
Abstract
Disinfectant decay by biofilms in distribution networks during stagnation can allow opportunistic pathogens' transmission and thus compromise drinking water safety. Applying phosphate-based corrosion inhibitors to the system can exacerbate disinfectant decay by providing nutrients to biofilms growing inside premise plumbings. In this study, we evaluate the impacts of corrosion inhibitors on biofilms' structural and chemical properties that form in premise plumbing, and the resulting implications for disinfectant decay. Two commonly used phosphate-based (phosphate blends and phosphate) corrosion inhibitors were added separately to simulated drinking water for biofilm development over 1 to 2 years. Optical coherence tomography (OCT) imaging showed that the studied biofilms' thickness, porosity, and porous structure did not change after exposure to free chlorine for 24 h or monochloramine for 120 h. Compared with groundwater biofilms, phosphate-based biofilms had the highest overall porosity due to their many connecting channels. The phosphate-based biofilms consumed free chlorine or monochloramine at a faster rate than groundwater biofilms. Experimental results showed that phosphate-based biofilms consumed more monochloramine after 96 h of contact than other biofilms. A separate set of experiments involving disinfectant decay with suspended biomass material, together with the OCT results, provided parameters for a simplified quasi-first-order reaction–diffusion model so that predictive modeling of decay in biofilms under stagnation conditions could be attempted without parameter fitting. The biofilm modeling results provided a close estimate for free chlorine decay while underestimating monochloramine decay. In agreement with the experimental results, the model results indicate that the phosphate-based biofilms led to slightly faster free chlorine consumption and monochloramine consumption than groundwater biofilms and indicate that diffusion limitation imposed by biofilm pore structure on disinfectant decay is important. The study results suggest that using phosphate-based corrosion inhibitors may lead to a rapid depletion of residual disinfectant during stagnation in the presence of biofilms.
Introduction
Biofilms are ubiquitous in drinking water distribution systems (DWDSs) (Bloetscher et al., 2010). Corrosion inhibitors, commonly added to decrease lead and copper corrosion products in drinking water, are usually phosphate based (about 94% of utilities in the United States) and can provide nutrients to microorganisms grown as biofilms in drinking water systems (Kogo et al., 2017). Residual disinfectants (free chlorine and monochloramine) are required by the USEPA to prevent opportunistic pathogen propagation in the drinking water system (Prest et al., 2016; Rennecker et al., 2001; Wang et al., 2012). However, residual disinfectants can be depleted by organic matter, biofilms, and pipe material in both large- and small-scale distribution systems, further exacerbated by long stagnation and low flow rate devices implemented for water conservation practice in premise plumbing (Buse et al., 2019b; Lautenschlager et al., 2010; Ley et al., 2020; Li et al., 2019; Ling et al., 2018; Tolofari et al., 2021; Wang et al., 2012).
Opportunistic pathogens can find shelter in biofilms and can pose health risk to water users when the disinfectant concentrations are inadequate (Abdel-Nour et al., 2013; Cooper and Hanlon, 2010; Declerck, 2010; Declerck et al., 2009; Farhat et al., 2012). The disinfectant decay depends on multiple factors, including pH and natural organic matter in the water, scaling extent on the pipe surface, and the pipe material in the DWDS (Vikesland et al., 2001b; Wang et al., 2013a; Xue and Seo, 2013). Disinfectant reactivity has been measured with single-cell cultures, suspended particles in drinking water sources, single-species biofilm clusters, and biofilms harvested from simulated drinking water sources (Boccelli et al., 2003; Du et al., 2022; Lee et al., 2011; Zhao et al., 2018). Chlorine decay in large-scale DWDSs has been modeled based on these disinfectant reactivities. However, estimating or optimizing a disinfectant dose in biofilm-covered drinking water systems can be complicated because biofilms are multispecies and develop under various water sources and hydraulic regimes (Abokifa et al., 2016a; Abokifa et al., 2016b; Douterelo et al., 2016; Fish et al., 2017; Fish et al., 2015; Zhu et al., 2020).
Kinetics of free chlorine decay is commonly modeled using first-order kinetics, in large scale distribution (e.g., Monteiro et al., 2014) and premise plumbing networks (Zheng et al., 2015), and while chloramine decay is sometimes approximated with as first-order kinetic with additional autodecay, more rigorous simulation requires significantly more complex approaches (Hossain et al., 2022). In either case, the presence of biofilm has been indicated as an additional factor impacting decay kinetics and disinfection progress (e.g., Buse et al., 2019a; Huang et al., 2020a), requiring separate consideration of kinetics associated with the biofilm (Hossein et al., 2022; Monteiro et al., 2014). Water-filled pores and channels in the biofilm matrix have been shown to accommodate the aqueous-phase transport of nutrients, oxygens, and disinfectants within single-species laboratory-grown biofilms (Lee et al., 2018; Quan et al., 2022). The penetration of free chlorine into biofilms by diffusion was impacted by its strong oxidant nature that leads to a fast reaction with biofilm material, while monochloramine can penetrate deeper into the biofilm matrix due to its lower reactivity (Lee et al., 2018; Lee et al., 2011).
These observations suggest that biofilm structure may play a significant role in disinfectant transport and effectiveness. However, effective diffusion coefficients of mature biofilms depend on the structural heterogeneity of biofilm, which can be altered by phosphorus addition, a common ingredient in corrosion inhibitors (Davit et al., 2013; Fang et al., 2009; Quan et al., 2022). Thus, the effect of biofilms' structure and chemical composition developed under these common corrosion inhibitors on disinfectant decay should be studied.
