
Abstract
As the ecological barrier of a lake environment, lakebanks with plant/soil/microorganism systems play roles in runoff sink function. However, without proper control, the pollutants absorbed by plants can remain in plant litter and return to the water body in the plant fall season, resulting in secondary pollution. To reuse plant litter and further enhance the ecological function of lakebanks, in this study, plant litter was collected to produce biochar, which was incorporated into the lakebank soil in a microcosm. Then, the results and mechanisms of nitrogen (N) interception/removal by biochar-amended soil (biochar-based lakebank) were examined in the plant fall season (December–February) and growth season (June–August), respectively. The results showed that: (1) in both plant fall and growth seasons, the biochar-based lakebank exhibited higher total nitrogen (TN) and nitrate N (NO3−-N) removal efficiency from runoff than natural lakebank, especially in the plant growth season; (2) in the plant growth season, the TN removal efficiency of biochar-based lakebank increased by 32.85%, 12.00%, and 18.97%, and NO3−-N removal efficiency increased by 218.19%, 67.22%, and 85.91% under low, medium, and high influent pollution loads, respectively, when compared with those of natural lakebank; and (3) biochar amendment into soil promoted plant growth and increased the abundance of microorganisms related to the N cycle such as nitrobacteria, nitrifying bacteria, and aerobic denitrifying bacteria, which can further enhance the ecological function of lakebank in N removal from runoff.
Introduction
During rainfall, nitrogen (N) loss with runoff threatens the river water quality (Song et al., 2017; Casazza et al., 2018), because phytoplankton can utilize N in different forms (e.g., nitrate-nitrogen [NO3−-N]) to cause water-quality degradation (Parsons et al., 2002; Beman et al., 2005). Impermeable surfaces can easily cause a large amount of runoff, which produces a high concentration of NO3−-N in the water body (Caraco and Cole, 1999; Kojima et al., 2011; Nagy et al., 2012). In addition, agricultural nonpoint source pollution can increase the pollution load of runoff, especially N pollution with the runoff entering water bodies causing eutrophication (Lusk et al., 2020). Currently, for runoff pollution control, the technologies commonly used around the world include constructed wetlands, detention ponds, percolation filters, and grass ditches.
Riparian buffer zone is an area where vegetation is naturally formed or artificially planted at the junction of water and land. Its function is to capture sediment, nutrients, organic matter, and chemical substances when runoff passes through the vegetation area and reduce the impact of surface runoff pollutants on the water body (Nigel et al., 2013). Therefore, the lakeshore plays an important role in the purification and restoration of lake water. With the deterioration of a large number of aquatic ecosystems owing to human activities, artificial construction of riparian buffer zones has become a universal measure to protect water quality (Osborne and Kovacic, 2006). In lakebank ecosystems, which are a key part of nonpoint source pollution control, pollutants are removed through three aspects, namely, soil, plants, and microorganisms. Soil can precipitate and absorb nutrients carried by the surface runoff. The strong adsorption function of soil is derived from the presence of a large amount of clay and silt (Zhang and Li, 2016). The soil and vegetation on the lakebank can form a biological and physical barrier, acting as a buffer for purification and filtration, and preventing some pollutants from entering the water body (Hefting et al., 2005). As the N content in plants accounts for 1–6% of their biomass (Mitra, 2015), the accumulation of plant biomass is one of the mechanisms of capturing excess nutrients in a bioretention setting. Microorganisms can play an important role in biological retention, because they affect many natural processes such as soil weathering, nutrient and carbon cycles, and symbiotic relationships, all of which affect the water quality of the biological retention system (Skorobogatov et al., 2020).
In the uncontrolled natural lakebank, plants absorb pollutants during the growth season (June–August), but the pollutants return to the soil in the form of plant litter and move into the water body with runoff, causing secondary pollution in the fall season (December–February). Therefore, regular harvesting of lakebank plants can achieve the goal of removing pollutants. However, previous studies have shown that harvesting should not be performed too frequently. For instance, harvesting once in the fall has been reported to lead to the highest N removal efficiency from the soil, whereas harvesting more than once a year has been found to significantly inhibit nitrifying bacteria, resulting in lower N removal efficiency (Qian et al., 2019).
With increasing awareness of environmental issues and scarcity of resources, there has been a growing interest in the use of biobased materials in various applications (Ashori and Nourbakhsh, 2010; Taneli et al., 2016). The preservation and use of plant straw are significant for sustainable agriculture. In addition, the rational utilization of plant stalks not only conserves soil and water, but also returns the remaining nutrients to the soil, water, and atmosphere, and reduces air pollution caused by traditional open burning (Jiang et al., 2015). Biochar is the product of crop waste, wood, and other biomass energy raw materials after pyrolysis under low oxygen condition (Mohan et al., 2014; Tripathi et al., 2016). When biochar is incorporated into soil, it can improve soil structure and increase soil water holding capacity (Mangrich et al., 2015) as well as reduce nutrient loss (Uzoma et al., 2011). Furthermore, biochar can enhance water permeability and reduce surface runoff (Peng et al., 2015; Sang et al., 2015). When applied to soil, biochar exhibits a higher stability against decomposition and an excellent ability to absorb ions, when compared with other forms of soil organic matter (SOM), owing to its higher surface area, negative surface charge, and charge density. However, studies on whether amendment of biochar from harvested plant litters into lakebank soil can enhance its N removal capacity have rarely been reported.
To verify the feasibility of this notion, biochar prepared from plant litter in lakebanks was incorporated into the soil in a lakebank microcosm to explore the effect of biochar amendment on N removal from runoff by lakebanks. In addition, the three aspects, namely, soil interception, plant absorption, and microbial utilization, which contribute to N removal, were analyzed and discussed to clarify the mechanism. The results obtained provide evidence for the feasibility of plant litter recycling measures in lakebank and help to further improve the N removal efficiency of lakebank soil.
Materials and Methods
Design and operation of the lakebank microcosm
To study the effect of biochar-based lakebank on the interception/removal of N in runoff, we constructed a lakebank microcosm system. A structure with two disconnected slopes with a slope angle of 45° was established. An overflow tank was arranged to distribute water evenly, and a water outlet was set at the bottom to collect water. The microcosm simulation diagram and real image are shown in Fig. 1.
