacteria and fungi makes biofiltration a good option for VOC treatment (Cheng et al., 2016b). Biofilter aeration in the absence of pollutant supply prevented the washout of inhibitory metabolites originating from toluene biodegradation (Jiménez et al., 2017).
Co-metabolic enhancement of 1H-1,2,4-triazole biodegradation through nitrification could result in the enhanced removal of 1H-1,2,4-triazole; functional species related to nitrification and biodegradation were enriched with the supplement of NH4+, confirming the key role of nitrification (Wu et al., 2019). A low nitrate concentration resulted in a toluene flux decrease concomitant with biofilm growth by using nitrate as an electron acceptor (Emanuelsson and Livingston, 2002). Toluene mineralization by denitrification was successfully achieved for conversion of nitrate, toluene, and acetate into N2 and CO2 in an upflow anaerobic sludge blanket reactor (Martínez et al., 2007). Anoxic denitrifying processes could treat O2-free toluene emission in biotrickling filtration (Saucedo-Lucero et al., 2014). The denitrifying bacterium used, Thauera aromatica K172, could biodegrade toluene with nitrate respiration (Farhadian et al., 2010). Nitrate-reducing toluene-degrading microorganism, Azoarcus spp., could achieve degradation of toluene and ethylbenzene (Yang et al., 2011). But in aerobic conditions, the combination of toluene removal and denitrification was less investigated. In our preliminary study, there were independent nitrification, denitrification, and degradation of organic pollutants in the biological treatment of sludge-dried malodorous waste gas under aerobic conditions, and this suggests that aerobic denitrification and toluene degradation together in some bacteria might be inducible, aerobic denitrifying bacteria may be mutated into toluene-oxidizing bacteria after the supply of gaseous toluene to realize elimination of toluene.
The objective of this work is to study removal of toluene waste gas by an aerobic denitrifying biotrickling reactor (BTF) using nitrate as an external electron acceptor. The microbial community structures and gene function in BTF were comprehensively analyzed by 16S rDNA and metagenomics. The study aims to utilize Gas Chromatography-Mass Spectrometer (GC-MS) toward the understanding of the formation of the intermediate products and their involvement in the reaction mechanism. The mechanistic of toluene mineralization by denitrification was elicited, which is believed to provide impetus for the application of the aerobic denitrifying biotrickling reactor.
Materials and Methods
Setup and operation of the BTF system
The schematic of an aerobic denitrifying BTF for toluene removal using nitrate as an electron acceptor is shown in Fig. 1. The BTF was packed with ceramsites (Chen Xiang Co., Ltd.) to a height of 450 mm, it was divided into three stages with the filter medium at each section, and was supported on a stainless steel screen that ensured homogeneous distribution of gas flow over the entire cross-section of the filter bed, which was set up to study treatment-stimulated waste gas containing toluene. The main properties of lightweight ceramsites as filter media: external diameter of 5–10 mm, the density of 0.36–0.4 g/cm3, and a large specific surface area of 3,900 m2/m3. The ceramsites serve as a support for the aerobic denitrifying microbial population of toluene degradation and provides a large specific surface area for toluene and oxygen mass transfer. The BTF was made of transparent rigid polymethyl methacrylate with an inner diameter of 90 mm and a height of 1,000 mm. It was divided into three stages with the filter medium. The denitrifying bacterium was screened and tamed under aerobic condition from the activated sludge collected from the Liede wastewater treatment plant (Guangzhou, China). Denitrifying microorganisms adhered to the surface of the ceramsite to form the biofilm. Liquid medium of denitrifying bacterium consisted of sodium succinate (4.7 g/L), KNO3 (1 g/L), NH4Cl (0.3 g/L), KH2PO4 (1.5 g/L), Na2HPO4·7H2O (5 g/L), MgSO4·7H2O (0.1 g/L), and trace mineral (2 mL). The trace mineral solution contained EDTA (100 mg/L), ZnSO4 (4.4 mg/L), CaCl2 (11 mg/L), MnCl2·4H2O (10.2 mg/L), Fe2(SO4)3·7H2O (10 mg/L), (NH4)6Mo7O24·4H2O (2.2 mg/L), CuSO4·5H2O (3.2 mg/L), and CoCl2·6H2O (3.2 mg/L). The toluene (Guangzhou Chemical Reagent Factory Co., Ltd.) supplied from the gas cylinders, was first diluted with the compressed air, passed through an air mixture bottle, then flowed upward from the bottom of the biotrickling filter. Nutrient-containing aqueous solution was sprayed downward by a submerged pump (HX-2500; HAILEA Co., Ltd.) from the top of column to maintain the moisture of the BTF and supply nutrients to the microbial population. The liquid flow rate (Yu Yao Instruments Co., Ltd.) was used to control the flow at 2 L/min.
