
Abstract
At least some forms of interpersonal violence could follow from a vision of the self as a fierce, dominant creature. This should be particularly true when psychopathic (more proactive, less reactive) tendencies are involved. Possible relations of this type were examined in two studies (total N = 278) in which college student samples were presented with a new, structured version of an old projective test typically used in psychotherapy contexts. Participants were presented with predator–prey animal pairs (e.g., lion–zebra) that were not explicitly labeled as such. For each pair, the person was asked to choose the animal that they would more prefer to be. Participants who desired to be predator animals more often, on this Revised Animal Preference Test (RAPT), tended toward psychopathy to a greater extent. In Study 1, such relations were manifest in terms of correlations with psychopathic traits and with an interpersonal style marked by hostile dominance. Further analyses, though, revealed that predator self-identifications were more strongly related to primary psychopathy than secondary psychopathy. Study 2 replicated the interpersonal style correlates of the RAPT. In addition, photographs were taken of the participants in the second study and these photographs were rated for apparent hostility and dominance. As hypothesized, participants who wanted to be predator animals to a greater extent also appeared more hostile and dominant in their nonverbal behaviors. These studies suggest that projective preferences can be assessed in a reliable manner through the use of standardizing procedures. Furthermore, the studies point to some of the motivational factors that may contribute to psychopathy and interpersonal violence.
Psychopaths are often likened to predators, both in popular press and in scientific psychology. For example, Hare (1996) states that psychopaths are “predatory” and “cold-blooded” and that their actions could be considered a form of intraspecies predation (p. 26). There may be good reasons for analogies of this type. Predators are fierce and dominant, and fearless dominance is a primary component of several models of psychopathy (Benning, Patrick, Hicks, Blonigen, & Krueger, 2003). Predators take advantage of others for their own gain, and this sort of behavior is frequently observed at higher levels of psychopathy (Jones & Paulhus, 2011). And psychopaths engage in a form of aggression—proactive/instrumental—that has been termed predatory in animal models (Weinshenker & Siegel, 2002).
Beyond these parallels, there may be a more fundamental way in which these analogies make sense. In their self-image, and in their relations with others, people with higher levels of psychopathy may wish to be predator-like. That is, they may wish to see themselves as fierce, dominant, and aggressive, and such predatory strivings may underlie at least a portion of their interpersonal behavior. In support of this idea, social cognitive models suggest that wishing to be a certain way portends acting that way (Bandura, 1986; Hoffner & Buchanan, 2005), including in the realm of aggression (Perry & Perry, 1976). And people readily identify with animals, ascribing qualities to them such as independence, nurturance, and aggression, depending on the animal (Borgi & Cirulli, 2015). Indeed, animals may be among the oldest symbols that people used to characterize both themselves and others (Herzog & Galvin, 1992).
The origins of the present work, however, emerge from a number of projective literatures, some (but not all) of which are fairly old. Investigators have used animal pictures in the Thematic Apperception Test (TAT) to encourage a greater amount of fantasy material (Bellak, 1986), particularly among children (Bills, 1950). A rationale here is that animals, relative to people, often exert their motivations in more basic ways, and thus writing about them can facilitate the expression of more basic desires (Bellak, 1986; Bills, 1950). In another projective technique, family members depict each other as animals (e.g., dad is an elephant, mom is a dolphin) and then tell stories about these animals (Arad, 2004). This technique is allied with play therapy (Moustakas, 1953) and is thought to result in significant insights into family dynamics in a nonthreatening manner (Arad, 2004).
Even more pertinent is a projective test called the Animal Preference Test (APT), whose earliest citation appears to be van Krevelen (1955). In the APT, people are asked which animal or animals they would like to be. A person might say “rabbit” if they wish to be cute and “lion” if they wish to be ferocious (Myers, 2002). Rojas (1981) found that people chose animals based on their aggressiveness, nurturance, autonomy, or aesthetic appeal, and Rojas and Tuber (1991) found some relationships between the APT and presenting clinical symptoms, including tendencies toward aggression. Tuber (2012) subsequently detailed ways in which the psychotherapist could use the APT, along with other projective tests, to obtain useful information about a patient’s object relations and affect-regulation style (Westen, 1991).
