
Abstract
In the case of a suspected anthrax attack, the United States Food and Drug Administration has a plan to immediately distribute stockpiled antibiotics to the affected population. If performed on a large scale, the amount of antibiotics entering wastewater treatment plants would quickly increase. This influx of drugs would likely be followed by unknown consequences to wastewater treatment plants, downstream ecology, and public health. We therefore investigated the influence of the antibiotic ciprofloxacin on bench scale activated sludge reactors. Reactor performance was unaffected regarding removal of total organics during 14 days of antibiotic loading; however, the reactor microbial community was affected immediately following ciprofloxacin-challenge. Aerobic colony-forming units were initially reduced at 24 h and then recovered after 72 h. Analysis of 16S rRNA gene sequences demonstrated an increased relative abundance of Burkholderiales, Bradyrhizobiaceae, Microbacteriaceae, and Emticicia spp. In addition, ciprofloxacin decreased the Shannon's diversity index and community richness. Further, analysis of randomly selected bacterial isolates demonstrated that the prevalence of multidrug resistance increased in the ciprofloxacin-challenged reactors. These results suggest that an emergency response-associated loading of ciprofloxacin into municipal wastewater treatment plants could alter the activated sludge microbiome and select for multidrug-resistant bacteria.
Introduction
Bacillus anthracis is classified as a Category A agent by the Centers for Disease Control and Prevention (CDC) due to the high mortality rates of inhaled anthrax and the potential for this organism to be weaponized (Jernigan et al., 2002; CDC, 2018). In the event of a suspected anthrax attack, the United States Food and Drug Administration (US FDA) has issued an order permitting emergency distribution of the antibiotics ciprofloxacin and/or doxycycline to populations suspected to be exposed to B. anthracis spores (Narayanan et al., 2018; US FDA, 2018). Such widespread antibiotic consumption and excretion has the potential to disrupt wastewater treatment by altering the activated sludge microbiome and selecting for antibiotic-resistant bacteria. In addition, the presence of antibiotic resistance genes could increase in frequency throughout the wastewater treatment infrastructure and downstream environments. Such emergency action plans may result in unplanned consequences for wastewater treatment operation and public health.
Since the introduction of antibiotics in human medicine and agriculture, bacterial pathogens capable of causing life-threatening infections have emerged that harbor one or many antibiotic resistance genes and/or mutations. It is predicted that by 2050, infections caused by antibiotic-resistant pathogenic bacteria will lead to 10 million deaths per year and a total GDP loss of $100 trillion (O'Neill, 2014). Today, serious infections caused by pathogens resistant to multiple antibiotic classes have become common. To illustrate how widespread antimicrobial resistance has become, a multiple antimicrobial-resistant strain of bacteria was isolated from the International Space Station (Singh et al., 2018).
One group of bacterial pathogens, described by the term “ESKAPE” (Enterococcus faecium, Staphylococcus aureus, Klebseilla pneumoniae, Acinetobactor baumannii, Pseudomonas aeruginosa, and Enterobacter spp.), are particularly capable of expressing multiple antimicrobial resistance (Pendleton et al., 2013). Besides the ESKAPE group, there are many other bacterial pathogens, including Citrobacter freundii that can express multiple antimicrobial resistance. This gram-negative organism is found in the gastrointestinal tract of humans and animals as well as environmental sources, and causes a wide array of diseases often in patients that are immunocompromised or suffering from underlying disease (Toranzo et al., 1994; Porres-Osante et al., 2015; Liu et al., 2018a).
Antibiotics enter municipal wastewater treatment facilities through hospital and pharmaceutical wastewater discharge, human excretion of unmetabolized antibiotic, and from improper household disposal. Concentrations of antibiotics in raw sewage are commonly reported to be in the ppb range (Karthikeyan and Meyer, 2006; Miège et al., 2009). During municipal wastewater treatment, many antibiotics will partition between the aqueous phase and wastewater solids; however, their overall mass usually persists (Golet et al., 2003; Lindberg et al., 2006; Heidler and Halden, 2008; Ferreira et al., 2016). Thus, many antibiotics enter the environment through wastewater effluent and biosolids (Zhang and Li, 2011), contributing to the selection of antibiotic-resistant bacteria within wastewater treatment plants (Hultman et al., 2018; Manaia et al., 2018).
Ciprofloxacin is easily detected in sewage due to its widespread use and low metabolism in patients (Rohwedder et al., 1990; Daughton and Ternes, 1999; Heberer, 2002; Karthikeyan and Meyer, 2006; He and Blaney, 2015; Johnson et al., 2015). Ciprofloxacin is a member of the fluoroquinolone class of antibiotics (Chin and Neu, 1984) that is commonly prescribed for the treatment of a wide variety of bacterial infections (Almalki et al., 2017). The wide spread clinical use of ciprofloxacin has led to the selection of bacterial pathogens that demonstrate resistance to this broad-spectrum antibiotic (Hooper and Jacoby, 2015; Vinué et al., 2016). One municipal wastewater treatment facility demonstrated ciprofloxacin concentrations ranging from 0.6 to 1.292 μg/L in raw wastewater, 0.213 to 0.504 μg/L in treated wastewater, and 0.0122 to 0.102 μg/L in surface water downstream of the wastewater treatment plant over a 4 month period (He and Blaney, 2015). Another study of four plants found ciprofloxacin concentrations ranging from 3.28 to 69.5 μg/L in the influent and 2.72 to 16.4 μg/L in the effluent (Kulkarni et al., 2017).
Some of the effects of ciprofloxacin loading on wastewater treatment plants has been evaluated at the laboratory scale. Concentrations of 0.2 and 2 mg/L, ciprofloxacin were found to reduce biological nitrogen and phosphorus removal processes in bench scale reactors (Yi et al., 2017). Ciprofloxacin was found to affect bench scale partial-nitritation wastewater bioreactors amended with 100 to 350 ng/L of ciprofloxacin by reducing nitritation and ammonium oxidizing bacteria while increasing Commamonas sp. (Gonzalez-Martinez et al., 2014). In a wetland experiment, mesocosms seeded with wastewater treatment plant-activated sludge were found to have reduced diversity of bacteria species and a temporary reduction in catabolic activities when dosed with 2 mg/L of ciprofloxacin (Weber et al., 2011). The addition of over 0.5 mg/L of ciprofloxacin in anerobic sludge microcosms was found to decrease Syntrophobacter and Methanothrix and affect reactor performance (Mai et al., 2018). Thus, a sudden and large loading of ciprofloxacin due to emergency antibiotic distribution after an anthrax attack poses a threat to wastewater treatment and downstream systems.
