
Abstract
Objective:
Spiritual well-being (SWB) has been shown to delay the onset of cognitive decline among older adults predisposed to Alzheimer’s disease and related neurodegenerative dementias. It was, however, unknown if SWB is also associated with delay in disease manifestation (“phenoconversion”) in rare, genetic neurodegenerative dementias, such as Huntington’s disease (HD). Thus, we sought to evaluate the association between SWB and phenocovnersion in people at-risk for HD.
Methods:
The “Prospective Huntington At Risk Observation Study” (PHAROS), a large-scale national prospective research study, recruited a substantial cohort of 1001 participants. These participants, who were first-degree relatives of individuals diagnosed with HD and had a 50% chance of carrying the HD genetic mutation, were blinded to their genetic status and underwent repeated clinical assessments every 12 months. The study used Cox proportional models to examine the relationship between SWB and time to phenoconversion while also considering subcomponents of SWB and adjusting for age, sex, and CAG repeat length.
Results:
The study found no significant association between SWB and time to phenoconversion in individuals with the HD gene mutation.
Conclusion:
While existing data suggest that SWB may be an actionable target to improve health-related quality of life in HD and other serious illnesses, there is a lack of evidence supporting the role of SWB in attenuating phenoconversion in people with the HD genetic mutation. Unlike Alzheimer’s, HD may be less responsive to analogous factors in delaying disease onset.
Introduction
High-quality palliative care involves screening for and enhancing purpose in life, a cornerstone of spiritual care and spiritual well-being (SWB). 1 Indeed, having a high purpose in life has also been linked to better health outcomes during various serious illnesses, including some neurodegenerative diseases. In Alzheimer’s disease (AD), for example, individuals who identified as having a high purpose in life had a significantly reduced risk of converting from mild cognitive impairment to AD dementia.2,3 Because AD shares pathophysiological similarities with Huntington’s disease (HD), such as abnormal protein processing, aggregation, and selective neuronal vulnerability, we were interested in evaluating whether patients with a high sense of purpose in life also had a reduced risk of disease onset, especially given the lack of FDA-approved disease-modifying therapies in this population. We assessed the relationship between SWB and disease onset (i.e., phenoconversion), examining both the composite SWB score and its individual components: existential and religious well-being. Existential well-being, which is closely associated with purpose in life, particularly in HD, 4 was a key focus of our analysis. Our hypothesis proposed that higher existential well-being would be significantly associated with a delay in disease onset, as would the broader concepts of spiritual and religious well-being. While specific trinucleotide repeat sequences are nearly 100% penetrant in causing the onset of clinical HD, we theorized that these psychosocial factors might serve as resilience mechanisms, potentially delaying disease manifestation by influencing or supporting adaptive brain networks in at-risk individuals. We used data from the Prospective Huntington At Risk Observational Study (PHAROS), a large, longitudinal, double-blind, prospective observational study, to evaluate our a priori hypotheses.
Methods
Subjects from 43 United States and Canada research sites were enrolled in PHAROS between 1999 and 2004. Participants included 1001 adults aged 26 to 55 who had a 50% risk of having the HD gene mutation. 5 All participants underwent testing for the HD gene mutation, where a blood test was obtained to measure the number of CAG repeats in the huntingtin gene. Those with an expanded CAG repeat length (i.e., more than 36 CAG repeats) have the gene mutation and, therefore, will develop pathological evidence of HD during their lifetime, barring other competing mortality risks. Neither the participant nor the research team were told the results of the research testing. The ethical oversight committees (institutional review boards) at each involved location approved the study’s guidelines and consent frameworks.
Over several years, PHAROS participants were examined annually by a trained rater who collected clinical and biological measurements associated with HD. The interquartile range of follow-up time among all PHAROS participants was 3.85 to 8.16 years. For this analysis, we focus on two key measurements. The first is phenoconversion, defined as the first time a clinician is at least 99% confident that the participant’s motor abnormalities are signs of HD. Not all participants with the HD gene mutation phenoconverted during the study. The second is measures of SWB, which were self-reported using the SWB Scale. 6 This scale consists of 20 statements, and the participant indicates the extent of their agreement or disagreement with each. Ten statements relate to religious well-being, and 10 relate to existential well-being. All statements are scored from 0 to 6 and added up for a total SWB score; the total SWB scores ranged from 0 to 120, where higher scores mean higher SWB. The subtotal for religious and existential well-being statements is called the religious subcomponent and existential subcomponent, respectively; scores for each subcomponent range from 0 to 60.