The APT is an unstructured personality test, as many projective tests are (Symonds, 1949). That is, people give open-ended responses to the APT (van Krevelen, 1955) and test responses are interpreted through clinical insight rather than objective scoring rules (Tuber, 2012). Such features have been heavily critiqued in the personality assessment literature because they tend to result in unreliable procedures with unknown validity (Lilienfeld, Wood, & Garb, 2000). Nonetheless, it seems to us that there is a core insight to the APT. People can identify with animals relatively easily (Borgi & Cirulli, 2015) and the animals that they identify with are likely to provide important clues concerning their interpersonal motivations (Rojas, 1981). In particular, people who identify with predator animals (e.g., a shark or a lion) are likely to possess greater tendencies toward interpersonal violence than people who do not.
To examine hypotheses of this type, we created a revised version of the APT termed the RAPT (Revised Animal Preference Test; Robinson, Bair, Liu, Scott, & Penzel, 2017). Similar to the original APT (van Krevelen, 1955), people are asked which animals they would like to be. However, the test is structured such that a series of animal pairs are presented and people pick their preferred animal (Borgi & Cirulli, 2015). We focus the test on predator–prey relations by constraining the pairs such that one animal is a predator of the other (e.g., lion–zebra). In a projective spirit, though, we do not mention anything about predators and we do not ask people whether they have traits similar to the animals. Accordingly, people’s answers on the RAPT are likely to reveal something about the extent to which they want to be fierce, dominant beings (Rojas & Tuber, 1991) independent of explicit queries along these lines (McClelland, 1987).
Myers (2002) found that boys wanted to be predatory animals (e.g., lions) more than girls did, and Robinson et al. (2017) found a similar trend among adults. Independent of possible trends of this type, we hypothesized that higher levels of predator self-identification on the RAPT would be linked to psychopathic tendencies, broadly defined. In circumplex terms, psychopathy is marked by interpersonal hostility as well as dominance (Rauthmann & Kolar, 2013), and these are qualities that people typically ascribe to predator animals as well (Rojas & Tuber, 1991). Accordingly, we predicted that the RAPT would correlate positively with both hostility and dominance, and that its circumplex location (Gurtman, 2009) would favor the hostile-dominant quadrant of the interpersonal space. We also hypothesized an inverse relationship between the RAPT and the personality trait of agreeableness, which is central to dimensional conceptions of psychopathy (Miller & Lynam, 2015).
In Study 1, we also administered the Levenson Self-Report Psychopathy Scale (LSRPS: Levenson, Kiehl, & Fitzpatrick, 1995), which was designed to capture the Psychopathy Checklist distinction between primary and secondary psychopathy (Hare, 1996) and has been successful in doing so (Brinkley, Schmitt, Smith, & Newman, 2001; Lynam, Whiteside, & Jones, 1999). Choosing to be a predator, it seemed to us, would correlate positively with the interpersonal features of primary psychopathy, which include callousness and manipulation (Hare, 1996). Self-fantasized predators may also have some tendencies toward impulsivity and social deviance (secondary psychopathy), but such tendencies may be weaker. Accordingly, we primarily hypothesized a positive relationship between RAPT scores and primary psychopathy levels.
In Study 2, we sought to expand the analysis beyond self-reported outcomes. Wishing to be tough and dominant should, conceivably, result in expressive behaviors that are apparent to others (Gifford, 1991). In support of this idea, psychopathic tendencies can be identified from thin slices of behavior (Fowler, Lilienfeld, & Patrick, 2009) and perceivers often concentrate on dimensions such as hostility and dominance when making personality-related inferences (Todorov, Said, Engell, & Oosterhof, 2008). Our question, then, was whether high RAPT scorers would seem more dominant and hostile in their interpersonal presentation, as rated from photos (Ambady & Rosenthal, 1992). Findings of this type would suggest that the processes captured by the RAPT manifest themselves, at least to some extent, in relevant nonverbal behaviors.