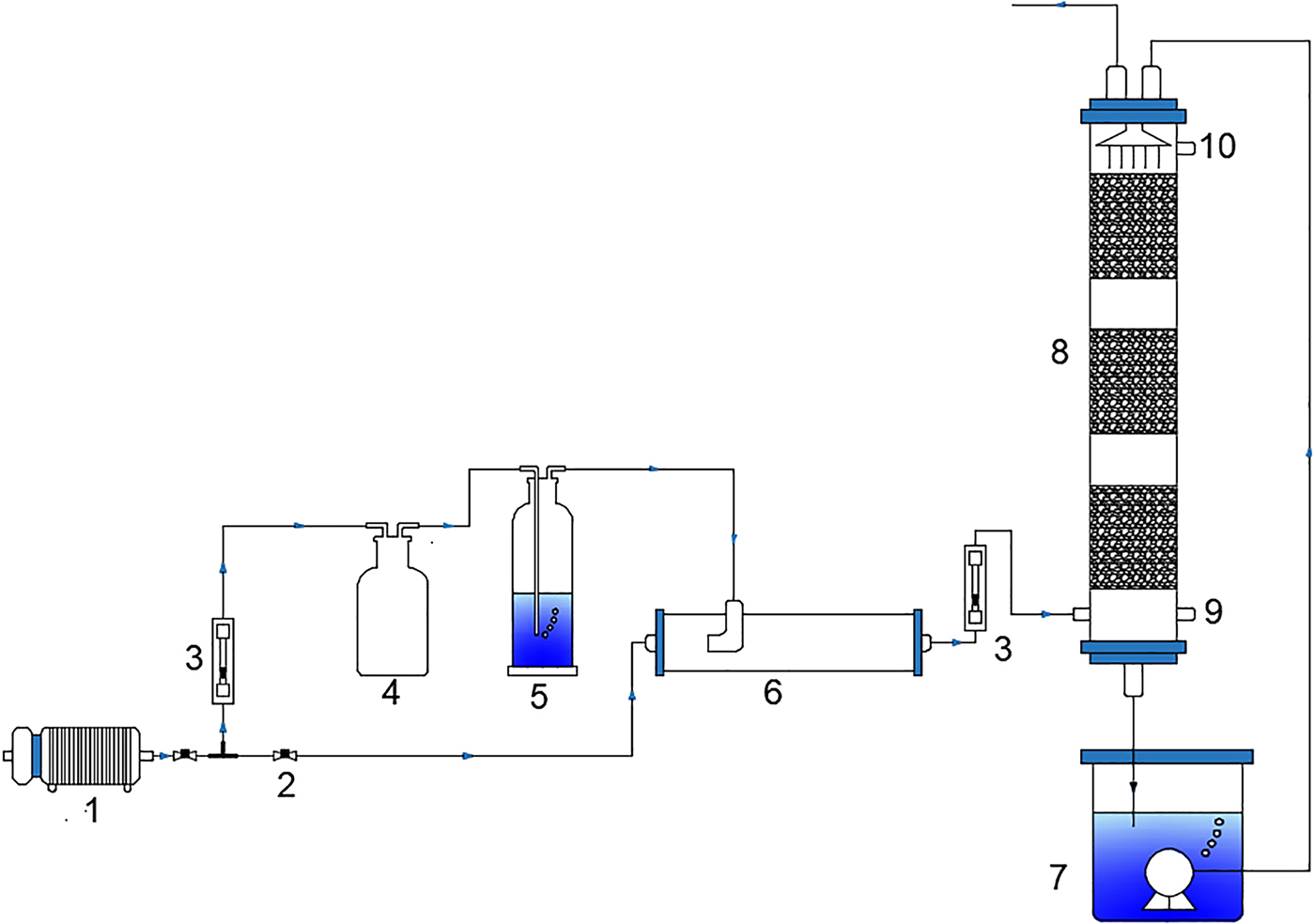
Schematic diagram of biotrickling filter for gaseous toluene removal with denitrification. (1) Air compressor, (2) Screw stop valve, (3) Flow meter, (4) Wide mouth bottle, (5) Monteggia washing bottle, (6) Mixing tube, (7) Nutrient tank, (8) Biotrickling filter, (9) Sample port of gas inlet, (10) Sample port of gas outlet.
The biotrickling filter was operated for toluene removal using nitrate as an electron acceptor and the only nitrogen source at oxygen concentration of 21% during the 105-day continuous running test. Eight phases were observed: start-up phase under the condition of toluene inlet concentration of 100.6–116.9 mg/m3, empty bed residence time (EBRT) of 34.3 s, and sprinkling amount of 2 L/min. From phase I to phase XI, we researched the influence inlet concentration, and EBRT on the BTF's performance was investigated in the phase II, III, IV, and V, under the conditions of inlet concentration of 151.3–170.1, 201.5–218.3, 250.8–266.4, 302.9–316.2, and 596.3–618.2 mg/m3, EBRT of 34.3 s, respectively. In the phase VII and VIII, under the conditions of the inlet concentration of 300.2–310.2 mg/m3, EBRT of 22.8 and 19.6 s, respectively. The performance of the BTF was evaluated in terms of toluene removal efficiency (RE, %) and the EC of toluene (EC, g/[m3·h]). To continuously create aerobic denitrifying condition for toluene biodegradation, we supplied 1.2–1.5 g KNO3 to the nutrient tank every 10 days, meanwhile we also supplied 1.5 g KH2PO4, 5 g Na2HPO4·7H2O, 0.1 g MgSO4·7H2O, 2 mL of trace mineral. The concentration of nitrate, nitrite, and ammonium were monitored, respectively, by UV spectrophotometry.