In the event of a suspected anthrax attack or other health emergency, the rapid and widespread release of antibiotics to the general population could quickly increase the amount of antibiotics entering wastewater treatment plants. In the present study, a relatively large dose of ciprofloxacin was loaded into laboratory scale activated sludge reactors to determine the effects of this antibiotic on reactor performance, the wastewater microbial community, and on the selection of antibiotic-resistant bacteria.
Methods
Development and operation of bench-scale activated sludge bioreactors
Combined activated sludge/sedimentation reactors were built as previously described (Stover and Kincannon, 1983; Barber and Veenstra, 1986). The reactors included a 3 L activated sludge compartment, which was aerated with an aquarium pump and synthetic wastewater (Boeije et al., 1999). Influent was fed into the activated sludge compartment by a hydrostatic pump. Activated sludge contents flowed under a sluice gate into a 2 L sedimentation compartment. Effluent flowed out a spigot at the top of the water line while solids settled. Contents in the activated sludge compartment were well-mixed through aeration such that churning on the bottom of the activated sludge compartment drew solids settled from the settling compartment back into the activated sludge side.
Synthetic wastewater was prepared as described previously (Boeije et al., 1999). Influent was loaded into the activated sludge compartment at a rate of 5 mL/min. Activated sludge collected from the wastewater treatment plant in Stillwater, Oklahoma, USA, was used to seed reactors at the mixed liquor volatile suspended solids (MLVSS) concentration of 1,000 mg/L. MLVSS was measured every 2–4 days and maintained below 1,600 mg/L by periodically removing excess solids. Dissolved oxygen (DO) was measured with a YSI 5100 Benchtop DO meter and maintained at 2 to 4 mg/L. The pH of the reactors was measured daily with a SevenCompact pH probe (Mettler Toledo, Columbus, OH) and maintained between 6.9 and 8.1.
Ciprofloxacin was purchased from Sigma-Aldrich (St. Louis, MO). After 14 days of stable operation with chemical oxygen demand (COD) removal of >50%, ciprofloxacin was added into the influent at a concentration of 2.7 mg/L for the triplicate ciprofloxacin-challenged reactors, and another set of triplicate reactors continued to receive unamended influent to serve as controls. This concentration of ciprofloxacin was based on the prophylaxis dosage of 1 g/[person·day] that the CDC would prescribe to the affected population in the case of a suspected anthrax attack (CDC, 2017), divided by an average wastewater usage rate of 370 L/[person·day]. This scenario assumes no metabolism in the body (Rohwedder et al., 1990) and a wastewater treatment plant serving an entirely affected population undergoing prophylaxis for anthrax exposure. Reactors were operated for 14 days with ciprofloxacin-amended influent to analyze the immediate impacts of mass prophylaxis on reactor performance and the microbial communities.
COD and total organic carbon (TOC) were measured from the effluent of the reactors every 6 days or less utilizing the Hach COD2 Mercury-Free Digestion Vials and Hach TOC Reagent Set, respectively (Hach Company, Loveland, CO).
Microbial community analysis
Samples were collected from the sludge every week, concentrated by centrifugation (9,600 g for 5 min), and total DNA was extracted utilizing the PowerSoil DNA extraction kit (MoBio, Carlsbad, CA) according to the manufacturer's instructions. All isolated DNA samples were submitted to Molecular Research LP (Shallowater, TX) for polymerase chain reaction amplification of the V4 variable region of 16S rRNA genes with primers 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACVSGGGTATCTAAT-3′) (Caporaso et al., 2012) and bTEFAP© Illumina MiSeq 2 × 300 bp barcoded amplicon sequencing (Dowd et al., 2008). Analysis with the Molecular Research DNA pipeline consisted of the joining of sequences, depletion of barcodes, removal of sequences less than 150 bp long or with ambiguous calls, removal of chimera sequences detected with UCHIME (Edgar et al., 2011), and delineation of operational taxonomic units (OTUs) at a 97% identity level with UCLUST (Edgar, 2010). Final OTUs were taxonomically classified using BLASTn against a database derived from GreenGenes (McDonald et al., 2012), RDP11 (Cole et al., 2013), and NCBI (www.ncbi.nlm.nih.gov). The total number of reads varied from 16,931 to 105,497 per sample. The microbial community was analyzed with the OTU percentage read depth (i.e., relative abundance) data using non-metric multidimensional scale (NMDS) analysis using the VEGAN package in R (Oksanen et al., 2018). Illumina MiSeq sequence data are available from NCBI under Bioproject PRJNA574652.
The 16S rRNA gene sequence data were analyzed for relative abundance on OTU and higher phylogenetic levels. The sequences for OTUs exhibiting enrichment in the ciprofloxacin-challenged reactors were phylogenetically analyzed using MEGA X (Kumar et al., 2018). Highly identical sequences from isolates were recovered from NCBI BLASTn. Sequences were aligned with MUSCLE, and evolutionary history was inferred with the Neighbor-Joining Method (Saitou and Nei, 1987). A bootstrap test (500 replicates) was performed for statistical analysis (Felsenstein, 1985), and the evolutionary history was computed using the Maximum Composite Likelihood method (Tamura et al., 2004). A total of 273 bp was used in the alignment and phylogenetic tree construction.
High-performance liquid chromatography analysis
One milliliter samples were collected in triplicate from the reactor effluents and passed through a 0.22 μm filter at 1, 2, 4, 5, 8, and 10 h following the addition of ciprofloxacin. Samples were also taken from the influent before and just after antibiotic addition. Samples were stored at −20°C. After 7 and 14 days, the amount of ciprofloxacin was measured in the reactor solids. The extraction of ciprofloxacin from solids was carried out as previously described by Golet et al. (2002) with modifications. In brief, 0.2 g of wet sludge was placed in a 50 mL centrifuge tube with 4 g acid-washed sand and 4 mL of a phosphoric acid (50 mM, pH 2.0) and acetonitrile (ACN) mixture (1:1 v/v). The tubes were then placed in boiling water for 10 min, followed by sonication for 10 min (Solid State/Ultrasonic FS-28; Fisher Scientific, Pittsburgh, PA). The suspensions were then centrifuged (10,000 g, 10 min, 25°C) and a portion of the supernatant was filtered through a 0.22 μm filter before the determination of ciprofloxacin concentration.