Study 1
In Study 1, we hypothesized that the motivations captured by the RAPT would translate into higher levels of psychopathy. We examined this hypothesis in two primary ways. First, we administered the LSRPS, which is an often-used instrument for capturing continuous variations in psychopathic traits and tendencies (Brinkley et al., 2001; Lynam et al., 1999; Sellbom, 2011). The scale is also useful for contrasting primary and secondary versions of psychopathy, which overlaps with the Psychopathy Checklist (Hare, 1996) distinction between trait/interpersonal features of psychopathy (Factor 1) and impulsive or criminal behaviors (Factor 2). The RAPT would seem to be more closely related to interpersonal processes than to criminal behavior per se, and we therefore predicted a stronger relationship with primary psychopathy, though this pattern may be most apparent when controlling for overlapping variance.
In addition, we sought to place the RAPT within the interpersonal space of the circumplex (Gurtman, 2009). Psychopathy has been linked to both hostility and dominance (Dowgwillo & Pincus, 2017), and particularly their combination (Rauthmann & Kolar, 2013). We expected that the same would be true of the RAPT.
Method
Participants and procedures
We recruited a sample of 127 (63.0% female; 80.8% Caucasian; M age = 19.79) undergraduate participants from a medium-sized university in the Midwest. They signed up for a social cognition study via SONA software and arrived to the laboratory in groups of six or fewer. After receiving some general instructions, participants were assigned to individual rooms with personal computers. The RAPT, which was an implicit measure, was completed prior to the explicit personality questionnaires. Both studies were run for a predetermined period of time, one that had resulted in sample sizes greater than 100 in previous research, and no participants were dropped prior to analyses.
Measures
Predator self-identification
The instructions for the RAPT asked people to choose animals that they would prefer to be, in Study 1 followed by a series of nine pairs. One animal of each pair was a predator (e.g., “lion”) and the other was a prey animal that the predator sometimes kills and eats (e.g., “zebra”). Multiple pairs (dove–hawk, lion–zebra, shark–fish, seal–polar bear, fly–spider, snake–rat, fox–rabbit, seal–killer whale, and deer–wolf) were presented so that overall scores capture general trends rather than animal-specific attractions. Although the pairs likely convey a sense of predator and prey, the animals are not labeled as such, thus preserving the implicit nature of the test.
We expected people to generally prefer to be predator animals given their lesser vulnerability. This normative trend was apparent in that people preferred to be predator animals 77% of the time, with the 95% confidence interval (CI) excluding .5 [.73, .81]. The standard deviation for the distribution was 0.23, however, meaning that there were pronounced individual differences in predator self-identification tendencies. The high (+1 SD) scorer essentially chose to be predator animals 100% of the time; the low (–1 SD) scorer, by contrast, was generally unbiased with respect to the predator–prey distinction. Importantly, these individual differences were reliable across the nine items of the test (α = .75).
Psychopathy measurement
We administered the LSRPS (Levenson et al., 1995), which is a well-validated (Brinkley et al., 2001; Lynam et al., 1999) 26 item measure of continuous variations in psychopathic traits and tendencies. The primary psychopathy scale (e.g., “For me, what’s right is whatever I can get away with to succeed”) focuses on core psychopathy traits such as selfishness, lack of caring, and manipulation, whereas the secondary psychopathy scale (e.g., “I find myself in the same kinds of trouble, time and time again”) captures tendencies toward impulsive behavior and a self-defeating lifestyle (Hare, 1996; Levenson et al., 1995). Participants indicated how well each statement described them (1 = strongly disagree; 4 = strongly agree), and average scores for both primary (M = 1.99; SD = 0.51; α = .88) and secondary (M = 2.15; SD = 0.44; α = .72) psychopathy were computed. The two forms of psychopathy were significantly, but moderately, correlated with each other, r = .34, p < .001.