16s rDNA sequencing and metagenomic sequencing
The biofilms in BTF were sampled on the 60th day, bacterial community compositions were assessed by 16s rRNA and Metagenomics sequencing. DNA was extracted from the sample by using the PowerSoil DNA Isolation Kit. Then the extracted DNA was amplified by the polymerase chain reaction. The amplicons were sequenced by Illumina Hiseq 2500 or Miseq platform, which used the paired-end sequencing method. Each sequence produced 250 (Hiseq 2500) or 300 bp (Miseq) reads from the 5′ and 3′ terminals. A series of software (Mothur, Flash, Trimmomatic QIIME, etc.) were utilized to filtrate the reads of low quality and got clean data, to assemble short pair-end reads, clean contigs, OTU picking, taxonomic annotation, ad analysis of alpha diversity.
The sample for Metagenomic sequencing (Magien Biotechnologies Co., Ltd.) was collected on the 60th day, DNA was extracted, amplified, and purified. A library of DNA fragments (300 bp) was constructed according to standard protocol of Illumina and PE150 sequenced by Illumina HiSeq 2500 platform. The raw data were converted to raw reads after base calling and stored in FASTQ file format. Then replicated reads, short reads, and reads containing excessive N-bases were removed from the dataset. This analytic method in this study was mainly for functional annotation, which was mapped against KEGG database (Kyoto Encyclopedia of Genes and Genomes).
Analytical methods
The toluene concentration was measured by a gas chromatography (JieDao GC1690) with an SE 30 column and a flame ionization detector. The temperature of the oven, sample injector, detector was set at 140°C, 200°C, 200°C, respectively. Flow rate of gas and liquid was measured by Model LZB-1 flow meters (Yuyao instruments Co., Ltd., Zhejiang, China). pH of the recirculating liquid was determined by Model pHB-3pH Tester (Shanghai Sanxin Instrument, Inc., Shanghai, China). Dissolved oxygen (DO) of the recirculating liquid was determined by DO analyzer (YSI550A; Yellow Springs Instrument, Inc., USA). Concentrations of NO3−-N, NO2−-N, and NH4-N in the recirculating liquid were determined by UV spectrophotometer (T6; Beijing Persee Instruments Co., Ltd., China). Concentrations of carbon dioxide in inlet and outlet were determined by carbon dioxide analyzer (testo 350; Testo AG Co., Ltd., Germany). The microbial thickness and biofilm oxygen vertical distribution were measured by the dissolved oxygen microelectrode (Unisense A/S Co., Ltd., Denmark). The intermediate biodegradation products in the liquid were concentrated by solid phase extraction (SPE) cartridge and then washed into methanol, which were ultimately determined by Triple Quadrupole Gas Chromatography/Mass Spectrometer (TSQ Quantum XLS, Thermo Co., Ltd.). The initial temperature was 40.0°C. Source temperature was 230°C. Initial hold time was 3.00 min. Full scan was from 29.000 to 550.000. There was one ramp. The rate was 10.0°C/min. Final temperature was 240. Electro-ion (EI) was set at 70 eV. Total hold time was 10.00 min. S/SL mode was splitless and temperature was 240°C. Carrier flow was 1.000 mL/min.
Results
Toluene oxidation coupled to denitrification
Figure 2 showed removal performance of toluene in waste gas by aerobic denitrification using nitrate as an external electron acceptor in a biotrickling filter during eight-phase (105 days) operation. During the start-up phase (day 1–25), toluene removal efficiency changed from 30.1% on the first day, fluctuated slightly between 30.1% and 35.6% from day 2 to 14, increased from 45% on the 15th day to 92.3% on the 19th day, and then fluctuated between 89.9% and 94.8% from day 20 to 25. EC was only 3.22 g/[m3·h] on the first day, increased from 5.16 g/[m3·h] on the 15th day to 10.99 g/[m3·h] on the 19th day. This indicated that the microbes had basically adapted to the toluene as the sole carbon source, nitrate as an external electron acceptor, and the biofilm acclimatization, stabilized, and the biodegradation should be responsible for the gaseous toluene removal.

