
Abstract
One component of geomorphic complexity results from spatial heterogeneity in river corridors. The characteristics of this form of complexity have important implications for habitat and biodiversity, attenuation of downstream fluxes, resistance and resilience of river ecosystems, river processes, ability to characterize patterns and changes through time, and river management and restoration. Numerous measures of complexity have been applied to heterogeneity from spatial scales of bed grain size distribution to entire river networks. Studies explicitly incorporating geomorphic complexity have increased substantially since 2000, but there is no single, widely used metric of complexity. Despite increasingly explicit scientific appreciation of the importance of complexity in river corridors, public attitudes toward rivers continue to emphasize an attractive appearance, which commonly equates to a physically simple and homogeneous river corridor.
I What is complexity?
Thoreau’s advice to “simplify, simplify” notwithstanding, complexity is inherent in rivers and our ability to characterize complexity reflects our understanding of processes and forms in river corridors – defined here as channels and floodplains. Complexity is used in different contexts within physical geography. One context involves complex behavior or complexity as a property of a system. Complexity in this context refers to the simultaneous presence of simple and complicated behaviors or the presence of nonlinear dynamics, self-organization, and emergent properties (e.g., Phillips, 2014; Werner and McNamara, 2007; Table 1). A second context for complexity involves spatial heterogeneity. This paper focuses on geomorphic complexity in the context of spatial heterogeneity, although spatial heterogeneity and complex behavior are commonly interrelated (e.g., Cadenasso et al., 2006; Murray and Fonstad, 2007; Peipoch et al., 2015; Phillips, 2007). Spatial heterogeneity in the following discussion subsumes concepts such as evenness, diversity, and physical complexity.
Examples of definitions of complexity.

Spatial heterogeneity can also be used in different ways and is dependent on the scale of interest (Adler et al., 2001; Li and Reynolds, 1995). Ecologists commonly describe spatial heterogeneity in terms of patches, which are discrete areas that differ in structure, composition, or function (Cadenasso et al., 2006). Spatial heterogeneity of patches can then be described with respect to number of different types of patches, number of each type of patch, spatial configuration of patches, and so forth (Table 1). Recognition of patches gave rise to hierarchical patch dynamics, which conceptualizes an ecosystem as a dynamic mosaic of physically and biologically defined patches (Thorp et al., 2008; Wu and Loucks, 1995). Although there is no directly analogous conceptual model in geomorphology, some of the primary elements of hierarchical patch dynamics, including continuous transient dynamics rather than equilibrium, correspond to concepts in geomorphology (e.g., Tooth, 2000).
As in ecology, spatial heterogeneity in river geomorphology varies with spatial scale and timespan of interest. Spatial heterogeneity of a sinuous river could be characterized with respect to grain diameters on a point bar at a moment in time, individual meander bend migration over decades, meander geometry over river reaches kilometers in length and through centuries, and so forth. Consequently, spatial heterogeneity can be quantified using metrics that emphasize spatial diversity within river characteristics such as cross-sectional or planform geometry at a given time, spatial statistics that identify pattern at varying scales, and metrics that include a temporal component that reflects variations in heterogeneity through time (e.g., Girard et al., 2010). Although complexity can vary with time following disturbance (e.g., Klaar et al., 2009), the remainder of this paper focuses on river geomorphic complexity through space rather than through time.
II Why does complexity matter?
As reviewed in this section, geomorphologists and ecologists have focused on geomorphic complexity in river corridors (channels and floodplains) for at least six reasons. First, complexity in the form of habitat diversity can correlate with biodiversity. Second, complexity influences attenuation of downstream fluxes of diverse materials – water, solutes, mineral sediment, and particulate organic matter. Third, complexity is closely connected to the resistance and resilience of river systems to natural and human-induced disturbances. Fourth, complexity both reflects and influences processes in rivers. Fifth, spatial heterogeneity influences our ability to characterize patterns and to detect changes through time and across space. Finally, because of the assumed or demonstrated correlations between complexity and biodiversity, attenuation, resilience, and process, complexity is increasingly a focus of river management, including restoration.