To fill the knowledge gaps on the role of corrosion inhibitors on biofilm characteristics and disinfectant consumption by these biofilms in the premise plumbing, we (1) characterized the structure and chemical composition of biofilms grown with or without phosphate-based corrosion inhibitors using optical coherence tomography (OCT) and Fourier-transform infrared spectroscopy (FTIR); (2) characterized the pore structure of the three biofilms by pore network analysis; (3) measured the reactivity of two disinfectants with biomass harvested from three biofilms; and (4) determined the disinfectant decay after stagnation contact with three intact biofilms, and modeled decay as a reaction–diffusion process within the biofilm.
Methods
A flow diagram of this study is shown in Fig. 1.
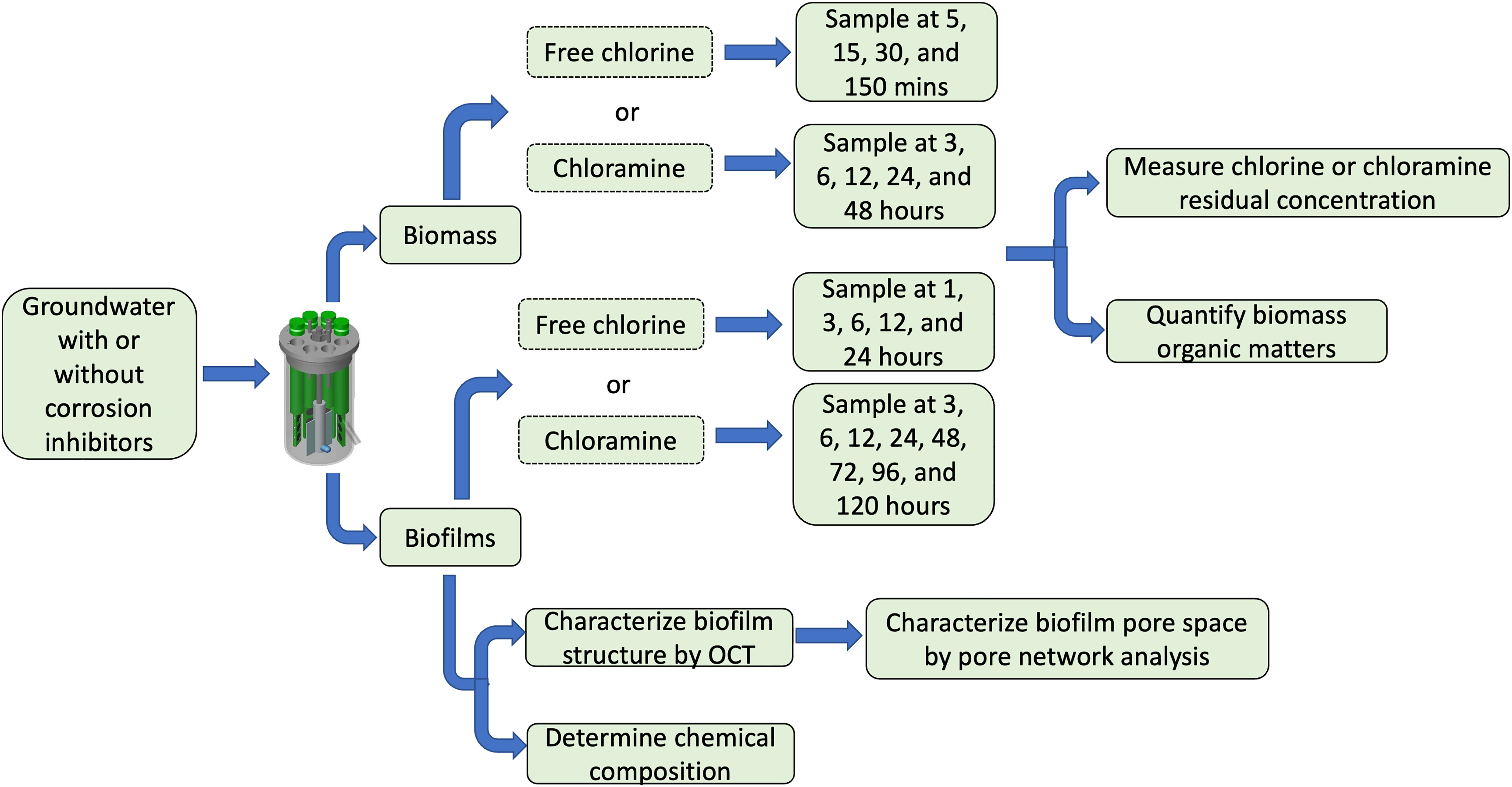
Flow diagram for experiments to characterize biofilm structure and disinfectant consumption.
Biofilm growth conditions
Biofilms were grown on coupon-holding polypropylene (PP) rods and polyvinyl chloride (PVC) coupons, materials commonly found in premise plumbing systems. These rods and coupons were inserted into three separate CDC reactors® (BioSurface Technologies Corporation) fed by groundwater with or without corrosion inhibitors. Groundwater, the drinking water source in Champaign, IL, was passed through a green-sand filter, which was similar to the filtration process used in local drinking water treatment plant. This filtration removed the precipitates formed by calcium and magnesium when groundwater is exposed to air. The original groundwater did not contain disinfectants. Two corrosion inhibitors were used in this study due to their popularity in utilities: (1) phosphate (2 mg/L as PO4) and (2) phosphate blends (2.4 mg/L as PO4). Groundwater without corrosion inhibitors was fed to a separate reactor as a control condition. The operation conditions of biofilm reactors are described in the Supplementary Information SI.