Toluene removal efficiency
From phase I to phase VI (day 26–85), the influence of toluene inlet concentration on the BTF's performance was investigated. In phase I, the toluene inlet concentration was stabilized at 101.2–118.8 mg/m3. The RE ranged within 91.7–97.4% EC of 10.22–11.91 g/[m3·h]. In phase II, the toluene inlet concentration was increased to 151.3–170.1 mg/m3, RE sharply dropped at 87.1% (EC of 14.71 g/[m3·h]) on the 36th day, but it recovered to 95.4% (EC of 15.13 g/[m3·h]) on the 37th day and then ranged within 90.2–95.4% (EC of 14.67–17.02 g/[m3·h]). In phase III, the toluene inlet concentration was further increased to 201.5–218.3 mg/m3, RE was declined at 88.0% (EC of 19.21 g/[m3·h) on the 46th day and it gradually recovered to 95.0% (EC of 20.07 g/[m3·h]) on the 52nd day, but then RE dropped to 91.0% (EC of 19.34 g/[m3·h]) on the 55th day. In phase IV, the toluene inlet concentration rose to 250.8–266.4 mg/m3, RE was declined to 85.6% (EC of 22.86 g/[m3·h]) on the 56th day, and it did not show an uptrend, which ranged within 83.0–85.6% (EC of 21.98–23.02 g/[m3·h]). In phase V, the toluene inlet concentration was increased to 302.9–316.2 mg/m3, RE dropped at 79.9% (EC of 26.04 g/[m3·h]) on the 66th day and ranged within 79.2–82.3% (EC of 25.30–26.81 g/[m3·h]). In phase VI, the toluene inlet concentration was drastically increased to 596.3–618.2 mg/m3, RE ranged within 77.7–78.8% (EC of 48.65–50.33 g/[m3·h]). This demonstrated that the biofilm showed relatively high biological activity, BTF can withstand the impact of different loads and maintain high efficiency.
In phase VII and VIII (day 86–105), the influence of EBRT on BTF's performance was investigated, shortening the EBRT to 22.8 s in phase VII and 19.6 s in phase VIII. The toluene inlet concentration was maintained at 300.2–310.2 mg/m3. In phase VII, RE sharply dropped to 53.0% (EC of 16.94 g/[m3·h]) on the 86th day and then was stabilized within 53.0–54.0% (EC of 16.75–17.51 /[m3·h]). This downtrend was mainly due to the short EBRT, which could decrease the mass transfer for the hydrophobic toluene from air to liquid. In phase VIII, RE dropped to 39.85% (EC of 12.74 g/[m3·h]) on the 96th day and it recovered to 46.4% (EC of 14.68 g/[m3·h]) on the 101st day. Then it ranged within 46.9–49.8% (EC of 14.67 to 16.09 g/[m3·h]) from days 102 to 105, which was still lower than that in phase VII. The slight recovery was likely to be attributed to the growth of biofilm in such an operating condition.
Figure 2c showed the cumulative toluene and nitrate consumption during the 30 days (31st to 60th day). We could see that the amount of toluene removed by the BTF was continuously increasing from zero in the beginning to 0.31 mol on the 30th day. The nitrate consumption increased instantly to 0.02, 0.04, and 0.06 mol on the 5th, 14th, and 24th day respectively, after nitrate supplement and then quickly kept stable. Meanwhile, the concentration of nitrate in the circulating liquid was close to zero. In Fig. 2d, we determined the oxygen environment of the biofilm in BTF. It was clearly seen that the thickness of the biofilm was about 540 μm. The aerobic zone (DO >2.00 mg/L) was about 300 μm in thickness and the anoxic zone (0.50 mg/L > DO >0.20 mg/L) was about 40 μm. Moreover, the anaerobic zone (DO <0.20 mg/L) was hardly being detected. The aerobic environment occupied most of the biofilm. Based on previous research, when nitrate was used as sole nitrogen source under aerobic condition, the aerobic denitrification should be responsible for the nitrogen removal (Lin et al., 2020). Moreover, the nitrate and oxygen could be simultaneously used as electron acceptor in aerobic denitrifying process (Robertson et al., 1988; Huang and Tseng, 2001). This could explain the growing disparity between toluene and nitrate removal in Fig. 2c. Probably it was because the nitrate supplement was insufficient, the BTF system could utilize much more nitrate as electron acceptor. Meanwhile, the oxygen could participate in the toluene oxidation.
The toluene removal performance and possibility under anaerobic condition with nitrate as electron acceptor was deliberately investigated in many kinds of reactors, in which the efficiency of toluene removal and denitrification in the liquid phase could exceed 95% (Evans et al., 1992; PeA-Calva et al., 2004; Martínez et al., 2007). In addition, it was reported that the toluene degradation rate of PKO1 under hypoxic conditions was greatly enhanced in the presence of nitrate while the remaining was unaffected under aerobic conditions (Leahy and Olsen, 1997). It was believed that oxygen could be used first for the initial oxygenase attack on aromatic hydrocarbons in aerobic denitrification process and then yield oxidized intermediates more susceptible to anaerobic degradation (Kim and Jaffé, 2008). In our research, the anaerobic zone in the biofilm was hardly being detected, so we think that most nitrates were reduced at a much higher oxygen concentration. The nitrate could get efficiently utilized for toluene biodegradation in a completely aerobic environment.
These results confirmed that the biotrickling filter could achieve toluene oxidation using nitrate as an electron acceptor, and the aerobic denitrification was confirmed to be coupled to toluene biodegradation with a high efficiency (Saucedo-Lucero et al., 2014); the oxygen and nitrate could play the role of electron acceptor at the same time. When nitrate is lacking, oxygen could make up for the gap. Toluene-degrading community has been formed and acclimatized, and denitrifying bacteria subjected to long-term exposure of toluene could reduce nitrate to nitrogen through denitrification.
Toluene degrading and denitrifying microbial community
Bacterial community compositions were assessed by 16s rRNA. The filtered sequences were 26,477 reads, which were taxonomically classified into 208 OTUs. The Chao 1 index for estimating OTUs was 259.908 as well as the Shannon index of 4.571.