Interpersonal style
To examine possible links between predator self-identification and interpersonal style, we administered the hostility, dominance, and arrogance markers of the Revised Interpersonal Adjective Scale (IAS-R: Wiggins, Trapnell, & Phillips, 1988), which was carefully refined over many years (Wiggins & Trapnell, 1996). Participants were asked to characterize their interpersonal behaviors (1 = extremely inaccurate; 6 = extremely accurate) in terms of 16 adjective markers per scale for the dimensions of hostility (e.g., “cruel,” “hard-hearted”; M = 2.04; SD = 0.69; α = .93), dominance (e.g., “assertive,” “forceful”; M = 4.30; SD = 0.69; α = .90), and arrogance (e.g., “boastful,” “cocky”; M = 3.74; SD = 0.75; α = .87). Consistent with circumplex logic (Wiggins & Broughton, 1991), interpersonal arrogance was positively linked to dominance, r = .53, p < .001, and hostility, r = .30, p < .001, but dominance and hostility were largely independent, r = .05, p = .540. Regardless, we expected that self-identified predators would score higher in both dominance and hostility.
Agreeableness
Because the RAPT is an implicit measure, we thought it would predict psychopathic tendencies even after controlling for agreeableness, a personality trait linked to both hostility and psychopathy (Miller & Lynam, 2015). Accordingly, we assessed agreeableness using the established scale of Goldberg (1999), which correlates highly with other measures of agreeableness as well (John & Srivastava, 1999). Participants indicated the degree to which (1 = strongly disagree; 5 = strongly agree) 10 agreeableness statements (e.g., “have a soft heart”) describe the self, and an average score was computed (M = 4.07; SD = 0.61; α = .85).
Results
Does predator self-identification predict psychopathy?
Simple regressions established that self-identified predators tended toward both primary psychopathy, t = 4.39, p < .001, β = .37, and secondary psychopathy, t = 2.08, p = .039, β = .18, with estimated means (Aiken & West, 1991) shown in Figure 1. The relationship with primary psychopathy was stronger and is likely to be more robust when controlling for overlapping variance. To examine this possibility, we created a residual primary psychopathy score that was uncorrelated with secondary psychopathy and a residual secondary psychopathy score that was uncorrelated with primary psychopathy (Tangney, 1995). Predator self-identification correlated with the purified primary psychopathy score, r = .30, p < .001, but not with the purified secondary psychopathy score, r = .06, p = .495. Thus, there is a somewhat unique relationship between predator self-identifications and primary psychopathy.