Biofilm porous structure characterization
Structures of the three studied biofilms were characterized by cross-sectional scans with a dimension of 3.125 × 4.18 × 4 mm using OCT. OCT image allowed a nondestructive study of biofilm structure based on the signal intensity distributions in the OCT images (Hou et al., 2019). The axial resolution of the OCT was around 8 μm, and the lateral resolution was around 20 μm. One to three locations were used randomly from one to three biofilm-covered coupon from each reactor. The OCT images were rotated to correct for taking images at an angle and modified to reduce reflection caused by water using ImageJ Fiji v 2.0 (Schindelin et al., 2012) and Avizo (Thermo Fisher Scientific). About 30–180 images from each image stack were selected for analysis because of the image quality, as described in a previous study (Huang et al., 2020b). The overall biofilm thickness was determined using a MATLAB code described previously (Derlon et al., 2012; Shen et al., 2015). The physical thickness of biofilm was determined by dividing the optical thickness by the corresponding refractive index from 1.33 to 1.38. The average thickness of each biofilm image stack was calculated as described in previous studies (Huang et al., 2020b).
The upper and lower halves of the biofilm OCT image stacks were analyzed separately after dividing the biofilm at half of the average thickness. The porosities and pore network were characterized in the top and bottom halves of the biofilms using the image analysis framework described in a previous study (Huang et al., 2020b).
Chemical composition characterization by FTIR
We used FTIR measurements to determine the chemical composition of biofilms developed with or without phosphate-based corrosion inhibitors based on the interactions between the electromagnetic radiations and vibrations of chemical bonds (Fadlelmoula et al., 2022). Biomass was scraped from the holding rods into clean Petri dishes and dried overnight at 4°C. Two biomass samples were subjected to FTIR scans, in which 15 scans were obtained for each biofilm type. The absorbance of the infrared radiation by the dried biomass over the wavenumber from 400 to 4,000 cm−1 was measured by Fourier transform infrared spectroscopy equipped with an attenuated total reflection element (ATR-FTIR). The wavenumbers associated with the absorbance peaks were compared with the IR spectrums reported in previous studies (Holman et al., 2009; Schmitt and Flemming, 1998). The results of the IR spectrum are shown in the Supplementary Information SI.
Disinfectant decay in contact with suspended biomass
Our first decay experiments involve exposing biomass, scraped from the coupon biofilm and sonicated to form a suspension, to free chlorine or monochloramine. The free chlorine and monochloramine stocks were prepared as described in the Supplementary Information SI. A 2 mM bicarbonate buffer (pH: 7.8–8.1) was prepared by dissolving sodium bicarbonate (Fisher Scientific) in Milli-Q water to simulate the drinking water alkalinity condition (200 mg/L as CaCO3). The disinfectant concentration was measured using a standard curve method, as described in the Supplementary Information SI.
Biomass was scraped from the rods in the CDC reactors using a sterilized spatula and dispensed in 50-mL glass vials containing 15 mL bicarbonate buffer. Then, these biofilms were disrupted by 1 min of vortex followed by 20 min of sonication. Free chlorine or monochloramine stock was added to the suspended biomass to reach a final concentration of 4.0–4.1 mg/L as Cl2. The vials were wrapped in foil and placed in a shaker for a designated reaction time, after which a 2.5 mL aliquot was filtered through a 0.22 μm PES syringe filter (Millipore), and the free chlorine or chloramine concentration was measured using the DPD standard method (4500-Cl CHLORINE). The reaction times were 5, 15, 30, 150 min for free chlorine and 3, 6, 12, 24, and 48 h for monochloramine. These reaction times were selected based on the reaction time for free chlorine or monochloramine to reach the detection limit of 0.02 mg/L as Cl2 with biomass in preliminary experiments.
To the remaining suspended biomass in buffer, 2.7 g/L sodium thiosulfate was added to quench the residual disinfectant, and then 10 N NaOH at a final concentration of 0.25 M (pH at 11) was added to digest the sample. The sample was then placed on a shaker table at 250 rpm for 3 h in the dark. The volume of sodium thiosulfate needed to neutralize the residual disinfectants was checked with preliminary experiments. These samples were filtered through a 0.22 μm PES filter and the pH was lowered to 2 by adding concentrated HCl. The total organic matter (TOC) concentrations of digested samples were determined by a Shimadzu TOC-VCPH analyzer. Experiments were conducted in triplicates in free chlorine experiments and duplicates in monochloramine experiments because of sample limitation.
Disinfectant decay in contact with intact biofilms
Two biofilm-covered PP rods with three PVC coupons were removed from the CDC reactor and immersed in glass flasks filled with 250 mL bicarbonate buffer. Freshly prepared disinfectant stock (free chlorine or chloramine) was added into the bicarbonate buffer to reach a final concentration of 4 mg/L as Cl2. The disinfectant concentration was measured using the DPD method and the corresponding standard curves, as described in Disinfectant decay in contact with suspended biomass section. As aforementioned sample collection, a 2.5 mL sample was taken from the flask and filtered by a 0.22 μm PES syringe filter (Millipore) to measure disinfectant concentrations after the corresponding contact time. The remaining samples after determination of the disinfectant concentrations were quenched with 2.7 g/L sodium thiosulfate. The contact times for biofilms with free chlorine were 1, 3, 6, 12, and 24 h, and the contact times with chloramine were 3, 6, 12, 24, 48, 72, 96, and 120 h due to the low reactivity of chloramine. After these times, the biofilms were scraped from the PVC coupons on the rods and resuspended into a 15 mL bicarbonate buffer.
As described above, the biofilms were disrupted by vortex and sonication, followed by digestion with NaOH addition in the shaker. TOC concentrations were determined from the digested samples. The total biomass concentration was computed by multiplying the TOC concentration obtained from the scraped biofilms by the ratio of total biofilm area to the scraped biofilm area.