The bacterial community compositions at both phylum and genus levels, as well as the relative abundance are shown in the Fig. 3, at phylum level, the relative abundances of dominant bacteria Proteobacteria, Bacteroidetes, Patescibacteria, Planctomycetes, Dependentiae, Armatimonadetes, and Acidobacteria were 54.7%, 14.2%, 7.0%, 5.3%, 4.4%, 4.1%, and 3.3%, respectively. Proteobacteria was an aerobic denitrifying bacteria (Wang and Chu, 2016). Proteobacteria and Bacterioidetes were dominant in the enriched consortia, which were cultured by toluene (Su et al., 2014). Proteobacteria was the predominant phylum for anoxic biodegradation of BTEX (Akmirza et al., 2017).
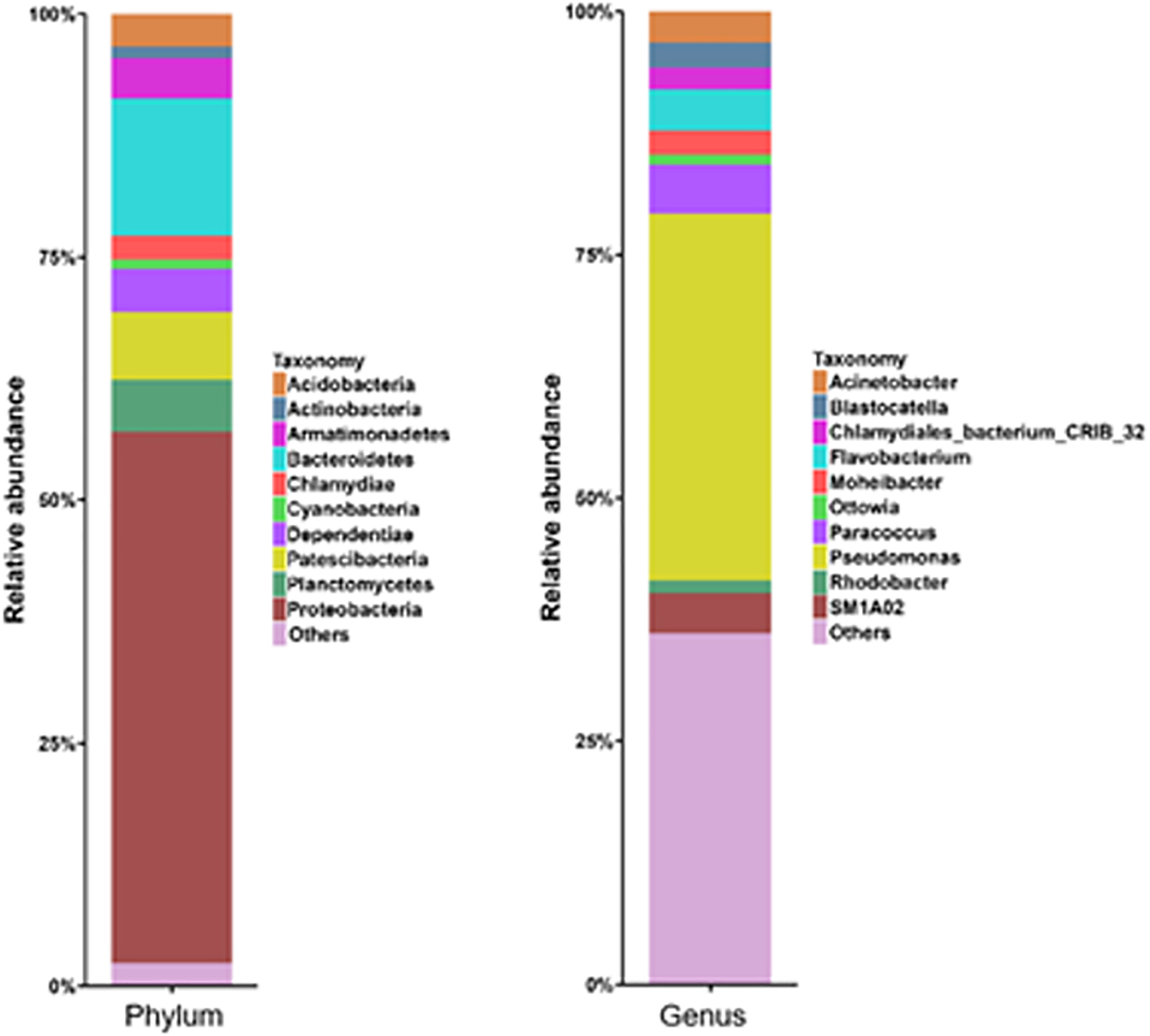
Toluene degrading microbial community at phylum and genus levels.
As shown in Fig. 3, at the genus level, the relative abundances of ten dominant genera Pseudomonas, Paracoccus, Flavobacterium, Acinetobacter, Blastocatella, and Moheibacter were 37.7%, 5%, 4.2%, 3.2%, 2.5%, and 2.4%, respectively. Pseudomonas could utilize the benzene, toluene, and xylene as carbon resources, and degrade the toluene (Martino et al., 2012). The genera Paracoccus and Flavobacterium had already been proven to be denitrifiers (Carvajal et al., 2018).