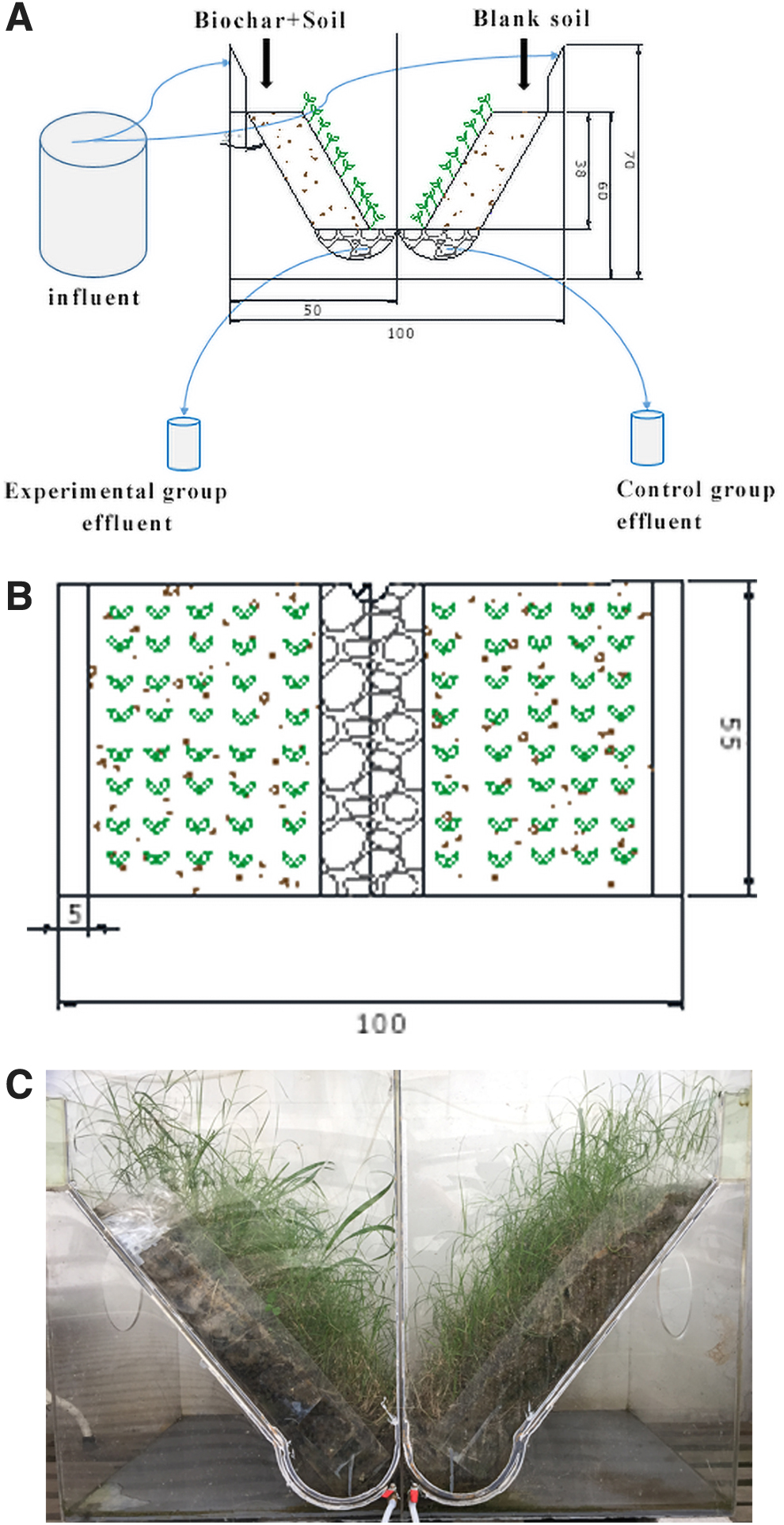
The microcosm simulation diagram side view
According to the size of the microcosm, two soil containers were designed. The size of each container was 55 cm × 48 cm × 10 cm (length × width × height). In the control group, natural dried soil was added. In the experimental group, natural dried soil mixed with 4% biochar was added. The weight of the soil in each container was 35 kg. The two groups of soil were placed outdoors, but were protected from natural precipitation and watered every day. After 20 days of stabilization, equal amounts of Zoysia matrella seeds were sown into the soil of the two groups and stored under the same conditions (sunlight exposure and protection from natural precipitation). After the Z. matrella seeds germinated and grew for a period of time, the soil drawers were placed in the microcosm. According to the preset pollution load and flow, water was fed into the two groups of overflow tanks under the same conditions of sunlight exposure and protection from natural precipitation. In this study, the plant fall season was considered from December 2, 2017, to February 3, 2018, lasting for 63 days, and the plant growth season was from June 20, 2018, to August 22, 2018, and lasting likewise for 63 days.
The influent flow was determined according to the monthly precipitation and the days of monthly precipitation in Shanghai from 2011 to 2015 (Shanghai Municipal Bureau of Statistics, http://tjj.sh.gov.cn/). The maximum average daily rainfall during 2011–2015 was 41.67 mm, and the catchment area of each soil drawer (calculated by horizontal projection, with slope angle of 45°) was determined to be 0.16 m2. According to the maximum average daily rainfall of Shanghai from 2011 to 2015 and the rainwater catchment area, the influent flow was calculated as follows (Kadlec, 2010):
Q, influent flow, mL/min;
k, safety factor, considered 1.30;
S, catchment area of each soil drawer, m2;
q, maximum average daily rainfall in Shanghai from 2011 to 2015, mm.
The influent flow of each drawer was determined to be 6.00 mL/min. According to the rainwater quality in Shanghai and the local runoff quality (Shanghai Environmental Monitoring Center, www.semc.gov.cn/), the data obtained were sorted out and summarized to determine the total nitrogen (TN) and NO3−-N concentrations under low, medium, and high pollution loads (Table 1).
The Concentration of Total Nitrogen and NO3-N in Influent Under Low, Medium, and High Load
TN, total nitrogen.
Soil, biochar, and plants
The soil was collected from 0- to 20-cm soil layer of the lakebank in Chongming Island, Shanghai, China, and the plant rhizome, gravel, garbage, and other sundries were removed. The soil type was newly reclaimed poor soil, with a soil bulk density of 1.22 g/cm3, TN content of 283.56 mg/kg, available phosphorus content of 22.13 mg/kg, and organic carbon content of 4.50 g/kg.
Biochar was produced from Z. matrella litters on lakebank by pyrolysis at 450°C in a reflux carbonization furnace designed by the Energy and Mechanical Engineering Faculty of Tongji University, China, with an energy self-sustaining system that uses the refluxed combustible components produced during biochar production to continuously generate heat, thus meeting environmental and economic sustainability. The dried plant litters were put into the furnace, and the initial temperature of 200°C was slowly increased every 1.5 h by 50°C to 450°C in an N2 atmosphere. The furnace temperature was maintained for 30 min at 450°C until no volatile chemical release was evident. Afterward, the products were cooled with N2 without water rinsing (Cao et al., 2016). Next, the biochar products were weighed and ground to 1- to 2-mm particle size for further analyses and application. The chemical properties of straws and their biochars were analyzed (Table 2).