Partitioning of ciprofloxacin between the influent synthetic wastewater media and the solids in activated sludge was studied in batch experiments as well. Activated sludge was oven dried (105°C, 24 h) and then amended at 2,000 mg/L into the synthetic wastewater influent containing 1, 2, or 3 μg/L ciprofloxacin, each in duplicate. Controls without amended sludge were analyzed as well. The amounts of ciprofloxacin in the liquid and solid phases were analyzed after shaking (200 rpms, 0.5 h, 25°C), followed by overnight stationary incubation, and 1 h of additional shaking before extraction.
Ciprofloxacin concentrations were determined by reverse phase high performance liquid chromatography (HPLC) analysis using a Waters HPLC Breeze system equipped with a 1500 Series HPLC pump, 2487 dual wavelength absorbance detector, and 717 plus autosampler (Waters, Milford, MA). The detection method was modified based on a method described by Vella et al. (2015). Separations were carried out using an XTerra C18 reverse phase 150 mm × 4.6 mm column with a 5 μm particle size containing 3-(chlorodimethylsilyl)-propyl-N-dodecyl carbamate-bonded amorphous organosilica ACN (Waters). The mobile phase contained 77% of 0.02 M phosphate buffer (pH 2.7) and 23% ACN under isocratic elution with a flow rate of 1.5 mL/min. The analysis was conducted by injecting 50 μL of supernatant and recording UV signals at 277 nm with a 10 min run time. The retention time of ciprofloxacin was ∼2.3 min. Standard control calibration curves were produced by analyzing ciprofloxacin at the following concentrations: 0.0, 0.5, 1.0, 2.0, 3.0, and 4.0 mg/L. All HPLC analyses were conducted in triplicate with standard calibration curves at the beginning point and ending point of each batch.
Reactor sampling, determination of colony-forming units, and isolation of lactose fermenting organisms
Reactors were sampled by withdrawing 3 mL of activated sludge just before ciprofloxacin addition, and after 24 and 72 h of ciprofloxacin exposure. The samples were serially diluted 10-fold in sterile phosphate-buffered saline, and colony-forming units per milliliter (CFU/mL) were determined by plating 100 μL of each dilution on nutrient agar (NA; Becton Dickinson and Company, Cockeysville, MD) and MacConkey agar (MCA; Becton Dickinson and Company). Individual isolates were obtained for identification and antibiotic susceptibility testing by streaking sludge from each reactor on MCA plates and incubating overnight at 37°C. From the colonies that were positive for lactose fermentation, four were picked at random from each reactor and restreaked on MCA plates. The species of each isolate was determined by matrix-assisted laser desorption/ionization time of flight mass spectrometry as described previously (Johnson et al., 2018). Working cultures of each isolate were prepared by inoculating a single colony into 3 mL of nutrient broth or Mueller Hinton broth (MHB; Becton Dickinson and Company) and incubated overnight (37°C, 200 rpm).
Antimicrobial susceptibility testing
Antimicrobial susceptibility profiles were determined by standard Kirby-Bauer analysis following Clinical and Laboratory Standards Institute guidelines (CLSI, 2018). Cells from overnight cultures were diluted in MHB to a final optical density at 600 nm (OD600nm) of 0.1 and spread uniformly across the surface of plates containing Mueller Hinton agar (MHA) using sterile cotton-tipped applicators. Antimicrobial disks (Oxoid, Thermo-Fisher Scientific, Waltham, MA) were then applied to the surface of each MHA plate, incubated for 24 h at 37°C, and the resulting zones of inhibition were measured.
Statistical analysis
Estimated CFUs from control and ciprofloxacin-challenged reactors were log10 transformed, tested for normality using the Shapiro-Wilk W-test, and compared by two-tailed Student's t-test. Prevalence of ciprofloxacin resistance and multidrug resistance was compared for each sampling time using Fisher's exact test (GraphPad QuickCalcs, https://www.graphpad.com/quickcalcs/contingency2/). Survival of organisms from control and amended reactors at each timepoint following challenge with ciprofloxacin was compared by Student's t-test as described above. All statistical analyses apart from the Fisher's exact tests were performed using JMP Student Edition v12 (SAS Institute, Cary, NC).
Results
Ciprofloxacin concentrations
Upon the introduction of ciprofloxacin into the influent, we expected to see the concentration of ciprofloxacin increase in the effluent. The amount of ciprofloxacin recovered in the influent was measured at 0.461 ± 0.022 mg/L, which is less than the 2.7 mg/L that was added. This suggests incomplete solubilization in the media and/or partial absorption of the ciprofloxacin to the influent container. Ciprofloxacin was first detected in effluent between 4 and 8 h after the initiation of the ciprofloxacin-challenge and reached 0.045 ± 0.022 mg/L by 10 h in the triplicate reactors (Fig. 1). This relatively low concentration in the effluent compared to the influent is likely due to sorption of ciprofloxacin onto the reactor sludge. At days 7 and 14, the amount of ciprofloxacin was 178 ± 30.3 and 198 ± 37.7 mg/kg wet sludge, respectively. Batch sorption experiments verified that ciprofloxacin absorbed heavily to the reactor solids (Table 1) compared to the media.

The ciprofloxacin concentration in the reactor effluent is shown over time in ciprofloxacin-challenged reactors. Each of the three reactors (□, Δ, ○) are shown separately and the error bars represent standard deviations of triplicate samples from each reactor.
Percentage Recovery of Ciprofloxacin Determined by High-Performance Liquid Chromatography in Batch Experiments
Reactor performance
Measurements of COD and TOC were used to monitor reactor performance over time. The influent media was measured to have a COD of 716.5 ± 10 mg/L and a TOC of 213 ± 19 mg/L. After 1 week of stabilization, the COD and TOC in the effluent was consistently measured between 239–394 and 40–80 mg/L, respectively (Fig. 2). After ciprofloxacin amendment commenced for the challenged reactors, both the effluent COD and TOC maintained at stable levels similar to that of the controls. Thus, the loading of ciprofloxacin at the concentration examined did not statistically affect the COD or TOC removal efficiencies of the reactors during this time frame. The MLVSS, DO, and pH were maintained similarly between the ciprofloxacin and control reactors (Supplementary Figs. S1–S3).