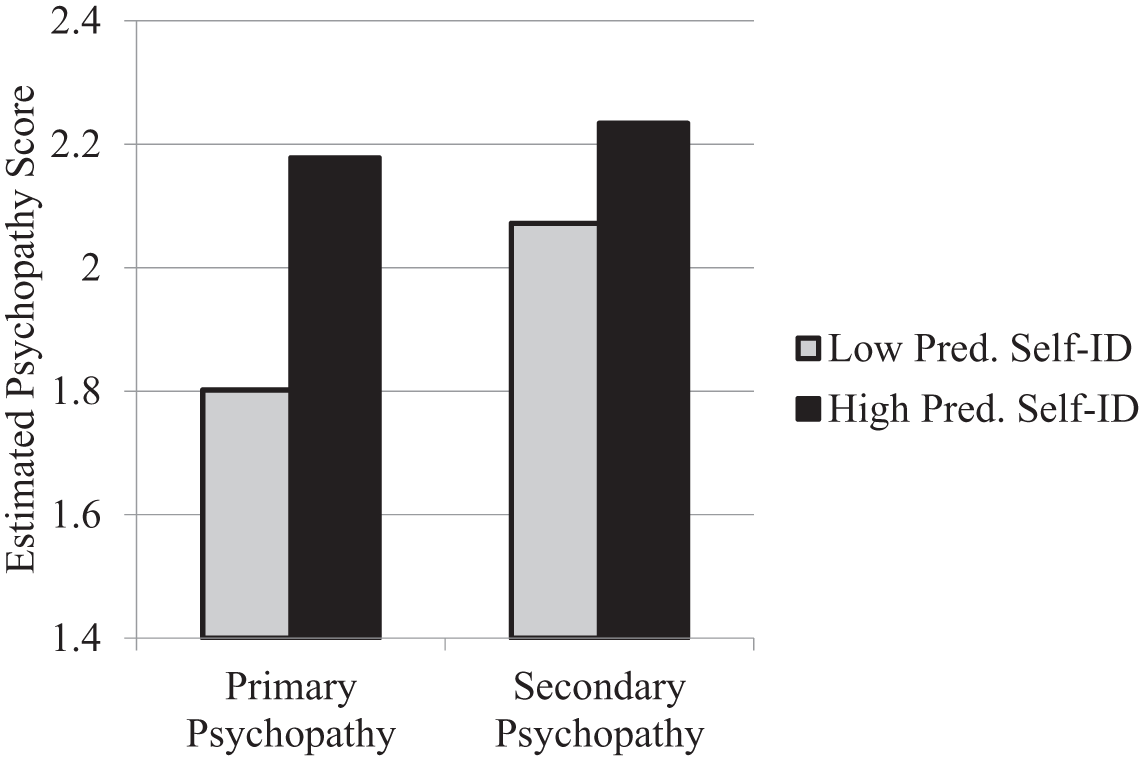
Estimated psychopathy means as a function of low (–1 SD) and high (+1 SD) levels of predator self-identification, Study 1.
What is the interpersonal signature of the RAPT?
The interpersonal signature of psychopathy includes tendencies toward both dominance and hostility (Jones & Paulhus, 2011), and we expected a similar interpersonal profile for the RAPT. Consistent with these expectations, self-identified predators scored higher in IAS-R dominance, r = .29, p = .001, IAS-R hostility, r = .35, p < .001, and the diagonal vector (Wiggins & Broughton, 1991) of IAS-R arrogance, r = .31, p < .001. Thus, aspiring predators were both hostile and dominant.
As a way of more precisely locating the RAPT within the interpersonal space, we used the procedures of Wiggins and Broughton (1991), who presented formulas for determining the circumplex placement of measures as a function of their relationships with dominance and warmth (hostility) factor scores. We applied these procedures to the RAPT. As shown in Figure 2, the interpersonal signature of the RAPT appears to be one that combines hostility and dominance, a quadrant that also encompasses tendencies such as narcissism, psychopathy, and aggression (Jones & Paulhus, 2011; Wiggins & Broughton, 1991).

Circumplex locations of high RAPT scorers, Studies 1 (“1”) and 2 (“2”).
Additional analyses
A one-way ANOVA revealed a fairly pronounced sex difference on the RAPT, F(1, 125) = 68.64, p < .001,
Multiple Regression Results for the Sex by RAPT Interaction Term, Studies 1 and 2.

Note. Sex did not moderate relations between the RAPT and the outcome variables. For these analyses, males were coded as −1 and females were coded as +1. RAPT = Revised Animal Preference Test; IAS-R = Revised Interpersonal Adjective Scale.
As one might expect, there was a negative correlation between predator self-identification scores and the personality trait of agreeableness, r = −.27, p = .002. However, the RAPT continued to predict primary psychopathy, interpersonal dominance, and interpersonal hostility (but not secondary psychopathy) when controlling for agreeableness, as shown in Table 2.
Predator/Outcome Relationships After Controlling for Agreeableness, Studies 1 and 2.