1 Habitat and biodiversity
River geomorphic complexity in the form of spatial heterogeneity of channel substrate, cross-sectional geometry, planform, or other characteristics does not necessarily correspond to habitat complexity, which reflects niche diversity (the variety of habitats in which an organism can live) (Peipoch et al., 2015). Although a common assumption is that geomorphic complexity links to habitat complexity and ecological function as expressed in biodiversity and bioproductivity, this is not always the case if other influences such as introduced species or limited connectivity constrain biodiversity (Lepori et al., 2005; Palmer et al., 2010). Several studies, however, have found that greater habitat diversity correlates with greater biodiversity and bioproductivity (e.g., Bellmore and Baxter, 2014; Greene and Knox, 2014; Luck et al., 2010; Scott et al., 2003; Wyżga et al., 2012). This correlation underlies the shifting habitat mosaic concept of Stanford et al. (2005), which describes how large floods create and destroy habitats, so that the assemblage of habitat types remains relatively stable and diverse, even though the spatial patterning of floodplain habitats changes through time.
2 Attenuation of downstream fluxes
Geomorphic complexity spanning a range of spatial scales – from small sand ripples or minor bank embayments through channel-spanning logjams to secondary channels and floodplain wetlands – can attenuate downstream fluxes over time scales ranging from minutes for solutes trapped in a small eddy to millennia for sediment stored on a large floodplain. Complexity within a channel can promote flow separation that reduces velocity and transport capacity and increases transient surface storage (Gooseff et al., 2007). Complexity can also cause flow obstructions, backwaters, in-channel sedimentation (Brummer et al., 2006), enhanced hyporheic exchange (Boano et al., 2014), and increased overbank flow and sedimentation (Jeffries et al., 2003). Studies examining correlations between complexity and attenuation of fluxes include those focused on peak and base flows (Westbrook et al., 2006; Woo and Waddington, 1990), stream metabolism and nutrient spiraling (Craig et al., 2008; Ensign and Doyle, 2006), trapping and retention of organic matter (Compson et al., 2009; Sheldon and Thoms, 2006; Small et al., 2008), and sediment storage (Pollock et al., 2007; Wohl and Scott, 2016, in press). A specific example of a positive feedback between complexity and flux attenuation comes from logjams that, by obstructing flow, enhance bank erosion and overbank flow. This leads to channel avulsion and the formation of multi-thread channel planforms, further increasing complexity and attenuation of downstream fluxes (e.g., Collins et al., 2012; O’Connor et al., 2003). Analogously, complexity in the form of riparian vegetation enhances hydraulic roughness, facilitating sediment deposition that creates planform complexity and additional attenuation of downstream fluxes of water, sediment, and organic matter (Corenblit et al., 2009).
Connectivity is another aspect of the attenuation of downstream fluxes. Connectivity can be used to describe lateral, vertical, and longitudinal fluxes within a river corridor (e.g., Ward, 1989). Geomorphic complexity can either increase or decrease different forms of connectivity. Complexity in the form of logjams, for example, decreases longitudinal connectivity of water, solutes, sediment, and organic matter (Beckman and Wohl, 2014), but increases vertical connectivity between the channel and hyporheic zone (Sawyer et al., 2011) and lateral connectivity between the channel and floodplain (Jeffries et al., 2003).
3 Resistance and resilience
Ecologists define resistance as the ability of a system to resist changes in form and process caused by external factors (Webster et al., 1975); a resistant system experiences little change. Geomorphologists have expanded on this to distinguish structural resistance as a result of location sensitivity, or position of each element relative to the point of change (e.g., distance to base level), and transmission resistance, which is the capacity of the system to transmit inputs of matter or energy (Brunsden, 2001). Ecologists define resilience as the persistence of an ecosystem and its ability to return to pre-disturbance conditions following disturbance (Holling, 1973; Webster et al., 1975); a resilient system recovers quickly from disturbance and is persistent rather than transient. Resilience is more commonly referred to in fluvial geomorphology as either (i) response time (Bull, 1991) or the transient form ratio, which describes the transience or persistence of a landform relative to the recurrence interval of the flow that created the landform (Brunsden and Thornes, 1979), or (ii) sensitivity, defined as the severity of a response to a disturbance, relative to the magnitude of the disturbance force (Fryirs, 2016; Reid and Brierley, 2015). In the context of river corridors, resistance, resilience, sensitivity, and related concepts essentially seek to characterize system response to disturbance such as floods or human-induced changes in water and sediment yield and channel geometry.