Like previous PHAROS analyses, 7 we define each participant’s baseline SWB visit as the first visit at which the main predictor of interest (in our case, SWB) was measured. The goal of this analysis is to evaluate the association between time to phenoconversion (i.e., the number of years from baseline SWB visit to phenoconversion) and spiritual well-being (i.e., separately analyzing the total SWB, the religious subcomponent, and the existential subcomponent). The outcome, phenoconversion in years, was right-censored at the participant’s final recorded PHAROS visit if they did not meet the criteria for phenoconversion by then. We evaluated the associations separately before and after adjustment for biological sex, age at baseline SWB visit, and CAG repeat length. The inclusion criteria for the analysis are as follows: participants must (1) have SWB scores measured at least once, (2) have greater than 36 CAG repeats, and (3) not meet the criteria for phenoconversion at their baseline SWB visit. Anyone whose first SWB score was measured at their phenoconversion or censoring date was also excluded, since their contribution to the dataset would only be a single timepoint. We evaluated the associations between time to phenoconversion and each measure of SWB using Cox proportional hazards regression models. The Cox models rely on a proportional hazards assumption, which we verified by checking that the Schoenfeld residuals were independent of time. There was no evidence that this assumption does not hold, so using the Cox model was justified. All analyses were conducted in R version 4.1.3.
Results
Of the 1001 participants enrolled in the study, 476 had SWB measures and had less than 37 CAG repeats and were excluded; 211 had SWB measures and met the additional inclusion criteria (see Fig. 1). Of those 211 participants, 43 phenoconverted during the study time.
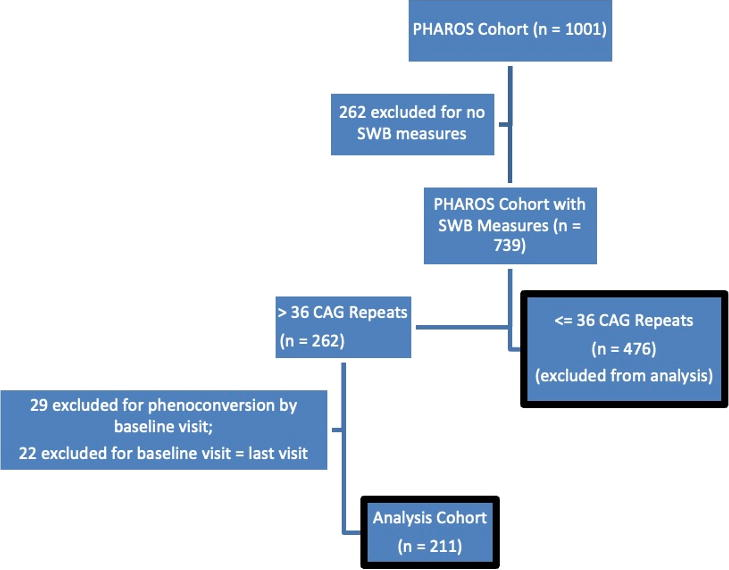
Flow diagram describing the creation of the analysis cohort.
Table 1 presents descriptive statistics for two groups at their baseline SWB visit: (1) the 211 participants that met all inclusion criteria (the analysis cohort) and (2) the group of participants with SWB measures but less than 37 CAG repeats. This table serves a dual purpose in that it describes the analysis cohort and also compares key characteristics from this analysis cohort to the subset PHAROS participants that did not have >36 CAG repeats.
Descriptive Statistics for the PHAROS Cohort at Baseline SWB Visit
This total refers to the sum of Group 1 and Group 2.
This is the p value testing for a difference between Group 1 and Group 2. For continuous variables, we use a two-sample t-test, and for binary variables, we use a chi-square test.
PHAROS, Prospective Huntington At Risk Observation Study; SWB, spiritual well-being.
For the 43 participants who phenoconverted, their average number of years from the baseline SWB visit to phenoconversion was 2.99 (std. dev. = 1.53). There were 25 participants who phenoconverted whose total SWB score was above the median of the analysis cohort (93.0), and 18 participants below the median. For those participants above the median, their average number of years to phenoconversion was 3.07 (std. dev. = 1.38); for those participants below the median, it was 2.88 years (std. dev. = 1.75).