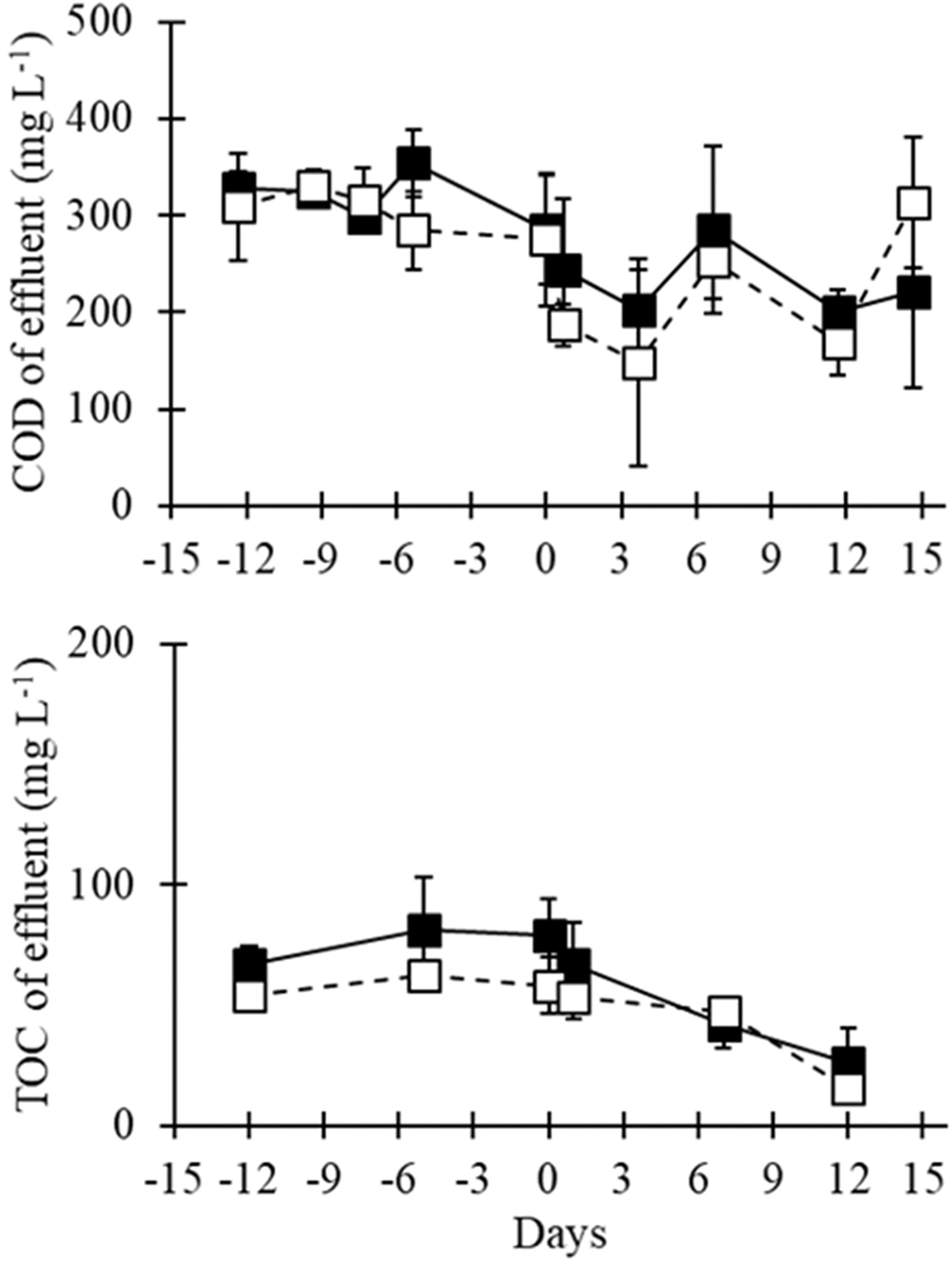
COD and TOC in the effluents of the ciprofloxacin-challenged reactors (solid squares/lines) and control (ciprofloxacin-free) reactors (open squares, dashed lines) over time. Day 0 denotes the initiation of ciprofloxacin loading. COD, chemical oxygen demand; TOC, total organic carbon.
Microbial community analysis
The microbial communities over time in the ciprofloxacin-challenged and control reactors were monitored with 16S rRNA gene analysis. An NMDS analysis of the OTU-level 16S rRNA gene data was used to determine if the overall microbial communities were altered during the ciprofloxacin challenge. The analysis indicated that the microbial communities changed in both the control and ciprofloxacin-challenged reactors over time, although the microbial communities in the ciprofloxacin-challenged reactors shifted more markedly (Fig. 3). By Day 7, the ciprofloxacin-challenged reactors diverged in community structure from the control reactors. This divergence became more pronounced by Day 14. To analyze how the communities changed, the evenness and richness of the communities were determined (Table 2). The richness, Shannon's diversity index and Shannon's evenness were lower in the ciprofloxacin-challenged reactors than in the controls on Days 7 and 14. On Day 14, the ciprofloxacin-challenged reactors had on average 31% fewer OTUs compared to the controls. The NMDS and diversity analyses both indicate that ciprofloxacin induced strong selection against several bacterial species.