Functional genes, toluene degrading, and denitrifying microorganism
As shown in Fig. 4, functional genes for toluene biodegradation (tmoABCDEF, tbuA1UBVA2C, todC1C2BADEF) and denitrification (napA, napB, nirK, nirS, norB, norC, nosZ) were found out to have relevant taxonomic origins and different contributions from specific genera. The relative abundance of denitrification (napA, napB, nirK, nirS, norB, norC, nosZ)-related genes was the highest (0.0549%), followed by toluene monooxygenase (tmoA/tbuA1/touA, tmoB/tbuU/touB, tmoC/tbuB/touC, tmoD/tbuV/touD, tmoE/tbuA2/touE, tmoF/tbuC/touF) (0.0284%), subunit xylene monooxygenase and benzaldehyde dehydrogenase (xylC, xylX, xylY, xylZ, xylL) (0.0124%), toluene dioxygenase (TDO) (todC1, todC2, todB, todA, todD, todE, todF) (0.0039%), phenol hydroxylase gene (dmpK, dmpL, dmpM, dmpN, dmpO, dmpP, dmpB, dmpC, dmpD, dmpH) (0.082%), catechol 2,3- dioxygenase Cat (catA, catB, catC) (0.008%). The relative abundances of other toluene degrade functional genes mhp, bph, pca, praC, box, and pobA were 0.0196%, 0.0191%, 0.0539%, 0.0111%, and 0.0093%, respectively. Among these functional genes, tmoABCDEF, tbuA1UBVA2C, touABCDEF, and dmpKLMNOP were responsible for the synthesis of toluene 4-monooxygenase (T4MO), toluene 3-monooxygenase (T3MO), toluene/o-xylene monooxygenase (ToMO), and phenol/toluene 2-monooxygenase (T2MO), respectively. xylC, E 1.1.1.90, benABC-xylXYZ, bend-xylL, catA, catB, catC were responsible for the synthesis of benzaldehyde dehydrogenase, aryl-alcohol dehydrogenase, benzoate/toluate 1,2-dioxygenase, dihydroxycyclohexadiene carboxylate dehydrogenase, catechol 1,2-dioxygenase, muconate cycloisomerase, muconolactone D-isomerase, respectively (Byrne et al., 1995; Bertoni et al., 1998).
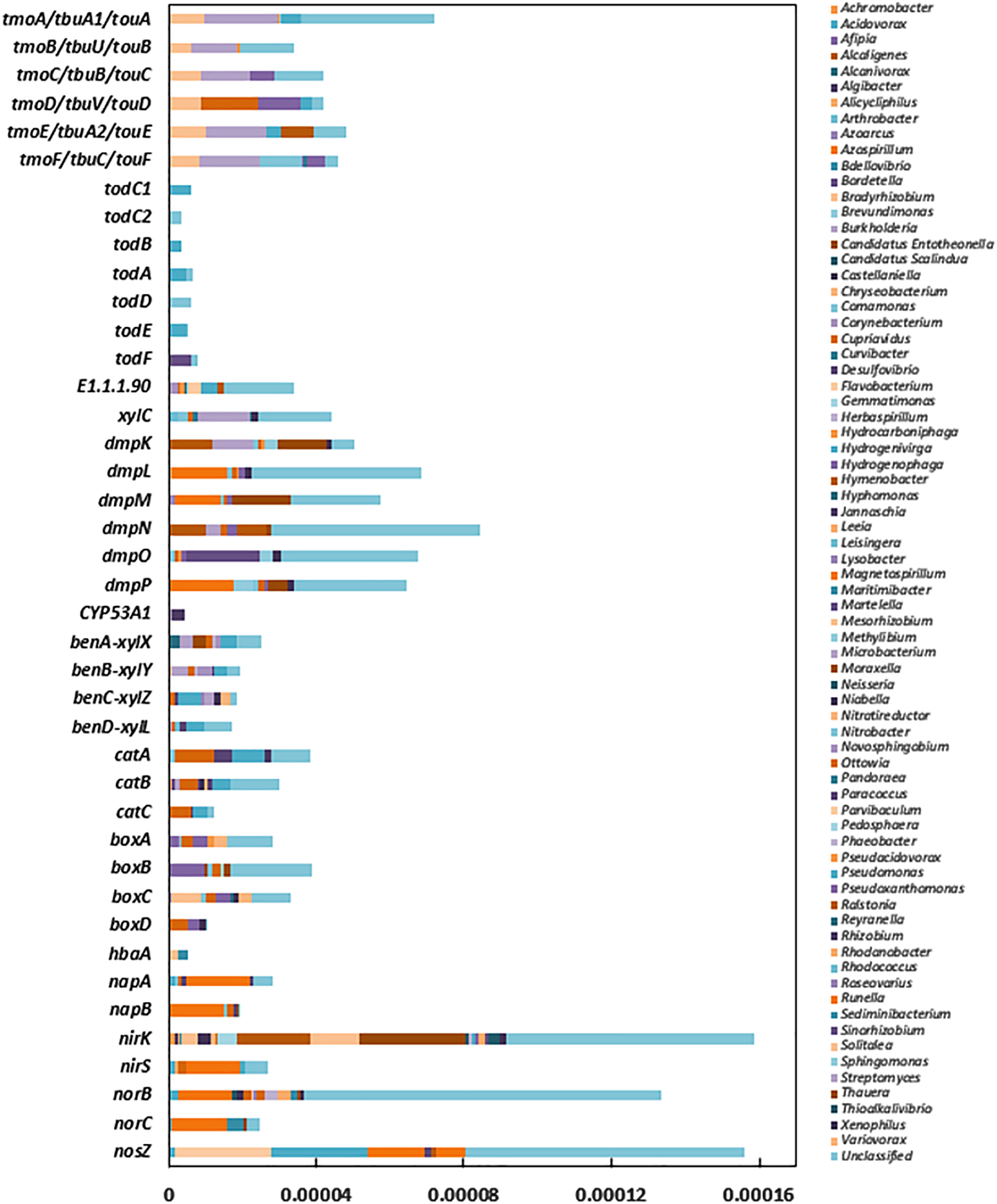
Segmental functional genes of toluene biodegradation and denitrification.