An example of how complexity can influence resistance and resilience comes from beaver meadows, which are spatially heterogeneous, wet-valley bottoms where numerous beaver dams create obstructions that increase the depth, extent, and duration of overbank flow and maintain high riparian water tables (Westbrook et al., 2006, 2011). Beaver meadows can increase open water area and base flow during dry periods (Hood and Bayley, 2008), thus increasing resistance and resilience to drought. Personal observations in the Colorado Front Range indicate that the complexity of beaver meadows also facilitates resistance to wildfires and to extreme floods.
4 River processes
Geomorphic complexity in the form of spatial heterogeneity can reflect river processes and influence river processes. Complexity of channels and floodplains reflects the history of hydraulic force and sediment movement through time. Choné and Biron (2016), for example, found a strong link between channel mobility and habitat diversity. Examples of complexity influencing river processes include diversity of grain size and bed elevation that increase hydraulic resistance, with consequent effects on hydraulic force and sediment movement (e.g., Yochum et al., 2012); and the piece complexity (branching) of large wood, which influences its mobility within a river and thus the effects of the wood on hydraulics and aquatic habitat (Vaz et al., 2013).
Analyses of changes in channel pattern downstream, across a network, or through time provide a useful illustration of how complexity can influence ability to detect pattern and change. Numerous papers are devoted to characterizing planform geometry of sinuous rivers along which individual meander bend geometry varies, reflecting the difficulty of choosing appropriate metrics to characterize this planform (e.g., Hooke, 2013). In the vertical dimension, uncertainty associated with registration error of aerial imagery increases as terrain relief (one form of complexity) increases (Lea and Legleiter, 2016).
5 River management and restoration
Measures of geomorphic complexity have been used in several studies to compare rivers with different levels of anthropogenic alteration and particularly to compare restored river segments to altered, unrestored river segments and reference segments (e.g., Gooseff et al., 2007; Laub et al., 2012; Legleiter, 2014b; Polvi et al., 2014). A critical consideration here is that context matters: different river segments have differing forms and magnitudes of complexity, so that assessments of complexity in restored rivers must use appropriate reference segments (e.g., Fryirs and Brierley, 2009).
Complexity has been used to assess the effects of specific management actions, such as introducing engineered logjams, on channel form and functions such as sediment storage (Brooks et al., 2004), or the effects of specific human disturbances, such as excess sediment (Bartley and Rutherfurd, 2005). River restoration can also be explicitly designed to increase geomorphic complexity (Wohl et al., 2015), although it remains challenging to determine what types (e.g., bedform versus planform) and what magnitudes (e.g., for bed grain size, maximum grain size versus sorting) of complexity are needed to restore desired river functions (e.g., Lepori et al., 2005). Just as restoration of habitat complexity may not be sufficient to restore biodiversity if other factors constrain biotic communities, so restoration of geomorphic complexity may not create the desired effect if watershed-scale processes such as flow or sediment regimes do not sustain the restored complexity (Roni et al., 2015).
III Measures of complexity
Despite the attention given to geomorphic complexity in river corridors, there is no single broadly applicable, widely accepted geomorphic complexity metric. Instead, there are a plethora of measures that have been employed in different studies during the past twenty years to assess various spatial scales and forms of complexity (Table 2). Quantifications of complexity range from the mathematically relatively simple approaches such as classification of units and count or percent data (e.g., Klaar et al., 2009), through clustering analysis (e.g., Hugue et al., 2014; Nelson et al., 2014), spatial statistics based on moving window averages and variograms or correlograms to test for spatial dependence (e.g., Legleiter, 2014a), to using fractal dimensions (Zhang et al., 2015) to analyze either linear or two-dimensional data for characteristic signals that relate to features such as grain size or bedforms (e.g., Singh et al., 2012; Timm and Wissmar, 2013). Basic categories include measures that characterize the diversity of features (e.g., number of separate functional surfaces), those that characterize the numerical range within a feature (e.g., standard deviation of channel width), and those that describe the arrangement of features in space (e.g., variogram). Of these, measures that characterize numerical range within a feature appear to be the most widely used in studies of river spatial heterogeneity (Table 2).
Examples of metrics used to characterize geomorphic complexity in the form of spatial heterogeneity and diversity within river corridors (after Bartley and Rutherfurd, 2005, Table 1; Laub et al., 2012, Table 2; Polvi et al., 2014, Table 2).