Mathematical modeling of reaction with biomass and diffusion–reaction with biofilm
Disinfectant decay in the presence of suspended biomass
Under conditions of suspended biomass, we ignore diffusion limitations and treat the time-dependent consumption of free chlorine or chloramine as a quasi-first-order kinetic,
where
Disinfectant diffusion and decay within intact biofilms
To simultaneously quantify the time-dependent impacts of both disinfectant diffusion within and reaction with porous biofilm, we applied a conventional transient reaction–diffusion model,
with initial and boundary conditions
respectively. This model treats the biofilm as a porous slab of finite thickness admitting one-dimensional Fickian diffusion of disinfectant concentration C. The biofilm has thickness l, total areal extent A, porosity n, diffusion coefficient
At the surface of the biofilm x = 0, the solution in the biofilm is in contact with the bulk solution that has a concentration,
with initial condition
Equations (2)–(6) were solved numerically using a conventional explicit-in-time finite-difference scheme with central differencing used for the diffusion term. This approach is simple to implement but requires honoring the von Neumann condition for stability of the diffusive flux calculation that is implemented here as
Application of the mathematical models
Predictive modeling requires independent determination of parameters. The determination of the decay and diffusion parameters is detailed in the Supplementary Information SI. The remaining parameters needed for Equations (2)–(6) were directly measured and include biofilm area, thickness (mean values from Fig. 2a), and porosity (mean values from Fig. 2b). The TOC concentration (B) for the intact biofilms was measured separately. For free chlorine cases, this procedure was repeated for all three biofilm types and completed the independent determination of values of parameters needed for the use of Equations (2)–(6) to simulate the decay data in the intact biofilm cases. The reaction rate in the case of monochloramine includes auto-decay at first-order, in addition to the kinetic model described above and in the Supplementary Information SI. The mechanisms of monochloramine auto decomposition are relatively complex and can depend on various factors such as temperature, pH, the ratio of ammonia, chlorine, and intermediates (Hossain et al., 2022).
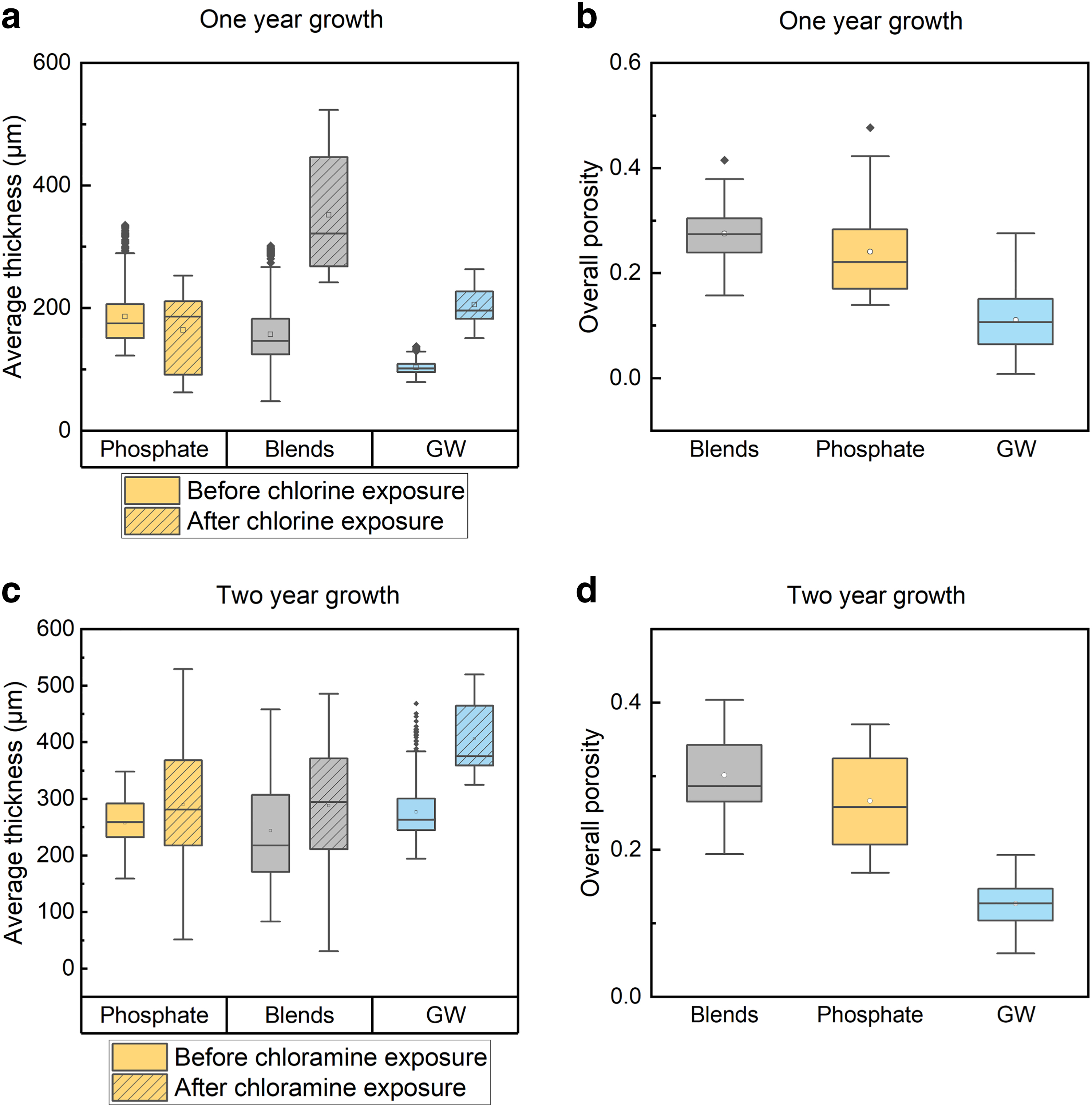
Biofilm mean thickness
In this study, we estimated the overall auto-decay rate of monochloramine to be
Model Parameters Obtained from the Disinfectant Decay Experiments and Biofilm Characterization in This Study
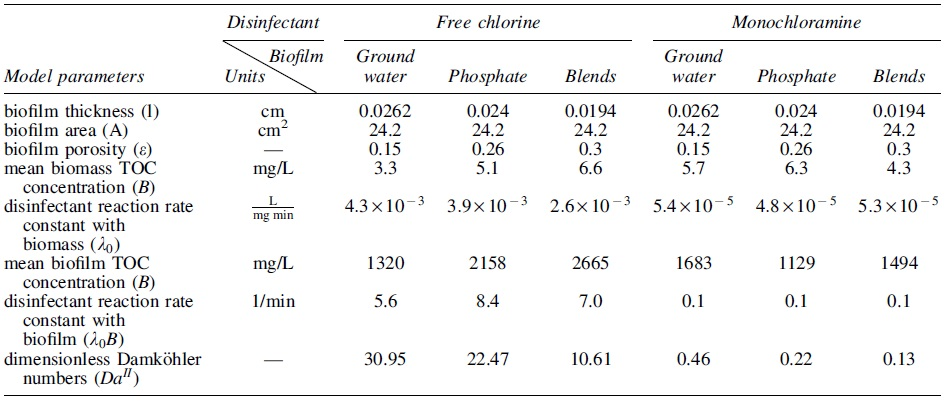
Also shown are dimensionless Damköhler numbers (“
Statistical analysis
Basic statistical analysis and hypothesis tests are described in the Supplementary Information SI.