Basic Properties of Straw and Straw Biochar
Mean ± standard deviations (n = 3).
Z. matrella, a representative lakebank plant, is the most cold-tolerant grass among the warm-season turf grasses. It has high ornamental value and water resistance, is susceptible to only a few pests and diseases, and is widely distributed in the tropical and subtropical areas of Asia and Oceania (Xuan et al., 2013). The Z. matrella seeds used in this study were purchased from the Wanbao landscape nursery stock farm, Shuyang County, Suqian City, Jiangsu Province, China.
Sample collection
Water sample collection
During the operation of the microcosm, the influent and effluent were collected every day. After completing the sample collection under one level of pollution load, the influent pollution load was changed to another level and the first sample collection under the new load level was conducted after 24 h of alteration.
Soil sample collection
Soil samples were collected before the start of the experiment and after operation of the microcosm under low, medium, and high pollution load. The soil (5–10 cm) was collected through five-point sampling, the plant roots and other sundries in the soil were removed, and the fresh soil was evenly mixed. A part of the fresh soil samples was stored in a refrigerator at 4°C for the analysis of fresh soil indices, including NO3−-N content and soil microbial community structure. Another part of the soil samples was naturally dried and sieved through a 60-mesh sieve for the analysis of dried soil indices, including TN concentration. The rest of the soil samples were stored in a refrigerator at −80°C for the analysis of soil microbial community structure.
Plant sample collection
During the plant growth season, after one cycle of operation of the microcosm, all the new plants in the soil drawer were collected using clean scissors, leaving behind 1–2 cm of roots. The aboveground part of the collected fresh plants was rinsed with deionized water and dried at room temperature.
Analytical method
Water quality
The TN concentration was determined according to protocols given in the Standard Methods (HJ 636-2012). The concentration of NO3−-N was ascertained by the hydrazine sulfate reduction method (Liu et al., 2002) using a 50 mL glass colorimetric tube containing a certain volume of pretreated water sample and sealed with a plug. If the NO3−-N concentration in the sample was too high, the volume of the sample was appropriately diluted.
Soil TN and NO3−-N contents
Soil TN content was determined by potassium persulfate oxidation-ultraviolet spectrophotometry according to a previously described method (Tang et al., 2011; Hu et al., 2014). Soil NO3−-N content was measured using a diffusion-conductivity analyzer (Gentlea et al., 2011).
Soil microbial community structure
The total DNA of the soil microorganisms was obtained by extracting the DNA from ∼0.5 g of the soil samples using the PowerSoil DNA Isolation Kit (Mo Bio Laboratories, Carlsbad, CA) according to the manufacturer's protocol. DNA concentration was determined using a spectrophotometer (NanoDrop ND-2000; Thermo Scientific, Waltham, MA). The Solexa technique was used to detect the DNA based on sequencing by synthesis and deoxyribonucleoside triphosphate reversible termination of chemical reactions (Quail et al., 2008). High-throughput sequencing was performed by the Shanghai Meiji Biomedical Technology Co. Ltd. (China). The prepared microbial genomic DNA was purified for the analysis of 16S diversity of the prokaryotic microorganisms. The detection region was 16S rDNA V1–V3, and the DNA was detected using the Illumina MySeq2 × 300 bp high-throughput sequencing platform. The sequencing results were analyzed on the ISanger cloud platform.
Plant biomass and TN content
Plant biomass refers to the total dry weight of one or more plants in a unit area or volume of habitat over a certain period of time. The plants were dried, crushed, and sieved over a 200-mesh sieve. Then, the TN content was determined using an automatic elemental analyzer, Vario EL III (Elementar Analysensysteme Comp., Hanau, Germany) (Chen et al., 2013).
Statistical analyses
The results of all the assays (except for sequencing) are presented as the averages of triplicate measurements, and the sequencing data presented herein are the comprehensive results of three samples. All the figures were performed using either SigmaPlot 10.0 or Microsoft Excel 2010. Statistical analyses were carried out using one-way analysis of variance with SPSS software 24.0. Multiple-comparison test in SPSS was used to further examine the significant differences (p < 0.05) among treatments.
Results
Enhanced N interception/removal of biochar-based lakebank
TN interception effect of biochar-based lakebank soil
The TN interception effects of the microcosm in the fall season and growth season are shown in Fig. 2. In the fall season, the TN removal efficiency was 7–34%, 4–22%, and 1–3% under low, medium, and high pollution loads in the natural lakebank, while it was 5–35%, 8–21%, and 1–6% in the biochar-based lakebank (Fig. 2A). Overall, the TN removal efficiency decreased as the pollution load increased. The average TN removal efficiency was 20.03%, 14.82%, and 5.31% in the biochar-based lakebank under low, medium, and high pollution load, while it was 15.21%, 7.83%, and 2.16% in the natural lakebank (Fig. 2C).