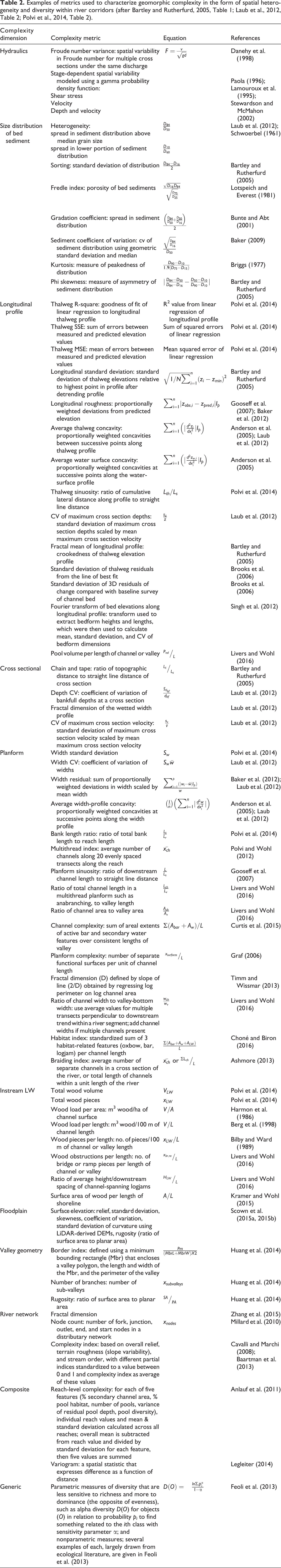
Different measures of geomorphic complexity in rivers are categorized in Table 2 as relating primarily to one of ten distinct aspects of spatial heterogeneity. The lack of consistently used measures likely reflects at least two factors. First, individual studies have different objectives. An investigation of salmonid habitat, for example, is likely to focus on substrate and hydraulic heterogeneity, whereas a study of flood frequency may emphasize heterogeneity of drainage density and topology at the scale of the entire river network. Second, investigators do not necessarily know a priori what types of complexity exert the greatest influence on a particular process or response variable. Studies of connectivity distinguish structural connectivity – the extent to which landscape units are contiguous or physically linked to one another – from functional connectivity, which describes the way in which interactions between multiple structural characteristics of the system in question affect geomorphic, ecologic and hydrologic processes (Wainwright et al., 2011). Analogously, structural and functional complexity can be differentiated. Structural complexity is all forms of spatial heterogeneity within a particular river corridor without reference to any functional effects, whereas functional complexity is spatial heterogeneity that can be shown to affect either a specific process, such as enhanced hyporheic exchange, or a specific response variable, such as biodiversity (Li and Reynolds, 1995; Livers and Wohl, 2016). In an example of a study focused on functional complexity, Schwartz and Herricks (2008) designated mesohabitat units such as pool front and pool rear based on the biological resource needs of different fish species. They then documented differences between fish assemblages and density among distinct units, suggesting that their designated units represented functional complexity.
The majority of the references cited in Table 2 were published after 2000. Given the rapid proliferation of studies attempting to quantify geomorphic complexity and the diverse objectives and spatial and temporal scales considered in these studies, the large number of complexity metrics is not surprising. Although there may be characteristic sources and magnitudes of complexity within distinct types of river corridors (e.g., meandering channel within a forested floodplain versus sand-bed braided channel with minimal woody vegetation), there is unlikely to be a single metric that adequately captures all forms of functional complexity within a river corridor.
IV Where next?
At least three issues represent major challenges to understanding geomorphic complexity in river corridors. The first is understanding sources and forms of complexity in diverse geomorphic process domains. Distinguishing structural and functional complexity, or finding a signal within the noise of diverse scales of spatial heterogeneity in river corridors remains a research challenge. Finally, enhancing public appreciation for complexity in river corridors is a critical component of applying river science.
1 Sources and forms of complexity
Sources of complexity refer to the processes that create and maintain spatial heterogeneity. Where instream wood is an important component of spatial heterogeneity within a channel, for example, the processes of tree growth and mortality, recruitment of wood to the channel, and the transport/storage of wood within the river corridor all influence this spatial heterogeneity. Forms of complexity refer to the categories of spatial heterogeneity listed in Table 1 – hydraulics, bed substrate, channel geometry, and so forth – and to the magnitude and spatial frequency of heterogeneity.