Results
Biofilm thicknesses, porosities, and their variation with treatments
First, we compared the thickness for 1- and 2-year biofilms before disinfectant exposure (Fig. 2). For a given biofilm type developed with phosphate corrosion inhibitors, 1- and 2-year biofilms have similar thicknesses (KS test, p > 0.05). Among the three 1-year biofilms, phosphate biofilms were the thickest with a median of 175 μm (95% confidence interval [CI]: 135–293 μm) followed by phosphate-blend biofilms with a median of 147 μm (95% CI: 84–246 μm). The 1-year groundwater biofilms were the thinnest (median: 102 μm, 95% CI: 87–126 μm). However, among the 2-year biofilms, groundwater biofilms were thickest (median: 263 μm, 95% CI: 221–374 μm). The 2-year phosphate biofilms were the second thickest with a median of 259 μm (95% CI: 178–319 μm) and followed by the 2-year phosphate-blend biofilm with a median of 218 μm (120 to 433 μm, 2-year), (KS test, p < 0.05). We found similar and slightly increased thicknesses for biofilms grown with phosphate and phosphate blends for 1 and 2 years. The thickness of biofilms grown without corrosion inhibitors more than doubled during the second year.
This observation suggests that phosphate allowed the biofilms to grow to a steady state faster. Thus, the time required for biofilm growth to reach the steady state may depend on biofilm age, feed water constituent, and hydrodynamic conditions, which was not studied in this work. (Mathieu et al., 2014; Shen et al., 2015; Simunič et al., 2020).
After 1 year of development (Fig. 2b), the overall porosities of phosphate blends (0.27, 95% CI: 0.19–0.36) were the greatest, followed by those of phosphate biofilms (0.22, 95% CI: 0.15–0.39) and groundwater biofilms (0.11, 95% CI: 0.015–0.25) (KS test, p < 0.05). After 2 years of development (Fig. 2d), the medians of overall porosity of phosphate blends, phosphate, and groundwater biofilms was 0.29 (95% CI: 0.21–0.40), 0.26 (95% CI: 0.17–0.37), and 0.13 (95% CI: 0.08–0.17), respectively. No significant change in the overall porosities between 1- and 2-year growth were observed in any of the biofilms (p > 0.05). The overall porosities in biofilms fed by phosphate and phosphate blends were similar (p > 0.05) and significantly greater than groundwater biofilms (p < 0.05, KS test).
For the 1-year biofilms (Fig. 2a), after 24 h of free chlorine exposure, the average thickness of phosphate biofilm reduced slightly (KS test, p < 0.05), while the average thickness of phosphate blends and groundwater biofilms increased. A similar trend was observed in 2-year-old biofilms exposed to monochloramine. This observation showed that short-term exposure (e.g., 24 h) to free chlorine and monochloramine did not remove the biofilm. For biofilms fed by the same water source, the differences in biofilm thickness can be from 48 to 469 μm. Depending on the scanned locations, the average biofilm thickness can fall in a wide range. The differences in the biofilm thickness before and after disinfectant exposure can be attributed to the multispecies biofilm heterogeneity. By comparing the upper and lower halves of the biofilm porosities separately (Supplementary Fig. 2S), no significant difference was found in the top or bottom section of phosphate-blend biofilm after free chlorine exposure (t test, p > 0.05). The porosities in the bottom section of the phosphate biofilms were reduced after free chlorine exposure (t test, p < 0.05), while the porosities in the top section remained the same (t test, p > 0.05)
However, the porosities in either top or bottom section of groundwater biofilms increased after free chlorine exposure (t test, p < 0.05). Similar trends of increasing porosities in the bottom or top section of groundwater biofilms were observed after monochloramine exposure (t test, p < 0.05). In either the top or bottom section of the 2-year-old biofilms, groundwater biofilms were the least porous than phosphate-based biofilms before and after monochloramine exposure (t test, p < 0.05). No significant difference in porosities was found in 2-year-old biofilms between the two sections (p > 0.05). We expected deeper diffusive transport into the biofilm compared with that of free chlorine because of monochloramine's slower reactivity. The porous top and bottom section in the 2-year-old phosphate and phosphate-blend biofilms may allow the slowly reacting monochloramine transport further into the biofilm matrix. We tested this hypothesis with experimental results presented below.