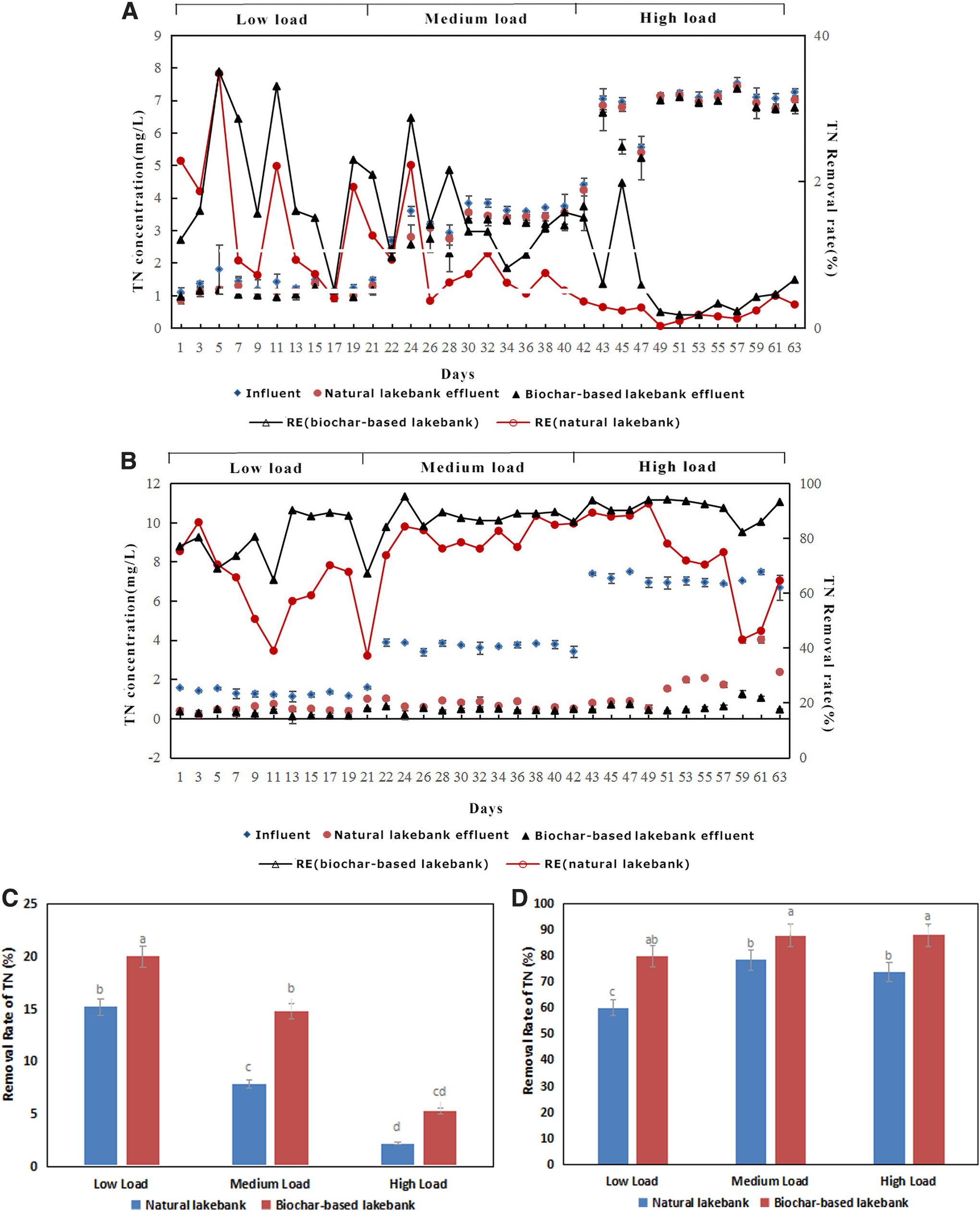
The removal rate of TN in the microcosm in fall season
In the growth season, the TN removal efficiency in the natural lakebank was 50–85%, 73–88%, and 43–92% under low, medium, and high pollution loads, while it was 64–90%, 84–95%, and 82–94% in the biochar-based lakebank (Fig. 2B). The TN removal effect was significantly better (p < 0.05) in the growth season than that in the fall season, indicating that the growth of plants significantly improved (p < 0.05) the TN removal efficiency. The average TN removal efficiency was 79.80%, 87.76%, and 87.88% in the biochar-based lakebank, and 60.07%, 78.36%, and 73.87% in the natural lakebank under low, medium, and high pollution loads, respectively (Fig. 2D). The TN removal efficiency in the natural lakebank significantly declined (p < 0.05) under high pollution load, implying that the interception of TN in the natural lakebank had reached saturation. In contrast, the TN removal efficiency in the biochar-based lakebank did not decline, but exhibited an increasing trend under high pollution load, suggesting that a biochar-based lakebank can prolong the effect of the system to intercept/remove TN in runoff and has a higher buffer capacity.
NO3−-N interception effect of biochar-based lakebank soil
Among the various forms of N, NO3−-N is negatively charged, and thus cannot be adsorbed by the negatively charged colloids that dominate most soils; therefore, N fertilizer leaching in soil mainly occurs in the form of NO3−-N (Weil and Brady, 2017). N leaching in soil not only results in wastage of fertilizer, but also produces the nitrate pollution in water bodies (Fu et al., 2017; Huang et al., 2017; Yang et al., 2017). Hence, we further explored the N interception effect of the biochar-based lakebank system.
In the fall season, the NO3−-N removal efficiency in the natural lakebank was 5–24%, 14–24%, and 1–14%, whereas that in the biochar-based lakebank was 3–28%, 9–32%, and 2–15% under low, medium, and high pollution loads, respectively (Fig. 3A). As the pollution load was increased, the NO3−-N removal efficiency in the natural lakebank and the biochar-based lakebank essentially remained stable. With a further increase in the pollution load to high, the NO3−-N removal efficiency showed a significant decline (p < 0.05). The average NO3−-N removal efficiency in the natural lakebank was 9.15%, 9.44%, and 8.50%, which were almost stable at the same level under low, medium, and high pollution loads, respectively. In the biochar-based lakebank, the average NO3−-N removal efficiency was 15.78%, 18.28%, and 9.18% under low, medium, and high pollution loads, respectively (Fig. 3C), with the highest and lowest NO3−-N removal efficiency noted under medium and high pollution loads, respectively, when compared with those observed in the natural lakebank.

The removal rate of NO3−-N in the microcosm in fall season
In the growth season, the NO3−-N removal efficiency was 90–100%, 93–100%, and 66–99% in the biochar-based lakebank, and 3–99%, 1–92%, and 8–96% in the natural lakebank under low, medium, and high pollution loads, respectively (Fig. 3B). It can be seen that the NO3−-N removal effect was stronger and more stable in the biochar-based lakebank than the natural lakebank under all the three pollution loads, whereas that in the natural lakebank considerably fluctuated and the effluent quality was extremely unstable. The average NO3−-N removal efficiency in the natural lakebank was 31.28%, 58.79%, and 47.71% under low, medium, and high pollution loads, respectively, which was relatively low. In contrast, in the biochar-based lakebank, the average NO3−-N removal efficiency was 99.53%, 98.31%, and 88.70% under low, medium, and high pollution loads (Fig. 3D), respectively, which was superior to that noted in the natural lakebank during the growth season.
Enhanced N interception/removal pathway of biochar-based lakebank
To explore the enhanced N removal mechanism of the biochar-based lakebank, three aspects, namely, soil interception, plants absorption, and microbial utilization, which contribute to N removal, were analyzed and discussed.