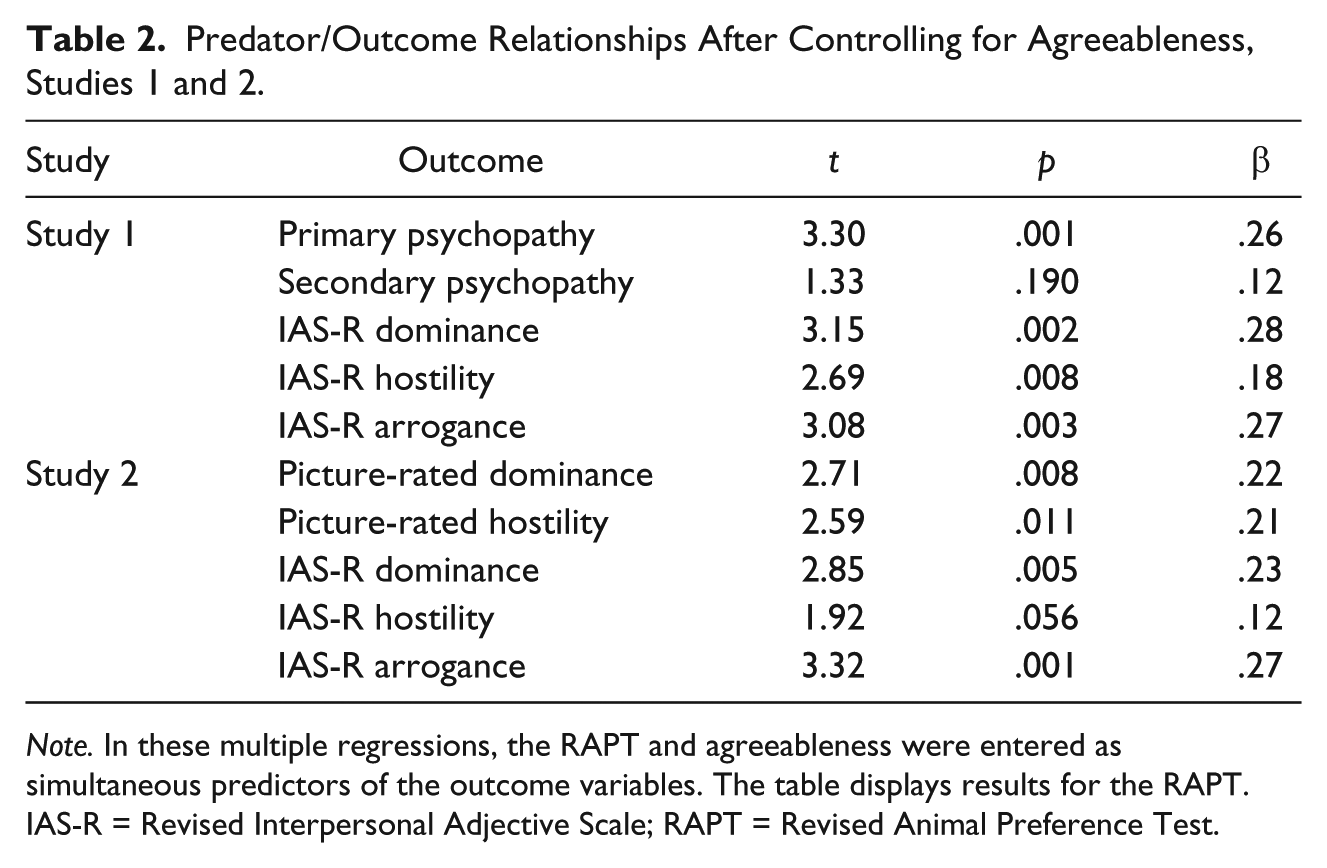
Note. In these multiple regressions, the RAPT and agreeableness were entered as simultaneous predictors of the outcome variables. The table displays results for the RAPT. IAS-R = Revised Interpersonal Adjective Scale; RAPT = Revised Animal Preference Test.
Discussion
Instead of asking people whether they are aggressive, or even want to be aggressive, the RAPT simply asks people which of two animals they would prefer to be. Despite the innocuous, somewhat playful framing of the test, we composed it such that one animal often kills and eats the other. Under these conditions, which animal one prefers to be could, or perhaps should, reveal something important about one’s dominance strivings (Arad, 2004; Levy, 1985). In support of these ideas, we found that people who self-identified as predators more frequently scored higher in primary psychopathy, and they tended toward an interaction style marked by hostility, dominance, and arrogance. Accordingly, Study 1 provides some initial evidence for the idea that the RAPT has value in ordering people in terms of their tendencies toward psychopathy. In Study 2, we sought to replicate and extend these ideas.