Pore structure analysis in biofilms grown with or without corrosion inhibitors
The pore structure of the biofilms was modeled as a newtork of spherical pores connected by cylindrical throats. The number of pores and throats in each biofilm used in the free chlorine and monochloramine scenarios were normalized by the total biofilm volume, including biomass and pore space. The pore network analysis showed that phosphate blends and phosphate biofilms had more pores per biofilm volume and more connecting throats per biofilm volume than groundwater biofilms (KS test, p < 0.05) (Fig. 3a). After one-year growth (Fig. 3a), the highest normalized number of pores was found in phosphate and phosphate-blend biofilms, followed by groundwater biofilms (KS test, p < 0.05 each). Groundwater biofilms had the lowest normalized number of throats (KS test, p < 0.05 each), while the other two biofilms had a similar number of throats after normalization (KS test, p > 0.05 each). The median normalized number of connecting throats in groundwater biofilms was 20 times less than that in phosphate-blend biofilms, and 18 times less than that in phosphate biofilms. Among the 2-year-old biofilms (Fig. 3b), phosphate blends and phosphate biofilms had more normalized throats than groundwater biofilms (KS test, p < 0.05).

Pore (solid) and throat (dashed) number normalized by biofilm volume after
However, no significant difference between the normalized number of pores was found in phosphate, phosphate blends, and groundwater biofilms. Overall, the great abundance of connecting throats in phosphate and phosphate-blend biofilms contributed to the greater bulk porosity of these biofilms compared with groundwater biofilms.
The biofilm structure can be influenced by microbial structure, influent water characteristics, and hydrodynamic conditions (Liu et al., 2016). In this study, biofilms developed under the influence of phosphate-based corrosion inhibitors were more porous with more connecting channels than those developed without phosphate corrosion inhibitors. A similar observation was reported from biofilms developed in groundwater with or without polyphosphate additives (Shen et al., 2018). Previous studies showed that the addition of phosphate induced changes in the microbial community and EPS production (Liu et al., 2016). These changes can play an important role in the reactivity of biofilms with chlorine and monochloramine and the disinfectant byproducts' formation (Lemus Pérez and Rodríguez Susa, 2017; Wang et al., 2013b). In the present study, larger connecting channels and pores in the pore structure found in biofilms developed under phosphate-based corrosion inhibitors may aid in the mass transport process of chlorine and chloramine into the biofilm matrix. A similar observation was reported as greater biofilm removal by chlorine and monochloramine was found after phosphorus addition (Fang et al., 2010).