Plant-absorbed TN in biochar-based lakebank and natural lakebank
In the growth season, the plant biomass was 26.89 and 47.73 g/m2 and the TN content in the plants was 35.79 and 31.00 mg/g in the natural lakebank and biochar-based lakebank, respectively (Table 3). The amount of TN removed by plant absorption was 962.53 mg/m2 in the natural lakebank and 1479.54 mg/m2 in the biochar-based lakebank. The TN removal efficiency of the biochar-based lakebank was 53.71% higher, when compared with that of the natural lakebank, and the TN content in plants in the biochar-based lakebank was lower than that in the natural lakebank. In contrast, the biomass of plants in the biochar-based lakebank was much higher than that in the natural lakebank, which led to increased TN absorption in the biochar-based lakebank. Overall, the plants in the biochar-based lakebank absorbed more TN than those in the natural lakebank because biochar amendment enhanced the growth of plants and improved plant biomass.
The Plant Biomass of Microcosm, Plant Biomass per Square Meter, Total Nitrogen Content of Plant, the Amount of Total Nitrogen Removed by Plant Absorption per Square Meter and Its Total Amount in Growth Season, with Biochar-Based Lakebank and Natural Lakebank
Soil-intercepted TN in biochar-based lakebank and natural lakebank
Before operating the microcosm, the TN content was 0.26 and 0.28 mg/kg in the natural lakebank and biochar-based lakebank, respectively; thus, the two exhibited only slight differences. However, after one operation cycle in the fall season, the TN content was 0.39 mg/kg in the natural lakebank and 0.82 mg/kg in the biochar-based lakebank. The amount of intercepted TN of the microcosm was 28.70 mg in the biochar-based lakebank, which was 110.26% higher than that in the natural lakebank (13.65 mg). Furthermore, after one operation cycle in the growth season, the TN content was 0.71 and 1.17 mg/kg in the natural lakebank and biochar-based lakebank, respectively (Fig. 4). The amount of intercepted TN of the microcosm was 40.95 mg in biochar-based lakebank, which was 64.79% higher than that in the natural lakebank (24.85 mg). The higher TN content in the biochar-based lakebank, when compared with that in the natural lakebank, indicated that the soil of the biochar-based lakebank had a better interception effect on TN than that of the natural lakebank. In the fall season, the TN content in the soil of the biochar-based lakebank significantly increased (p < 0.05), when compared with that in the natural lakebank. However, in the growth season, both the biochar-based lakebank and natural lakebank showed an increase in soil TN content, but the increase was only slightly higher in the biochar-based lakebank, when compared with that during the fall season. This finding implied that the biochar-based lakebank had absorbed a large amount of TN during the fall season, resulting in gradual saturation of its TN adsorption capacity.
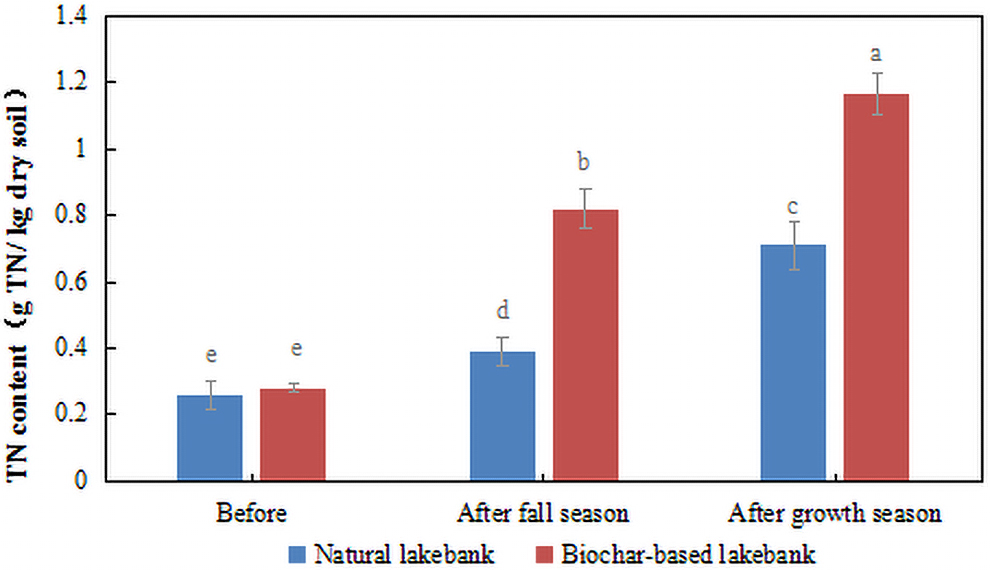
TN content in soil (g TN kg−1 dry soil) before, after cycle in fall season, and after cycle in growth season, respectively, with soil of biochar-based lakebank and soil of natural lakebank. Bars with the same letter(s) within each period are not significantly different at p < 0.05.
Estimation of the microbial effect on TN removal
The TN removal in the microcosm was first calculated, followed by the amount of TN removed by plants and soil, so as to estimate the quantity of TN removed by microorganisms, as follows:
Ainfluent, TN amount in influent, mg;
Aeffluent, TN amount in effluent, mg;
Aplant, amount of plant-absorbed TN, mg;
Asoil, amount of soil-intercepted TN, mg;
Amicroorganism, amount of microorganism-removed TN, mg;
Q, inflow, 0.006 L/min;
T, sampling interval time, 2 × 24 × 60 = 2,880 min;
Cinfluent(n), TN concentration in influent on day n, mg/L;
Ceffluent(n), TN concentration in effluent on day n, mg/L;
B, biomass of plant, g;
Cplant, TN content in plant, mg/g;
Msoil, weight of soil, 35 kg;
Csoil, TN content in soil after the whole cycle, mg/kg;
Cbefore, TN content before operation, mg/kg.
In the natural lakebank, the total amount of TN removal was 1838.23 mg after two operation cycles in the fall season and growth season. The amount of TN absorbed by plants was 254.11 mg, while that intercepted by soil was 24.85 mg. Thus, the TN amount removed by microorganisms was 1559.27 mg. In the biochar-based lakebank, the total amount of TN removal was 2269.08 mg after two operation cycles, which was 23.44% higher than that observed in the natural lakebank. The amount of TN absorbed by plants was 390.60 mg and that intercepted by soil was 40.77 mg. Accordingly, the quantity of TN removed by microorganisms was 1837.71 mg. The amount of TN removed by plants, soil, and microorganisms in the biochar-based lakebank was 53.71%, 64.06%, and 17.86% higher than those noted in the natural lakebank, respectively. To calculate the amount of TN removed by plants, soil, and microorganisms per square meter of the lakebank, the TN removal by these three aspects was, respectively, divided by the lakebank area, which was 0.264 m2 (Fig. 5). The results revealed that plants, soil, and microorganisms contributed to 13.82%, 1.36%, and 84.82% of TN removal in the natural lakebank, and 17.21%, 1.80%, and 81.99% of TN removal in the biochar-based lakebank, respectively. Thus, in both the natural lakebank and biochar-based lakebank, microorganisms played the predominant role in N removal, followed by plant absorption and soil interception. Besides, the total amount of TN removal in the biochar-based lakebank was higher than that in the natural lakebank, suggesting that the biochar-based lakebank had a better removal effect on TN than the natural lakebank. As microorganisms are the major contributors to TN removal, the differences in microbial characteristics of the natural lakebank and biochar-based lakebank were further analyzed to explore the enhanced mechanism of the biochar-based lakebank for N removal.