In this BTF system, Burkholderia was annotated with tmo, tbu, tou, dmp, xyl, and catB. Bradyrhizobium was annotated with tmo, tbu, tou, dmpB, and xylY. Hydrocarboniphaga was annotated with tmo, tbu, tou, and dmp. Pseudomonas was annotated with tmo, tbu, tou, todC1C2BAE, dmp, xyl, catA, catB, and catC. Rhodococcus was annotated with tmoA/tbuA1/touA, and todE. Cupriavidus was annotated with tmoD/tbuV/touD, xyl, dmp, catA, catB, andcatC. Ralsotonia was annotated with tmoE/tbuA2/touE, and dmpN. Comamonas was annotated with tmoF/tbuC/touF, xylC, dmpK, dmp, and catA. Pseudoxanthomonas was annotated with tmoF/tbuC/touF, and dmpP. Pandoraea was annotated with tmoF/tbuC/touF, xylC, and dmpC. Hydrogenophaga was annotated with tmo, tbu, tou, and dmp. Azospirillum was annotated with dmpL, dmpM, dmpP, and dmpH. Thauera was annotated with dmpK, dmpM, dmpN, dmpP, and dmpB. Theses genera were responsible for toluene metabolism. Pseudomonas could utilize the TDO, which was encoded by the cluster genes todC1C2BA, to catalyze the initial cis-dihydroxylation. The aerobic denitrifiers were dominantly represented by annotated genera, Acidovorax (napA, nirS, norB, norC, nosZ), Azospirillum (napB, norB, norC), Bradyrhizobium (nirK), Comamonas (napA, napB, norC), Cupriavidus (napA, napB, nirK, nirS, norB), Hydrogenivirga (nosZ), Magnetospirillum (napA, nirS, nosZ), Paracoccus (napA, napB, nosZ), Pseudomonas (nirK, nirS), Pseudoxanthomonas (nirK), Ralstonia (nosZ), and Thauera (napB, norB, norC).
As is shown in Fig. 5, Achromobacter, Acidovorax, Alicycliphilus, Azospirillum, Bradyrhizobium, Castellaniella, Comamonas, Cupriavidus, Desulfovibrio, Gemmatimonas, Mesorhizobium, Ottowia, Paracoccus, Pseudomonas, Pseudoxanthomonas, Ralstonia, Sinorhizobium, Thauera, and Xenophilus possessed related functional genes of both toluene degradation and aerobic denitrification, which were feasible to participate the toluene degradation driven by denitrifying. Pseudomonas (37.7%) was the dominant bacteria, which had the greatest potential to achieve denitrifying/toluene oxidizing. Thauera degrades toluene only under denitrifying conditions, Ralstonia can perform nitrate-enhanced toluene biodegradation using oxygen and nitrate as electron acceptor (Kim and Jaffé, 2008).

Different bacterial genera for toluene biodegradation and denitrification.
Discussion
Electron transfer analysis of toluene biodegradation
The biotrickling filter could achieve effective removal of toluene in 105 days of operation, toluene oxidation was driven by aerobic denitrification using nitrate as an external electron acceptor. From an electrochemical standpoint, toluene serves as electron donor in bacterial oxidation, nitrate played a role as electron acceptor in denitrification, oxygen as electron acceptor would participate in the toluene oxidation. When toluene was oxidized, the released electrons must be ultimately transferred to nitrate or O2. As electron acceptor in denitrification, nitrate can capture electrons released from toluene and be gradually reduced to nitrogen. O2 can capture electrons released from toluene in enzymatic oxidation. The stoichiometric balanced equation should be added here based on the assumption of the complete C oxidation (Heider, 2007).