Experimental results: free chlorine and chloramine decay in biofilms grown with or without corrosion inhibitors
The experimentally determined free chlorine or monochloramine decays were normalized by the biomass TOC concentrations (Fig. 4). We normalized the disinfectant concentrations by the biomass TOC concentrations because the disinfectant decay experiments with biomass were carried out independently at each sampling time. Free chlorine was rapidly consumed by the three types of biomasses within 2 h of contact. Among the three biomass, the groundwater biomass consumed free chlorine the least, regardless of sampling time (p < 0.05). On the other hand, phosphate-based biomass consumed the most free chlorine at all sampling times (p < 0.05). However, we did not observe one type of biomass consistently consuming more monochloramine than other types. Among the three biomass, greater TOC-normalized monochloramine concentrations were found in phosphate-blend biomass after 3, 6, 24, and 48 h of contact (p < 0.05). Groundwater biomass consumed the least monochloramine after 12 h of contact (p < 0.05).
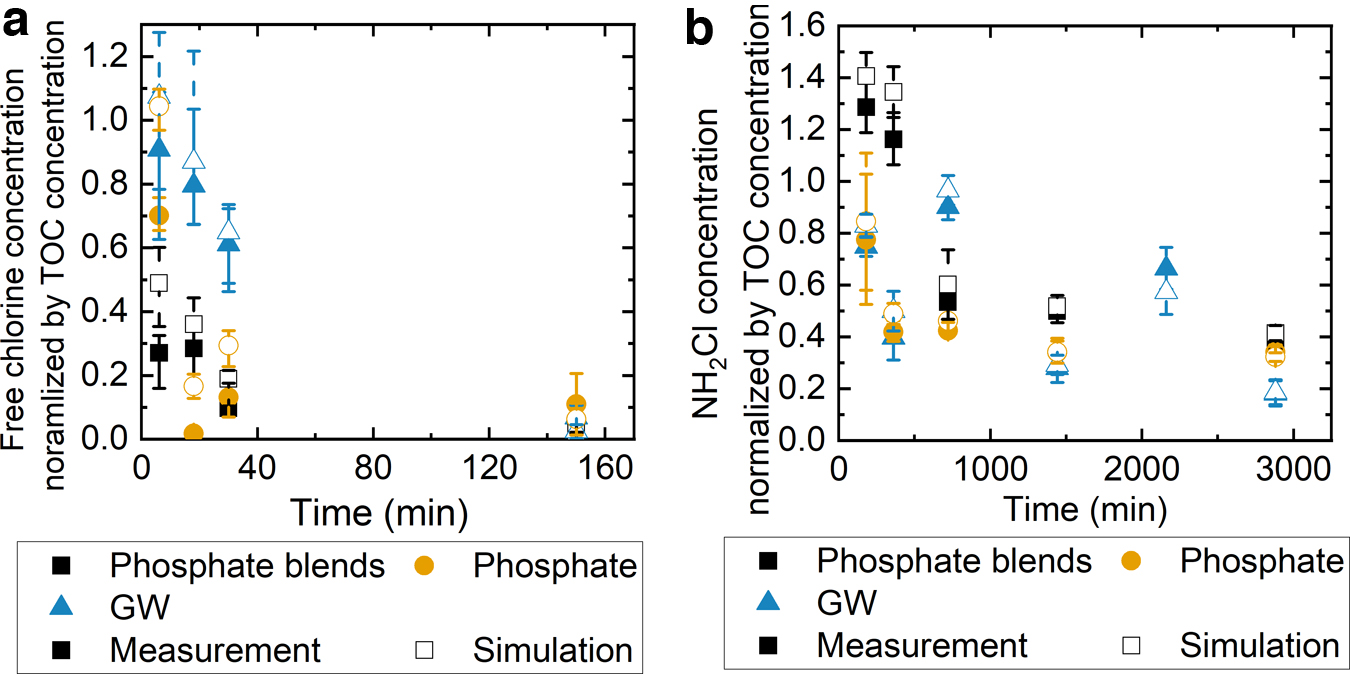
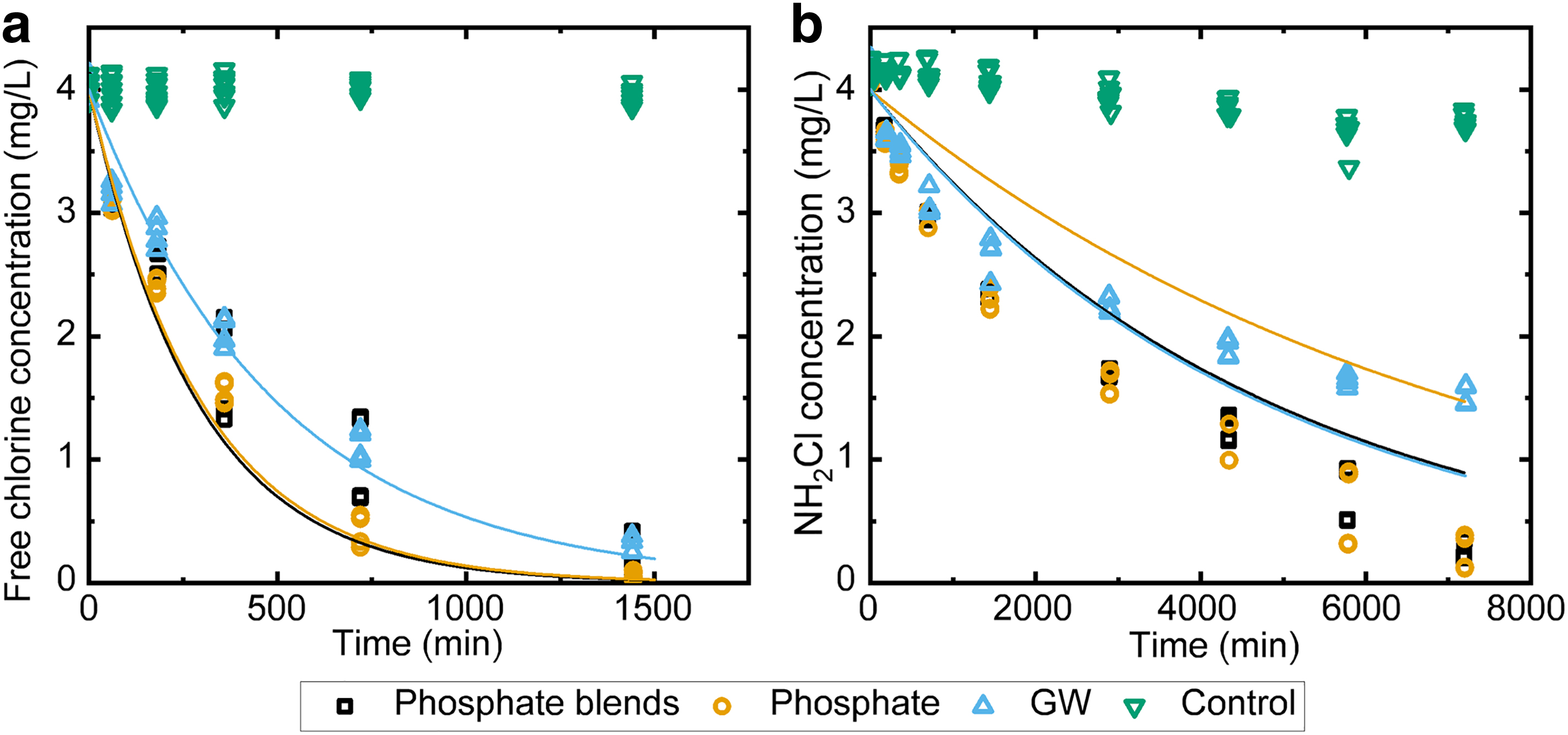
The experimentally measured (solid) and simulated (open) free chlorine
To determine the effect of the biofilm structure on the disinfectant reactive transport into the biofilms, we exposed the intact biofilms to either free chlorine or monochloramine. The disinfectant decays obtained with three biofilms are shown in Fig. 5a, b. Free chlorine was continuously and completely consumed by all three biofilms within 24 h of reaction, significantly faster than a scenario without biofilm (p < 0.05). In the first hour of contact, no significant difference in free chlorine decay was found among the three biofilms. After 3 h of contact, the phosphate biofilms consumed the most free chlorine, while the groundwater biofilms consumed the least free chlorine among the three biofilms (p < 0.05). In the first 3 h of contact, the three biofilms consumed monochloramine at a similar rate (p > 0.05). Monochloramine was consumed after 96 h of contact with biofilms grown with phosphate-based additives. No significant difference was observed in monochloramine concentrations between phosphate and phosphate-blend biofilms after 3 h of exposure (p > 0.05). This can be contributed by the similar general porosities in phosphate and phosphate-blend biofilms.