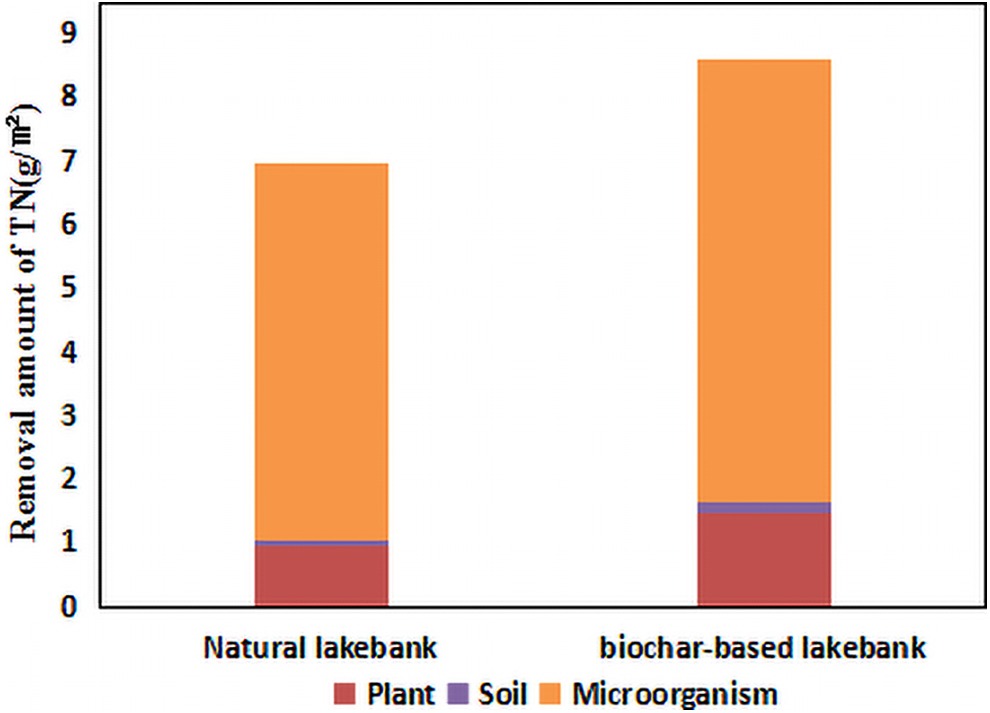
Removal amount of TN by plant/soil/microorganism per square meter (g/m2) calculated from average data in the whole cycle, with biochar-based lakebank and natural lakebank.
Differences in microbial community structures of biochar-based lakebank and natural lakebank, and the relationship with N removal efficiency
Richness and diversity of the microbial community in biochar-based lakebank and natural lakebank
The Ribosomal Database Project (RDP) classifier Bayesian algorithm was used to compare 97% of Operational Taxonomic Units (OTUs) representative sequences to obtain the corresponding species classification information of each OTU. The community composition of the samples was determined at multiple taxonomic levels and species annotation was conducted by the silvera128/16s bacteria database. Analysis of sample diversity (Alpha diversity) reflected the richness and diversity of the microbial community. Community richness represented the number of species, and community diversity indicated the uniformity of species distribution. The characterization indices used in this study were Chao, Ace, Shannon, and Simpson. Higher Chao and Ace values signified higher community richness, whereas a higher Shannon value and lower Simpson value implied higher community diversity. In general, the richness and diversity of soil microbial community were slightly higher in the natural lakebank, when compared with those in the biochar-based lakebank (Table 4).
Community Abundance and Diversity Indices for Soil Microorganisms in the Samples with 4% Biochar-Based Lakebank and Natural Lakebank
Venn diagrams can be used to determine the number of common and unique species (such as OTUs) in multiple groups or samples, which can intuitively indicate the similarity in species composition. The number of common species was the highest, reaching 428, while the number of unique species was 32 in the biochar-based lakebank, which was higher than that in the natural lakebank (number of unique species, 22) (Fig. 6).
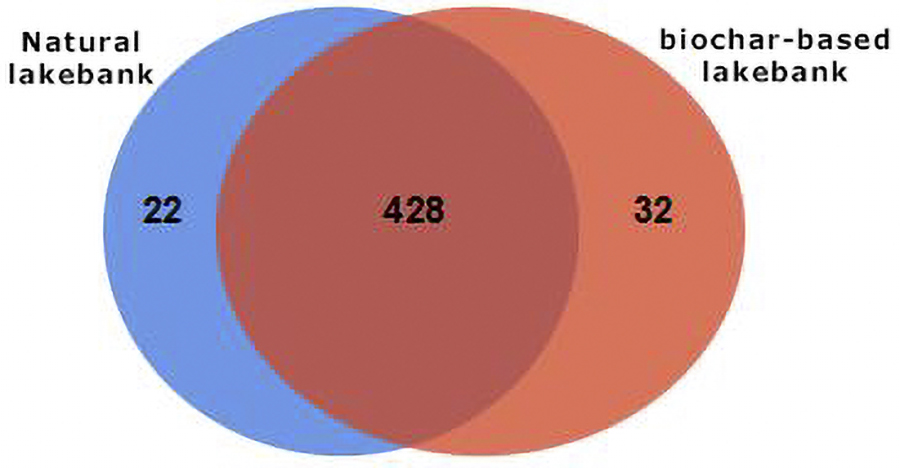
Venn diagram of unique and shared operational taxonomic units in the soil at genus level, with biochar-based lakebank and natural lakebank. The overlapping part represents shared operational taxonomic units and the nonoverlapping part represents unique operational taxonomic units.
Although the microbial species in the soil of the biochar-based lakebank and natural lakebank were mostly similar, certain differences were noted. In addition to microbial species, the abundance of dominant bacteria and the characteristics of specific bacteria related to N removal also affected the N removal efficiency. To further clarify the enhanced microbial N removal mechanism, the specific microbial species composition and proportion of dominant bacteria were analyzed.