Table 2 displays the unadjusted and adjusted hazard ratios for each of the three models and their 95% confidence intervals. These results show no evidence to suggest that SWB (total, religious, or existential) modifies time to phenoconversion, with and without adjusting for age at baseline SWB visit, CAG repeat length, and biological sex.
Hazard Ratios (HR) and 95% Confidence Intervals (CI) from Cox Proportional Hazards Model
The adjustment set includes biological sex, age, and CAG repeat length.
SWB, spiritual well-being.
Discussion
Our data do not suggest that SWB or its substituent subcomponents of religious or existential well-being play a role in delaying phenoconversion in people with the HD genetic mutation. One of the challenges with our data was the actual censoring rate for phenoconversion: only 20% of the individuals in the study experienced phenoconversion. Still, these results further support that elements of SWB in this population might reflect an inherent trait, given that, as prior research has shown, it does not change over time. 4 However, other research suggests that it remains potentially an amenable and modifiable palliative target for focused therapies, given its association with longitudinal patient-reported outcomes. 4 Indeed, while enhancing existential and religious well-being improves health-related quality of life (HRQoL) modestly in this population, 4 we do not believe our data suggest a compelling influence in delaying disease onset in this population. Finally, our data also suggest that there is indeed a difference between SWB modifying AD phenoconverson, where lifestyle, mood, and other factors are well-known modifiers, 8 and HD, which is monogenetic and may be resistant to influence its phenotypic manifestations through exogenous interventions. Further, while both AD and HD represent progressive neurodegenerative diseases, the studies that used AD had larger sample sizes, and we believe most were unaware of their genetic status, which could potentially motivate the adaptation of healthy lifestyle changes, which might also be a potential hypothesis for these discrepant results. Further, while both AD and HD are progressive neurodegenerative diseases, studies on AD typically involved larger sample sizes. We believe that most participants were unaware of their genetic status, which could have motivated them to adopt healthy lifestyle changes. This, in turn, might offer an alternative hypothesis for the discrepant results observed. Another limitation of our study pertains to the definition of phenoconversion in the HD population. Phenoconversion refers to the appearance of even the most subtle signs of cardinal motor features. However, by the time phenoconversion occurs, around 80% of people with HD already meet clinical criteria for mild cognitive impairment (MCI). 9 Some researchers have called for a revision in how manifest HD is, therefore, defined. A promising area for future research, which would align more closely with studies in the AD population, is to examine how SWB and its substituents might associate with the transition from normal cognition to nonamnestic MCI in HD. Since HD is a subcortical neurodegenerative disease affecting attention, executive function, and processing speed, this could provide valuable insights. In addition, there is growing awareness of modifier genes and the role of somatic instability in determining age of onset. 10 Gene expression patterns could be influenced by SWB, and age of onset is typically determined solely by motor signs. However, this may not provide a complete picture, as other cognitive domains might offer earlier indicators of both motor and cognitive decline. Notwithstanding these limitations, our preliminary data do not suggest an association between SWB and motor phenoconversion in people with the HD gene mutation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Data reported in this article were collected with support from the National Institutes of Health (NIH), the National Institute of Neurological Disorders and Stroke (R01Act HG002449; PI: Shoulson), L.S. is supported in part extramurally by the Huntington’s Disease Society of America (HDSA), SP0070054; and intramurally by the Scripps Clinic Physician Scientist Scholar Award. He has received research support from the Memorial Sloan Kettering MCP NCI R25 Training, and Northwestern Physician Scientist Training program. L.S. is site PI on the Phase III POLARIS-AD and ALZ-NET studies. L.S. is a paid consultant at Remo Health. D.B. has received personal compensation for consulting, serving on a scientific advisory board, speaking, or other activities with Speaker: Teva Pharmaceuticals, Acorda Therapeutics, Neurocrine Biosciences, Adamas Pharmaceuticals Consulting: Biogen Pharmaceuticals, Amgen Pharmaceuticals, Acadia Pharmaceuticals, Genentech, Inc, GE Healthcare, Gerson Lehrman Group, Guidepoint, L.E.K. C., and has received personal compensation in an editorial capacity for Editor: Annals of Clinical & Translational Neurology. M.C.A. was supported in part by the National Science Foundation Graduate Research Fellowship under grant DGE-2040435. B.K. is supported in part by the National Institute of Aging (K02 AG062745). K.M. and T.P.G. are supported in part by 1R01NS131225-01. A.K.S is supported by the NIA K24AG068312.