Toluene mineralization rate fluctuated slightly within 50–62% during the start-up phase (1–15-day) of BTF, and then gradually increased, stable about 80% from days 36 to 105. Kuhn et al. used a laboratory aquifer column and observed about 82% of the initial toluene-C transforming into carbon dioxide (Kuhn and Zeyer, 1988). Martinez et al. studied the toluene mineralization by denitrification in an upflow anaerobic sludge blanket reactor and obtained an average rate of toluene mineralization over 70% together with acetate (Martínez et al., 2007). Aerobic denitrifying bacteria used toluene as electron donor and the sole carbon source, nitrate as an external electron acceptor under aerobic condition, and oxidized toluene to CO2; the oxygen may participate in the toluene oxidation together with nitrate, and nitrate was reduced to organic nitride by assimilation denitrification, and nitrate was transformed into N2 by dissimilation denitrification (Chiang and Salanitro, 2005). Toluene oxidation could be efficiently coupled to aerobic denitrification. The stoichiometric balanced equation in BTF based on toluene mineralization rate of 80% can be expressed as:
Mechanism of toluene biodegradation driven by aerobic denitrification
Triple quadrupole gas chromatography/mass spectrometer implied that there were benzyl alcohol and benzaldehyde (Fig. 6) in the circulating liquid. Mechanism of toluene biodegradation driven by aerobic denitrification is shown in Fig. 7. In the aerobic denitrifying BTF system, toluene was oxidized to carbon dioxide and water through aerobic denitrification (napAB, nirKS, norBC, nosZ) using nitrate or oxygen as electron acceptor and toluene as electron donor. Toluene was initially catalyzed by toluene 4-monooxygenase (tmoABCDEF) to produce benzyl alcohol, then into benzaldehyde under the catalyzation of aryl-alcohol dehydrogenase (E 1.1.1.90) (Kasai et al., 2001). The 4-monooxygenase possesses similar structures (ferredoxin and NADH: ferredoxin oxidoreductase component) with the xylene monooxygenase, which can catalyze the introduction of a hydroxyl group on a methyl side-chain carbon of toluene (Yen et al., 1992). The benzoate would be next formed by the benzaldehyde dehydrogenase (xylC) (Inoue et al., 1995). Benzoate would be catalyzed by benzoate/toluate 1,2-dioxygenase (benABC-xylXYZ) and dihydroxy-cyclohexadiene carboxylate dehydrogenase (benD-xylL) to produce catechol. Benzoate and catechol were not detected in the liquid, because the consumption of the two substances were relatively complete and the accumulated amount was pretty small. The catechol would be catalyzed into succinyl-CoA and acetyl-CoA and finally entered the citrate cycle to produce carbon dioxide and water, which was successively catalyzed by a series of enzymes (cat, pca, dmp, mhp, bph, pra, etc.).

Detection of the intermediates of toluene degradation by GC-MS. GC-MS, Gas Chromatography-Mass Spectrometer.

Mechanism of toluene biodegradation for aerobic denitrification.
The mechanism we proposed was different from the previous research. Shinoda et al. isolated the new Thauera sp. Strain DNT-1 and studied its performance of toluene degradation under aerobic and anaerobic denitrifying condition (Shinoda et al., 2004). The toluene was initially catalyzed by the dioxygenase (todC1C2BA) into cis-toluene dihydrodiol and then transformed into 3-methylcatechol by toluene dihydrodiol dehydrogenase (todD), which was different from the monooxygenase (tmoABCDEF) pathway. Although we have annotated the same functional genes, we had not detected the key intermediate product 3-methylcatechol. This is probably because the tod gene cluster did not get fully expressed in the BTF system. We also did not find the strain DNT-1, and the genes bssDCABE did not get annotated in our research. This gene cluster usually got induced under anaerobic condition. The metabolic pathway of toluene is the formation of catechol through toluene monooxygenase or dioxygenase, and then through ortho- or interposition cutting ring; toluene degradation is catalyzed by the enzyme benzylsuccinate synthase using nitrate as electron acceptor under anoxic conditions (Vogt et al., 2008).
Conclusions
The present work proves that the aerobic denitrifying drove the gaseous toluene biodegradation in the biotrickling filter. Toluene removal efficiency achieved 92.33% and the removal capacity could achieve 10.99 g/[m3·h]. The dominant genera, Pseudomonas, Paracoccus, Flavobacterium, Acinetobacter, Blastocatella, Moheibacter, Chalamydiales, Rhodobacter, Ottowia at 60. Achromobacter, Acidovorax, Alicycliphilus, Azospirillum, Bradyrhizobium, Castellaniella, Comamonas, Cupriavidus, Desulfovibrio, Gemmatimonas, Mesorhizobium, Ottowia, Paracoccus, Pseudomonas, Pseudoxanthomonas, Ralstonia, Sinorhizobium, Thauera, and Xenophilus, were proposed to simultaneously participate in toluene biodegradation and aerobic denitrification. The mechanism for toluene oxidation by denitrification can be described, as the BTF used nitrate or oxygen as electron acceptor, toluene as electron donor, and thereby redox between toluene and nitrate was formed, to realize toluene oxidation. This provides a new way of VOC removal using an aerobic denitrifying BTF.
Footnotes
Acknowledgment
The authors gratefully acknowledge the financial support from the Nation Nature Scientific Research Foundation of China.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Nation Nature Scientific Research Foundation of China (21677178).