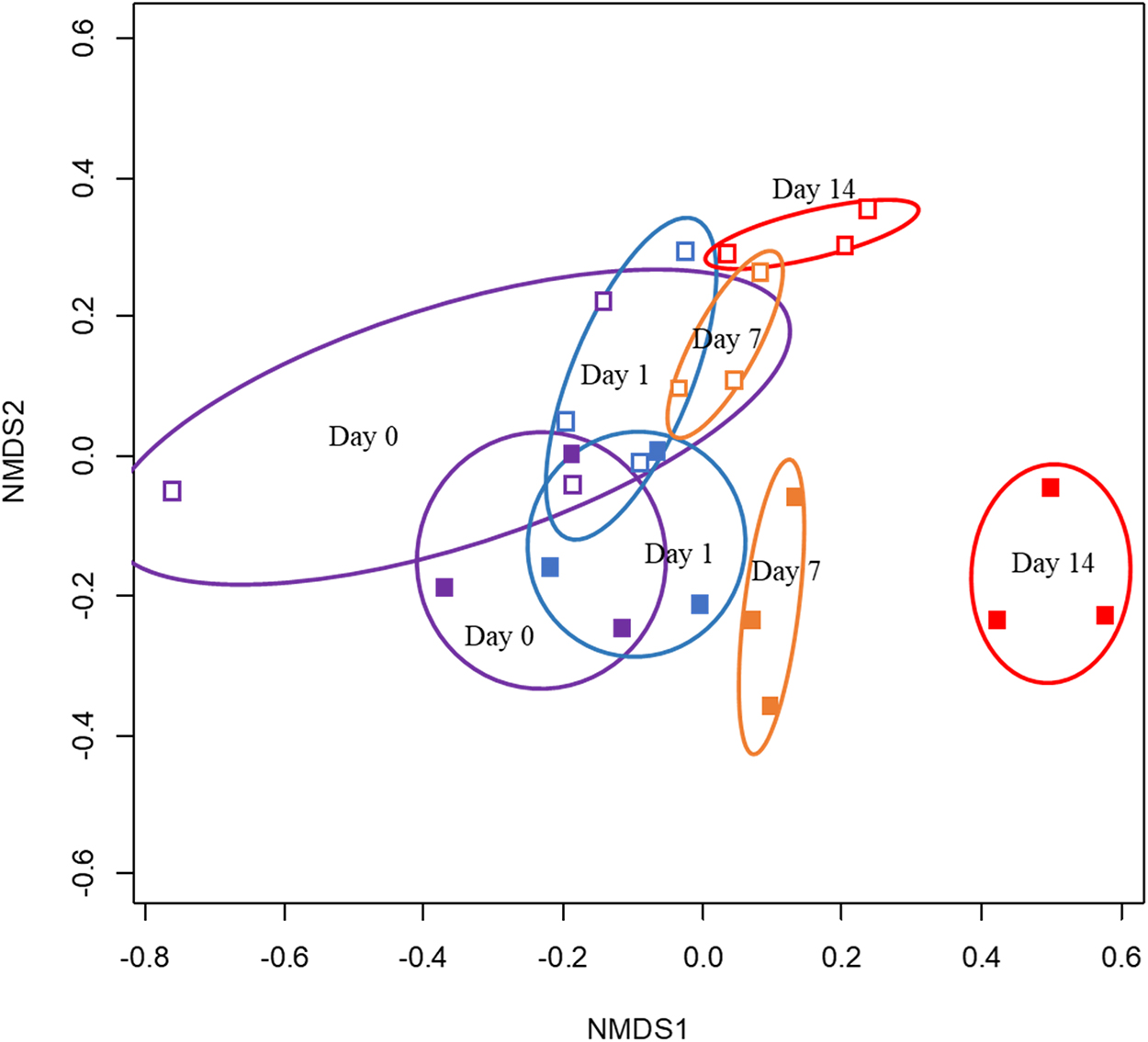
NMDS analysis (stress = 0.106) of the microbial community structure of the ciprofloxacin-challenged reactors (solid squares) and control reactors (open squares) at the initial time of ciprofloxacin introduction (Day 0, purple), and Day 1 (blue), Day 7 (orange), and Day 14 (red) after ciprofloxacin-challenge began. Ellipses refer to standard errors at the 95% confidence level. NMDS, non-metric multidimensional scale.
Richness, Shannon's Diversity Index, and Shannon's Evenness for Ciprofloxacin-Amended Reactors and Control Reactors
Student t-test, p ≤ 0.05.
OTU, operational taxonomic unit.
The phyla level relative read depth is shown in Supplementary Table S1 for DNA samples analyzed 5 days before, and at 7 and 14 days following ciprofloxacin addition to the influent. The phyla Proteobacteria and Bacteroides combined represented 74% to 87% of the relative read depth in all model reactor samples. At Day 14, four phyla (Gemmatimonadetes, Synergistetes, Cyanobacteria, and Verrucomicrobia) were significantly less abundant in the ciprofloxacin-challenged reactors compared to the control reactors (Student's t-test, p < 0.05). Of these four phyla, the read depth of Gemmatimonadetes demonstrated the greatest reduction (99.5%) from 1.65% ± 0.8% in the control reactors to 0.009% ± 0.004% observed in the in the ciprofloxacin-challenged reactors. The read depths of the Cyanobacteria, Synergistetes, and Verrumicrobia were 96.4%, 81.1%, and 17.4% less abundant in the ciprofloxacin-treated reactors compared to the control reactors, respectively.
Several phylogenetic groups were enriched in the 14 days in which ciprofloxacin was amended in the influent (Figs. 4, 5). The percentage read depth of OTUs exhibiting enrichment is shown in the Supplemental Information (Supplementary Table S2). OTUs related to the genera Inhella, Delftia, and Sphaerotilus within the Order Burkholderiales became enriched. Collectively, these OTUs increased to a relative read depth of 20.7% ± 5.6% by Day 7 in the ciprofloxacin-challenged reactors compared to 2.2% ± 3.2% in the controls. An OTU related to Emticicia spp. increased to 3.73% ± 2.29% by Day 14 in the ciprofloxacin-challenged reactors compared to 0.03% ± 0.02% in the controls. Two OTUs within Bradyrhizobiacea collectively increased to 0.77% ± 0.23% by Day 7 and 1.05% ± 0.8% and Day 14 and were closely related to Bosea spp. Three OTUs in the Microbacteriaceae also showed a modest increase to 0.73% ± 0.20% in the ciprofloxacin-challenged reactors compared to 0.19% ± 0.02% in the controls. Four OTUs within the Family Caulobacteraceae also showed 2.12% ± 1.17% on Day 14 in the ciprofloxacin-challenged reactors compared to 0.18% ± 0.07% in the controls, and an OTU closely related to Shinella spp. also showed some enrichment over controls. The Caulobacteraceae and Shinella spp. were highly variable between triplicate reactors on Days 0 and Day 1; thus, enrichment due to ciprofloxacin is less clear.