Composition of microbial species at the phylum and genus levels
The composition of microbial species with high relative abundance at the phylum level was similar in the natural lakebank and the biochar-based lakebank. The top 3 microbial species at the phylum level with the highest relative abundance were Proteobacteria, Acidobacteria, and Actinobacteria, which accounted for 37.32%, 16.20%, and 11.27% in the natural lakebank, and 42.96%, 8.45%, and 16.90% in the biochar-based lakebank, respectively (Fig. 7). The relative abundance of Actinobacteria and Proteobacteria in the biochar-based lakebank was higher, whereas the abundance of Acidobacteria was lower, when compared with those in the natural lakebank.

Taxonomic composition of soil microbial community at phylum level in the samples, with biochar-based lakebank and natural lakebank. Different colors represent different taxa, the abscissa shows the species abundance of the taxon.
The composition of microbial species at the phylum level in the biochar-based lakebank and the natural lakebank did not show significant differences (p < 0.05). To further understand the effect of microorganisms on N removal with biochar amendment, we explored the microbial community composition at the genus level.
The heatmap of the microbial community composition at the genus level (Fig. 8) revealed that microbial species with high abundance both in the natural lakebank and biochar-based lakebank were Acidobacteria, Nitrosomonadaceae, Nitrospira, Gemmatimonadaceae, Methylophilaceae, and Cyanobacteria. Furthermore, the abundance of Nitrospira, Nitrosomonadaceae, and Methylophilaceae in the biochar-based lakebank was higher than that in the natural lakebank, which indicated the higher abundance of microorganisms involved in nitrification and denitrification in the biochar-based lakebank. This may be an important reason for the better N removal effect of the biochar-based lakebank, when compared with that of the natural lakebank. In contrast, the abundance of some heterotrophic bacteria such as Actinobacteria, Sphingomonadales, Mesorhizobium, and Nocardioides was lower in the biochar-based lakebank, when compared with that in the natural lakebank.
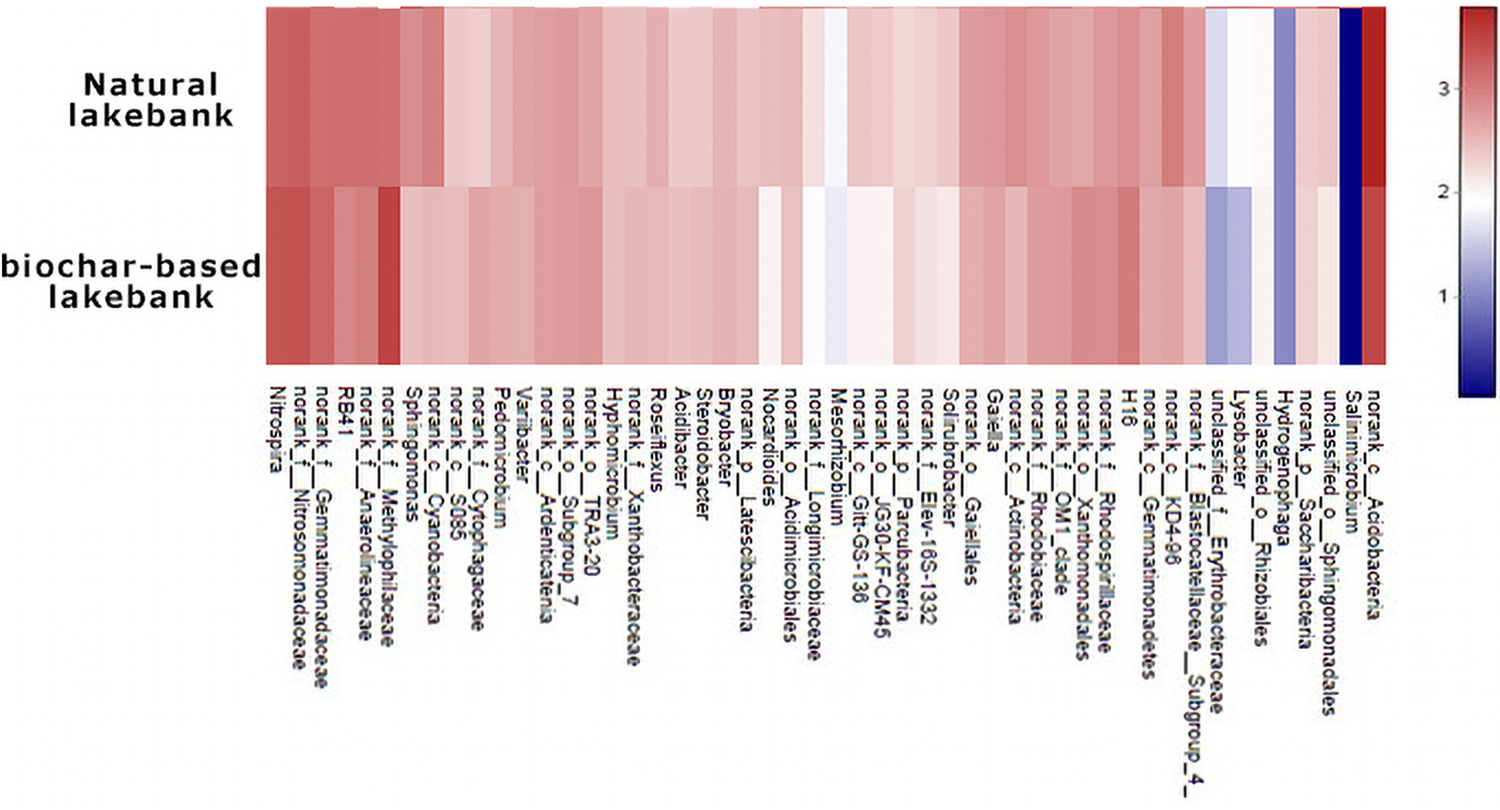
The heatmap of the community composition of soil microbial community at the genus level in the samples, with biochar-based lakebank and natural lakebank. Different colors represent different abundances of the taxa. Warmer color represents higher abundance. The abscissa shows the taxon.
In summary, the biochar-based lakebank exhibited only slight difference in microbial diversity and abundance, when compared with the natural lakebank. Although the microbial community structure in both the lakebanks was similar, there were obvious differences in the proportion of dominant bacteria, which may be an important reason for the difference in the microbial N removal efficiency and plant growth and absorption in the biochar-based and natural lakebanks.
Discussion
Biochar amendment promotes plant growth to enhance the N removal effect of lakebanks
In the growth season, the amount of TN absorbed by plants was 390.60 mg in the biochar-based lakebank, which was more than 50% higher than that in the natural lakebank (254.11 mg). Although the TN content was slightly lower in the biochar-based lakebank, when compared with that in the natural lakebank, the biomass was much higher in the biochar-based lakebank, indicating that biochar amendment can promote the growth of plants and improve their biomass, resulting in enhanced TN absorption. It has been reported that higher biomass has a certain dilution effect on plant N content (Marchão et al., 2018), which may be the reason for the lower TN content in plants in biochar-based lakebanks.