Free chlorine
However, after about 60% of monochloramine was consumed by the groundwater biofilms, monochloramine decay continued at a much slower rate (after 96 h). This reduced consumption of monochloramine may be due to the presence of fewer connecting throats (Fig. 3) and reduced porosities in top in the groundwater biofilms (Supplemenary Fig. 2S) because these throats may promote the transport of monochloramine into the biofilm matrix and lead to complete consumption in phosphate-based biofilms. Compared with monochloramine, free chlorine decay rate with biomass was much faster, with congruently higher Damköhler numbers indicating the larger role of diffusion in limiting the reaction. The variation in the free chlorine decay rates with biomass was large but the range reflected does not come close to the slowness of the diffusion-controlled reaction rates in the biofilm. On the contrary, the reaction kinetics evidently dominate the rate of monochloramine decay, with low Damköhler numbers (i.e., less diffusion limitation). In this case, the structure of the biomass still plays a significant role in the decay reactions.
In any case the Damköhler numbers indicate that diffusion limitation plays a larger role in the free chlorine case than it does in the monochloramine case.
Theoretical framework: free chlorine and monochloramine decay with biofilms
We first determined the specific reaction rate constants (
Slightly greater average monochloramine concentrations were simulated than measurements with phosphate blends and groundwater biomass after 3 h of reaction and two phosphate-containing biomass after 6 h of reaction (t test, p < 0.05). As shown in Table 1, the free chlorine rate constants were the greatest with biomass grown without corrosion inhibitors compared with those grown with phosphate-based corrosion inhibitors. Similar trends were observed in the monochloramine reactions with suspended biomass.
Turning to the cases involving intact biofilms, numerical solutions to Equations (2)–(6), independently parameterized as described in Experimental results: free chlorine and chloramine decay in biofilms grown with or without corrosion inhibitors section were used to simulate the dynamics of disinfectant decay for free chlorine (solid lines in Fig. 5a) and for monochloramine (solid lines in Fig. 5b), for all three biofilm types. The model underestimated the chlorine consumption within 1 h of contact for all three biofilms (t test, p < 0.05). The differences between the simulations and measurements of free chlorine were not statistically significant at all sampling times thereafter in the case of the groundwater biofilms (t test, p > 0.05). However, the model overestimated the free chlorine consumption at 3 and 12 h of contact and at 3 and 6 h of contact with phosphate blends and phosphate biofilms, respectively (t test, p < 0.05). Other than the aforementioned contact times for phosphate and phosphate-blend biofilms, the simulation agreed with the measurements (t test, p > 0.05).
On the contrary, the model did not agree with the simulations at all sampling time and biofilm growing conditions with monochloramine (t test, p < 0.05). With approximately constant reactive TOC the decay reaction is treated in both free chlorine and monochloramine cases as a first-order reaction with effective rate constant given by the product of the specific rate constant (
While differences in overall free chlorine decay patterns among the biofilm types are subtle, the variation of predicted concentrations between biofilm types is larger than the errors between data and simulation in any given case. The prediction of the dynamics in the monochloramine case is much less successful. Simple inspection of the monochloramine decay in the case of suspended biomass strongly suggested that the reaction was not quasi-first-order (Equations (1) and (2)) because the trends in disinfectant concentrations are not exponential. Speciation reactions may also complicate the reaction and diffusive transport dynamics (Vikesland et al., 2001a). Second, the reaction–diffusion model assumes homogeneous porosity, whereas the pore networks exhibited heterogeneity. It is well known that when reactions are linear (as in the free chlorine case), such small-scale heterogeneity does not generally invalidate reactive transport modeling at larger scales under the assumption of effectively homogeneous media; however, this is not demonstrated for nonlinear reactions such as may occur with monochloramine (Murphy and Ginn, 2000; Wood and Whitaker, 1998).
In the case of monochloramine, the model overpredicted decay in the phosphate-based biofilm but underestimated decay in the groundwater biofilm case. This may be associated with our finding of more connected channels in the former than in the latter biofilms. However, without rigorous pore-scale modeling of the reactive transport that is beyond our practical-modeling scope, this suggestion remains purely speculative.
Conclusion
In this study, we found that using phosphate-based corrosion inhibitors led to porous biofilms and complete consumption of free chlorine and monochloramine after 24 and 120 h of stagnation, respectively. After short-term exposure to free chlorine and monochloramine, no significant disturbance in biofilm thickness was observed. The results presented in this study suggested that short-term disinfectant exposure did not remove biofilm grown inside the PVC pipes, especially when the drinking water was under phosphate-based corrosion inhibition treatment. Because corrosion inhibitors are commonly used to prevent elevated levels of metal leaching from the pipes, especially in aged DWDS, consumption of disinfectant is unavoidable in the premise plumbing.
Footnotes
Acknowledgments
The authors acknowledge partial support from NSF grants 1855609, 1855211, and the NIH/NIBIB Center for Label-free Imaging and Multiscale Biophotonics (CLIMB) P41EB031772. They thank Dr. Rosa M. Espinosa Marzal and Dr. Mengwei Han (UIUC) for the FTIR scans, Dr. Nicolas Derlon and Professor Eberhard Morgenroth (Eawag) for the MATLAB code used for OCT image analysis
Authors' Contirbution
C.H.: Investigation, validation, methodology, formal analysis, visualization, and writing—original draft, except numerical modeling section. T.G.: Methodology, formal analysis, visualization, and writing—original draft, review, and edit on numerical modeling section. G.C.C.: Writing—review and edit. F.Z., J.W., and S.A.B.: Methodology, investigation, resources, and visualization on OCT image scans. T.H.N.: Conceptualization, supervisation, resources, funding acquisition, and writing—original draft, review, and edit.
Author Disclosure Statement
The authors declare that they have no actual or potential competing financial interests.
Funding Information
We acknowledge partial support from NSF grants 1855609, 1855211, and the NIH/NIBIB Center for Label-free Imaging and Multiscale Biophotonics (CLIMB) P41EB031772.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.