Conceptual understanding of river corridors suggests that there should be characteristic forms and magnitudes of complexity in distinct process domains (Montgomery, 1999) or river styles (Brierley and Fryirs, 2005) (Figure 1). Process domains and river styles are both conceptual frameworks that describe spatially identifiable areas or river segments within a drainage network that are characterized by distinct valley geometry, disturbance regime, and suites of geomorphic processes. Complexity in a steep, narrow bedrock canyon is likely to be associated with heterogeneity of the longitudinal profile and channel width, for example, whereas complexity in a low-gradient river with a broad floodplain may come primarily from a multithread channel planform and patches of distinct floodplain vegetation. Studies associating geomorphic process domains with particular sources and forms of complexity and with distinct biotic communities come mostly from mountainous environments (e.g., Bellmore and Baxter, 2014; Harris, 1988; Polvi et al., 2011), but this approach is amenable to diverse river systems (Fryirs, 2015; Fryirs and Brierley, 2009).
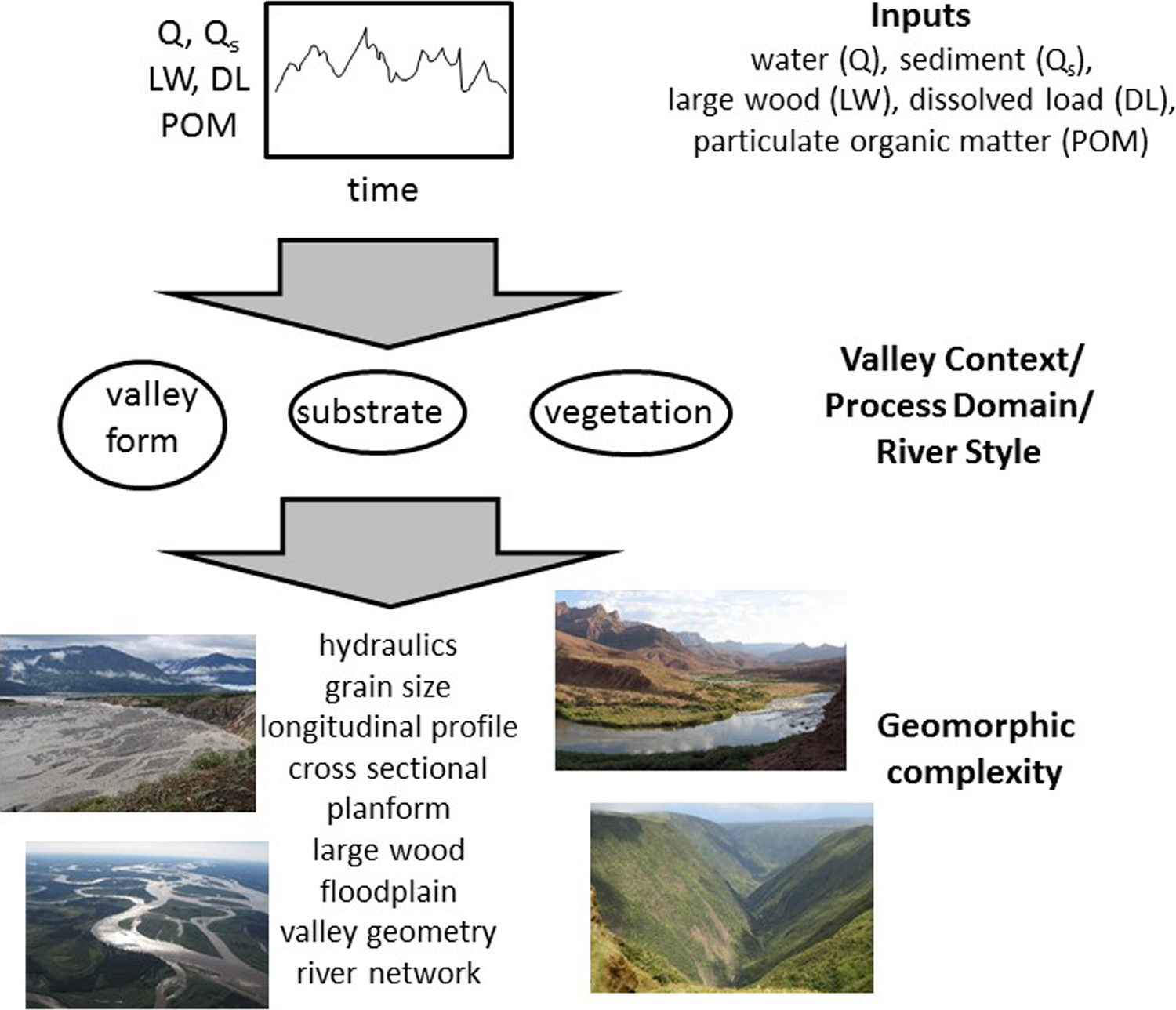
Conceptualization of how inputs interact with valley context to create distinct forms of geomorphic complexity. Inset photos illustrate different process domains and associated complexity.