The addition of biochar to soils has been noted to result in increased aboveground productivity, crop yield, soil microbial biomass, and rhizobial nodulation (Biederman and Harpole, 2012). Besides, biochar can improve the soil water-holding capacity (Laird et al., 2010), facilitating plant biomass gain (Kammann et al., 2011). Moreover, the porous structure, large surface area, and negative surface charge of biochar (Bird et al., 2008; Cheng et al., 2008; Downie et al., 2009; Novak et al., 2009) can increase the soil's cation exchange capacity and facilitate the retention of nutrients (Liang et al., 2006; Major et al., 2011).
In summary, biochar can promote plant growth by changing the soil physical and chemical properties, such as particle structure, pH value, water-holding capacity, and thermal dynamics, and can improve the soil nutrient environment by reducing leaching losses, all of which lead to the increase in plant biomass. Gautam and Greenway (2014) and Payne et al. (2018) argued that plants with higher growth rates, extensive root systems, and high plant biomass could remove more nutrients, thus validating the notion that biochar can enhance plant N absorption by increasing plant biomass.
Biochar amendment changes the soil microbial structure and proportion related to the N cycle
Biochar amendment can obviously influence soil microorganisms (Tian et al., 2019). The results of the microbial community structure analysis revealed that the microbial species in the natural lakebank and biochar-based lakebank were similar, but the microbial structure and proportion presented obvious differences. The abundance of microorganisms involved in nitrification and denitrification in soil, such as Nitrospira, Nitrosomonadaceae, and Methylophilaceae, improved with the addition of biochar, while the abundance of some heterotrophic bacteria such as Actinobacteria, Sphingomonadales, Mesorhizobium, and Nocardioides was lower in the biochar-based lakebank, when compared with that in the natural lakebank.
The addition of biochar altered the carbon source utilization patterns of microorganisms, possibly via changes in the microbial population (Pietikäinen et al., 2000). It has been shown that microbial utilization of amino acids and amines strongly increased after biochar addition owing to an increase in the activity of proteolytic (
The large surface area and micropores of biochar might immobilize more bioavailable carbon in soil (Zhang and Sun, 2014), which might be difficult for microorganisms to access. In addition, biochar itself is recalcitrant for microbial utilization. As a result, the abundance of some heterotrophic bacteria, such as Actinobacteria, which play a vital role in organic matter degradation, was lower (Antunes et al., 2016; Varma et al., 2018; Meng et al., 2019; Sun et al., 2019). In summary, biochar amendment alters microbial structure and species proportion mainly by changing the soil physical/chemical properties and substrate utilization pattern of microorganisms.
At the genus level, the abundance of Nitrospira, Nitrosomonadaceae, and Methylophilaceae was higher in the biochar-based lakebank, when compared with that in the natural lakebank. Nitrospira is a nitrifying bacterium with nitrification ability; Nitrosomonas (Nitrosomonadaceae) can oxidize ammonia to nitrous acid, and Methylophilaceae can perform aerobic denitrification of nitrate and nitrite (Beck et al., 2013). Thus, a higher proportion of these microorganisms in the biochar-based lakebank resulted in better N removal efficiency. However, the abundance of Mesorhizobium was lower in the biochar-based lakebank, when compared with that in the natural lakebank. Mesorhizobium is a Rhizobium, which can help the host plant to absorb free N in the atmosphere (Van et al., 1998; Sy et al., 2001). The lower abundance of Mesorhizobium in the biochar-based lakebank indicated lower N absorption by plants from the atmosphere, which may be one of the reasons for lower TN contents in plants in the biochar-based lakebank. However, the total amount of TN absorbed by plants was higher, which could be owing to the higher N absorption from the biochar-based lakebank system itself rather than from the atmosphere. Besides, the abundance of other heterotrophic bacteria such as Actinobacteria, Sphingomonadales, and Nocardioides was lower in the biochar-based lakebank, which is conducive to SOM preservation. SOM is an important source of plant nutrition and can also improve soil physical and chemical properties, maintain soil fertility, and promote plant growth (Mullen et al., 1999; Saffih-Hdadi and Mary, 2008). As a result, the biochar-based lakebank presented higher plant biomass and better N removal via plant absorption.
In summary, the biochar-based lakebank exhibited a higher abundance of nitrifying and denitrifying bacteria, which improved the N removal effect. The low abundance of Mesorhizobium reduced the amount of free N from the atmosphere in plants, leading to a decrease in TN content in plants. The total amount of N in plants was higher in the biochar-based lakebank due to the lower abundance of heterotrophic bacteria related to organic matter degradation, leading to SOM preservation, which resulted in higher plant biomass and improved N absorption by plants.
Conclusions
The following conclusions can be derived based on the results obtained in the present study:
Biochar amendment significantly enhanced (p < 0.05) N removal in lakebanks, especially in the plant growth season. Biochar amendment improved the N interception/removal capacity, mainly by increasing the effect of plant absorption and microbial utilization. Biochar amendment increased plant biomass by changing the soil physical/chemical properties and enhanced the soil nutrient environment to improve the total amount of N absorbed by plants, instead of increasing the N content in plants. Biochar amendment improved the abundance of nitrobacteria, nitrifying bacteria, and aerobic denitrifying bacteria related to the N cycle, which promoted microbial N removal, and decreased the abundance of some heterotrophic bacteria, which promoted plant growth and increased the N absorption capacity of plants. Production of biochar from lakebank plant litter and its subsequent amendment to lakebank soil not only allow recycling of plant litter, but also further enhance the ecological function of lakebanks.
Footnotes
Acknowledgments
We thank the International Science Editing for editing the article. We thank the authorities of the College of Environmental Science and Engineering, Tongji University, for providing necessary facilities during the course of our studies. We thank the authorities of School of Life Sciences, Shanghai University, for providing necessary facilities during the course of our studies. We also thank the authorities of Shanghai Academy of Environmental Sciences, Shanghai, China, for providing necessary facilities during the course of our studies.
Authors' Contributions
L.C.: Writing—original draft, methodology, software, validation, and data curation.
L.W.: Conceptualization, supervision, and funding acquisition.
X.L.: Methodology and investigation.
Q.Y.: Literature search.
D.L.: Writing—review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was financially supported by the National Natural Science Foundation of China (No. 21876127) and the China National Key Research and Development Program (No. 2017YFC0506004).