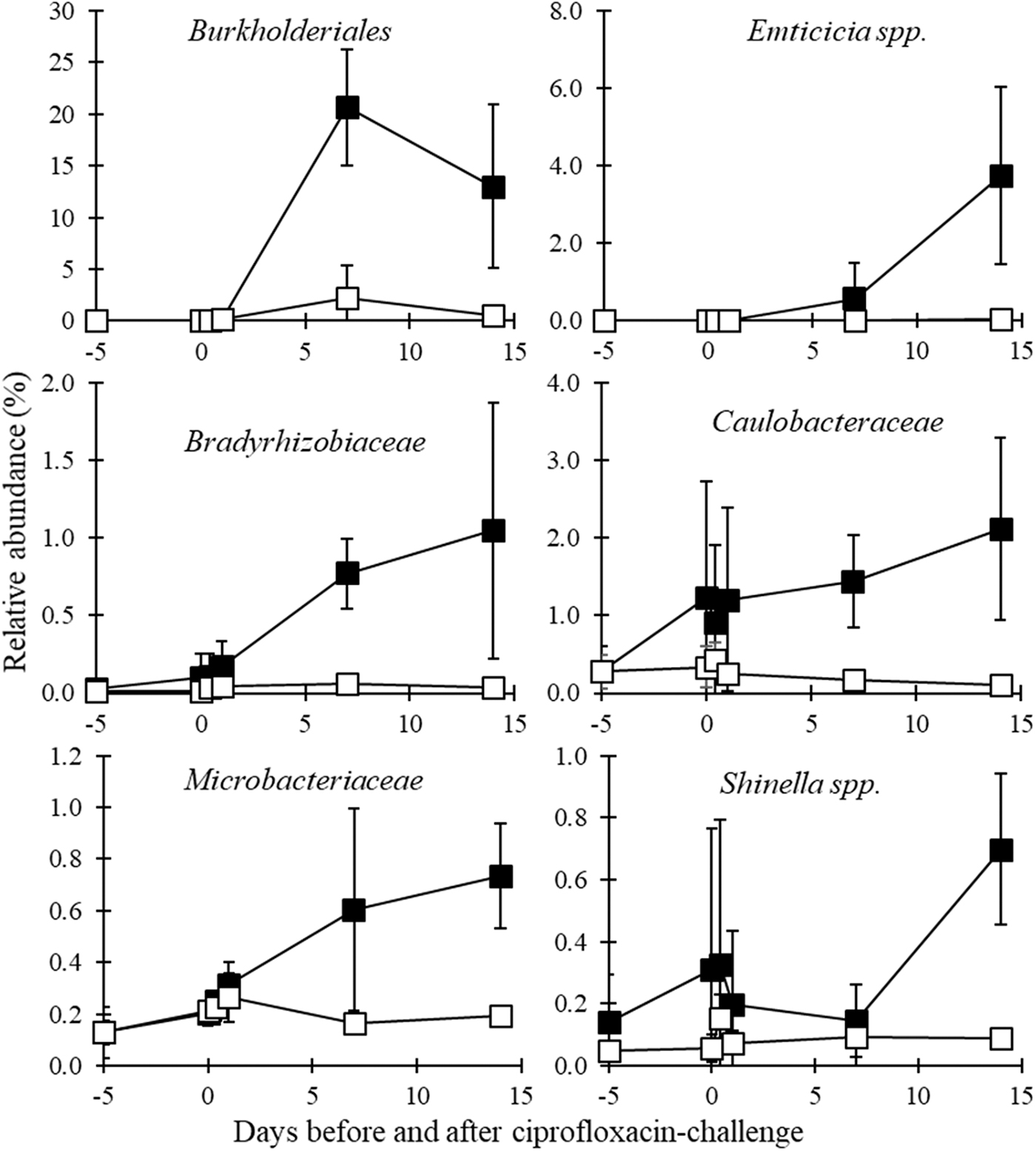
Bacterial groups found to have their relative abundance increase following ciprofloxacin challenge compared to controls based on 16S rRNA gene sequencing read depth (Student's t-test, p ≤ 0.05). Reactors with ciprofloxacin challenge are shown as black squares, while control reactors are shown with open squares.
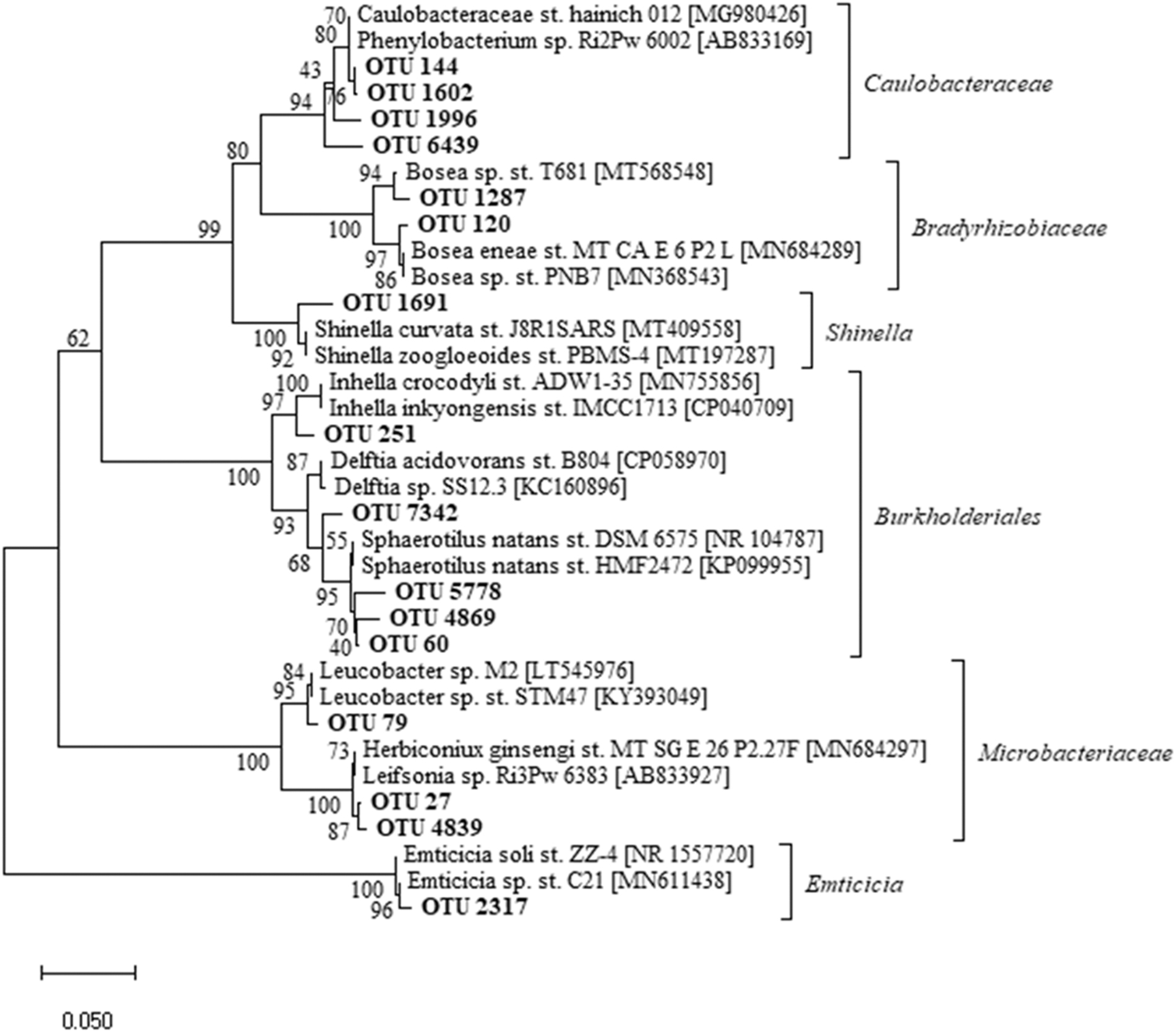
Phylogenetic characterization of OTUs showing enrichment in the ciprofloxacin-challenged reactors compared to controls. Percentage read depth information for Days 7 and 14 is provided for each OTU in the Supplementary Information (Supplementary Table S2). OTUs, operational taxonomic units.
Determination of CFUs, isolation of lactose fermenting organisms, and antimicrobial susceptibility
Bacteria isolates on selective media were used to analyze coliform counts and the spread of antimicrobial resistance. Ciprofloxacin-challenged reactors experienced a reduction in NA CFU counts compared to the control reactors following 24 h of ciprofloxacin-challenge (Student's t-test, p = 0.024; Table 3). No significant difference was observed between challenge and control reactors before ciprofloxacin challenge, nor following 72 h of ciprofloxacin challenge (Student's t-test, p ≥ 0.109 for both comparisons). No significant differences were observed for lactose fermenting coliforms on MCA at any time point (Student's t-test, p ≥ 0.289 for all comparisons).
Mean log10 Colony-Forming Units per Milliliter for Control and Ciprofloxacin-Challenged Reactors on Nutrient Agar and MacConkey Agar at 0, 24, and 72 h Following Ciprofloxacin Introduction in the Influent
Mean log10 CFUs/mL ± standard error.
Student's t-test.
CFUs, colony-forming units.
Although some randomly picked MCA colonies obtained from the control reactor sludge did express resistance to one or two β-lactam antibiotics, none expressed resistance to ciprofloxacin at any time point examined (Table 4). One C. freundii isolated at 72 h from the control reactor was multidrug-resistant (resistant to ≥ three classes of antibiotics) and demonstrated resistance to β-lactams, gentamicin, trimethoprim + sulfamethoxazole, and tetracycline.
Identity and Antimicrobial Resistance of Randomly Picked Colonies from the Control Reactors
AMC, amoxicillin; AMP, ampicillin; CN, gentamicin; I, Intermediate, R, Resistant; SXT, trimethoprim + sulfamethoxazole; TE, tetracycline; TIC, ticarcillin.