Closely related to understanding sources of complexity in a particular river environment is identifying first, what aspects of complexity to measure (Blue and Brierley, 2015) and second, what could be described as desirable forms of complexity. Both of these identifications reflect scientific and social context, as exemplified by the Platte River. Figure 2 illustrates a portion of the Platte River near Kearney, Nebraska in 1938 and 1969. In 1938, the channel was a broad, shallow, braided river with high annual and interannual variability of flow and minimal riparian vegetation. Construction of a dam upstream substantially reduced flow variability, which facilitated the establishment of woody riparian vegetation and transformed the channel to an anastomosing planform by 1969. Each of these alternative forms of this river segment has substantial spatial heterogeneity within the river corridor, but the forms of complexity present in 1938 sustained native, endemic Great Plains fishes that are now threatened or endangered partly because of loss of habitat associated with changes in the form of river complexity. In a social context in which sustaining endangered species receives high priority, the complexity present in 1938 is more desirable than the complexity present in 1969.

The Platte River near Kearney, Nebraska.
2 Structural and functional complexity
Identifying desirable forms of complexity relates to distinguishing structural versus functional complexity. Every form of spatial heterogeneity in a river corridor is likely to create an associated function. Heterogeneity of bed grain size, for example, can influence hyporheic exchange, nutrient retention, interstitial habitat, hydraulic roughness, sediment mobility and channel response to fluctuating flows, and other processes. Bed grain size is typically highly heterogeneous in steep channels, yet the complexity associated with this heterogeneity may not be as effective at trapping and retaining particulate organic matter as other forms of geomorphic complexity such as backwater pools at logjams (Livers and Wohl, 2016). The assumption underlying the concept of functional complexity is that identifying distinct sources and forms of complexity can be important in managing rivers when river complexity is constrained by human influences. Many lower gradient alluvial rivers historically had extensive floodplains with wetlands and secondary channels that have now been drained for agriculture (e.g., Wallick et al., 2007). Although existing land use may prevent restoration of the original complexity of the entire floodplain, restoring or maintaining other sources of functional complexity with respect to nitrate retention (e.g., riparian forest buffer zones, instream large wood), for example, can substantially improve water quality.
3 Public perceptions of complexity
A striking gap exists between the appreciation of geomorphic complexity by river scientists, on the one hand, and, on the other hand, the engineering mandate to simplify rivers in order to increase conveyance, along with the public perception that “messy” rivers (e.g., rivers with instream wood (Chin et al., 2008), braided rivers (Kondolf, 2006), or river corridors with floodplain wetlands (Wohl, 2013)) need to be “cleaned up” (McCormick et al., 2015). Episodes of geomorphic change, such as floods or large sediment inputs from hillslope failures, are commonly followed by strenuous efforts to clean up rivers and to put channels back in “their place.” This divergence of perception strongly influences reach-scale river restoration efforts, which can involve simplifying natural and functional river corridors that are perceived as unsightly or unhealthy (Kondolf, 2006).
I think that rivers need the type of public educational campaign that transformed the nineteenth-century image of jungles as dark, dangerous places to the twenty-first-century image of tropical rainforests containing enormous biodiversity and providing critically important ecosystem services. Freshwater organisms are among the most endangered groups of species on Earth (Ricciardi and Rasmussen, 1999). Shifting public attitudes toward rivers from an idealized expectation of a single channel meandering slightly through an open woodland to an appreciation of the enormous diversity and geomorphic complexity of river ecosystems would substantially enhance river management and restoration.
Footnotes
Acknowledgements
I thank Natalie Kramer Anderson, Bridget Livers Gonzalez, DeAnna Laurel, Katherine Lininger, and Dan Scott for helpful comments on an earlier draft of this paper. The paper was substantially improved by comments from two anonymous reviewers.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.