In the ciprofloxacin-challenged reactors, before ciprofloxacin, two randomly picked isolates displayed intermediate or full resistance to ciprofloxacin and one C. freundii isolate also displayed multidrug-resistance (Table 5). Following 24 h ciprofloxacin exposure, one C. freundii isolate and one Aeromonas caviae isolate displayed multidrug-resistance, including resistance to ciprofloxacin, five additional isolates demonstrated resistance to three β-lactams, and two additional isolates displayed resistance to a single β-lactam (Table 5). Eleven of the 12 randomly picked colonies isolated from the ciprofloxacin-amended reactors after 72 h were identified as C. freundii and expressed multidrug resistance, including ciprofloxacin resistance, and one C. freundii isolate demonstrated resistance to two β-lactams. The prevalence of ciprofloxacin resistance and multidrug resistance among the randomly picked colonies was greater in the ciprofloxacin-treated reactors compared to the control reactors (Fisher's Exact Test, p < 0.001).
Identity and Antimicrobial Resistance of Randomly Picked Colonies from the Ciprofloxacin-Challenged Reactors
AMC, amoxicillin; AMP, ampicillin; C, chloramphenicol; CIP, ciprofloxacin; CN, gentamicin; I, Intermediate, R, Resistant; SXT, trimethoprim + sulfamethoxazole; TE, tetracycline; TIC, ticarcillin.
Discussion
The ciprofloxacin concentration utilized in our study was higher than levels reported to be typical for wastewater treatment plants (Kulkarni et al., 2017). However, we did not find a significant impact on the operational parameters of our bench scale activated sludge reactors with regard to COD and TOC removal over 14 days. While ciprofloxacin reduced aerobic CFU counts at 24 h postchallenge, the CFU counts recovered to control levels by 72 h.
The microbial communities in the reactors were altered by ciprofloxacin, with less OTU richness and diversity, and a significant increase in 16S rRNA genes from the Order Burkholderiales, which includes a number of species that demonstrate intrinsic multiple antimicrobial resistance (Rhodes and Schweizer, 2016; Sfeir, 2018). For instance, Delftia spp. has intrinsic resistance to aminoglycoside antibiotics (Preiswerk et al., 2011). A recent study isolated a novel strain of Delftia spp. uniquely resistant to ciprofloxacin and other quinolones, which may have been the result of a single critical mutation on DNA gyrase (gyrA) (Kang et al., 2015). Sphaerotilus, long known in wastewater treatment for contributing to bulking (Dondero et al., 1961), has been found previously to increase in wastewater treatment communities with the antibiotics tetracycline (Du et al., 2018) and sulfadizine (Zhang et al., 2020). OTUs closely related to the genera Bosea, Leucobacter, and Phenylobacterium also increased in our study. These three genera have been detected in a ciprofloxacin-degrading culture (Liao et al., 2016), suggesting a possibility that growth in the reactors may be from the use of ciprofloxacin as a substrate.
With a full prophylaxis of 60 days (CDC, 2017) and the strong sorption of ciprofloxacin to the activated sludge solids found in this and other studies (Min et al., 2018), further research is required to determine the performance, and stability, of activated sludge reactors over a longer time period following the influx of relatively high antibiotic concentrations. In addition, other units of the wastewater treatment process, such as aerobic or anaerobic digestion, should also be analyzed as they may be more susceptible to performance upset. High concentrations of the bisphenol biocide triclosan, for example, have previously been shown to significantly impact the function of anaerobic digestion (McNamara et al., 2014).
The ciprofloxacin clearly selected for multidrug resistance in C. freundii in the activated sludge within our model. Our laboratory findings on the association between ciprofloxacin exposure and resistance to multiple antibiotics are similar to those produced by sampling hospital wastewater and wastewater treatment plants (Yim et al., 2012; Marti et al., 2014; Vaz-Moreira et al., 2015; Anssour et al., 2016). While C. freundii has been reported to be highly susceptible to ciprofloxacin (Samonis et al., 1987), fluoroquinolone resistance in clinical isolates of Citrobacter spp. is now established (Gesu et al., 2003). The mechanism by which C. freundii expresses resistance in our experiments will be studied further as both chromosomal mutations in genes encoding the targets of fluoroquinolones DNA gyrase (gyrA) and topoisomerase IV (parC), and the acquisition of plasmid or transposon associated qnr family quinolone resistance genes, have been reported in organisms exposed to ciprofloxacin in the environment (Yim et al., 2012; Marti et al., 2014; Vaz-Moreira et al., 2015; Anssour et al., 2016). The dosage used in this study is lower than the ciprofloxacin concentration required to select for gyrA mutations in Mycobacterium bovis, and higher than the concentration required to select for parC mutations in S. aureus in vitro (Dong et al., 1999). It has also been reported that Citrobacter carry specific qnr genes on plasmids, and multiple antimicrobial-resistant strains of C. freundii also can carry genes that encode β-lactamases, extended spectrum β-lactamases, as well as carbapenamases, which degrade penicillin and related antibiotics (Hanson and Sanders, 1999; Wang et al., 2000; Zhang et al., 2008; Maraki et al., (2017); Jacoby et al., 2011; Kanamori et al., 2011; Yim et al., 2012; Marti et al., 2014; Vaz-Moreira et al., 2015; Anssour et al., 2016; Liu et al., 2018a, 2018b). Infections with multidrug resistance strains of C. freundii have been associated with a higher rate of mortality in hospitals compared to infections caused by susceptible strains (Deal et al., 2007).
The numbers of the genera and species isolated on MCA were all found below or near detection limits in the 16S rRNA gene analysis, and enrichment of C. freundii was not observed. The reason why C. freundii appeared to be enriched in our antimicrobial resistance analysis was likely due to the selective isolation process on the growth media (MCA), which contains bile salts and crystal violet that inhibit the growth of most gram-positive organisms. The increase frequency of C. freundii being isolated on MCA under ciprofloxacin-challenge conditions at 72 h presumably is a result of the other strains selected for on MCA plates decreasing in comparison to the C. freundii.
Under antibiotic loadings associated with an emergency response to anthrax, our results suggest a rapid increase in antibiotic resistance and raises concern about potential for increased spread of antibiotic resistance genes and antibiotic-resistant bacteria throughout the environment. A recent study on antibiotic-resistant bacteria in discharge from a wastewater treatment plant reported that of the 109–1012 CFU per day, per inhabitant equivalent in the discharge, at least 107–1010 could express antibiotic resistance (Novo and Manaia, 2010). Some mitigation of this antibiotic-resistant bacterial discharge could be possible through increased disinfection (Oh et al., 2014); however, this approach would face logistical complications in an emergency situation.
Ciprofloxacin release downstream of wastewater treatment plants is likely to have direct environmental effects (Maul et al., 2006), including entering drinking water supplies downstream. Biological treatment to remove ciprofloxacin is known to be limited in municipal wastewater treatment plants (Lindberg et al., 2006; Amorim et al., 2014); however, a bacterial culture degrading ciprofloxacin has been characterized (Liao et al., 2016). Promotion of biodegradation may be made possible by the addition of magnetite or 2-bromoethanesulfonate in anaerobic conditions (Yang et al., 2017). Still, even if biodegradation of the ciprofloxacin can be induced, multiantibiotic resistance spread would still occur and require remediation. In addition, tertiary treatment systems such as advanced oxidation are able to degrade ciprofloxacin (Pereira et al., 2007). Such treatment options should be evaluated for deployment as a component of emergency response planning, preferably upstream of the biological treatment processes.
Conclusions
Research has focused on studying the effects of chronic widespread concentrations of antibiotics in the environment. However, it is also important to evaluate the risks and consequences associated with acute, high dosages of antibiotics. In the United States, ready-to-activate emergency response plans would release large amounts of antibiotics within days of a suspected anthrax attack. Our results indicate that such a release of the antibiotic ciprofloxacin strongly alters the microbial community. Decreases in aerobic CFU counts on nutrient media within 24 h indicate immediate selective pressure from ciprofloxacin on the community. This selective pressure leads toward lower community richness and diversity over 14 days, which may impede wastewater treatment efficacy and resiliency in the longer term. Increases of Burkholderiales and other bacteria groups, several of which have isolates known to cause disease, and the emergence of multidrug resistance indicate a potential to create follow-on public health crises. More research is required to develop and incorporate mitigation procedures into emergency response plans that will thwart the untoward negative effects of massive antibiotic release into wastewater treatment plants and the downstream environments.
Footnotes
Authors' Contribution
X.F. and W.L.J. contributed equally to this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Work was supported by the Oklahoma State University Vice President of Research office and the Oklahoma Agricultural Experiment Station.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.