Study 2
The first goal of Study 2 was to replicate the circumplex findings of Study 1. In this connection, we were particularly interested in whether the angular displacement for the RAPT would remain stable across studies, which would suggest some degree of certainty in the probable correlates of predator self-identification (Gurtman, 2009). Somewhat more ambitiously, we were interested in whether the interpersonal strivings captured by the RAPT possess implications for nonverbal behavior (Gifford, 1991). People who are motivated to be dominant often display such goals through mechanisms such as an erect body posture and direct eye contact (Knight & Mehta, 2014). Similarly, hostility can be signaled through cues such as furrowed brows and nonsmiling (Oosterhof & Todorov, 2008). In Study 2, we sought to determine whether RAPT-identified predators display some of these cues, as manifest in the expressive information available in nonposed photographs.
Method
Participants and procedures
We recruited a new sample of 151 (68.9% female; 89.4% Caucasian; M age = 19.53) undergraduates from the same university as Study 1. They completed the RAPT prior to completing the IAS-R and the agreeableness measure. Participants were also photographed at the end of the session (see below).
Measures
Predator self-identification
We added three new pairs to the predator self-identification measure of Study 1 (hawk–rabbit, gopher–snake, and wolf–sheep). For each of the 12 pairs, participants were encouraged to choose the animal that they would rather be by making a mouse click on the corresponding button. As in Study 1, we scored these choices in terms of whether predator (1) or prey (0) animals were chosen and then averaged across items. There was a normative preference for predator animals, M = 0.68; 95% CI from .64 to .73, but individual differences were also apparent (SD = .27). As in Study 1, high scorers almost always preferred to be predator animals, whereas predator–prey status seemed less relevant to the choices of low scorers. The internal reliability of the measure was good (α = .84).
Picture ratings of dominance and hostility
Toward the end of the study, the computer screen informed the participant that this portion of the study was done. The participant then informed the experimenter, who stated that the last task involved taking a picture of the participant. The participant was directed to stand in front of a cream-colored wall area in the main room, at which point a picture was taken using a Sony NEX 3NL compact digital camera. The picture was taken in landscape format from about 4 feet back and depicted the person from the waist to head. Experimenters approached the task in a low-key manner to capture natural forms of expression. That is, participants were not prompted to pose in any particular way. After the picture was taken, the participant was debriefed and allowed to leave.
At the completion of the study, we created a MediaLab program that randomly ordered the pictures for two rating tasks. Six judges (graduate students and undergraduate research assistants), who were blind to RAPT scores as well as hypotheses, rated how dominant (1 = not very dominant; 6 = very dominant) and agreeable (1 = not very agreeable; 6 = very agreeable) each participant appeared. As with the IAS-R warmth dimension, we reverse-scored agreeableness ratings and refer to this reversed dimension in terms of hostility (Wiggins & Trapnell, 1996). There was a fair degree of rating agreement across judges, and we therefore averaged across the six ratings per participant to quantify individual differences in displayed dominance (M = 3.57; SD = 0.65; α = .57) and hostility (M = 2.98; SD = 0.90; α = .84).
Interpersonal style
Participants also characterized their own interpersonal tendencies in terms of 16 adjectives each for the IAS-R (Wiggins et al., 1988) dimensions of dominance (M = 4.28; SD = 0.72; α = .89), hostility (M = 1.85; SD = 0.58; α = .91), and arrogance (M = 3.52; SD = 0.73; α = .84). As in Study 1, arrogance was correlated with dominance, r = .50, p < .001, and hostility, r = .23, p = .004, but dominance and hostility were uncorrelated, r = −.05, p = .575.
Agreeableness
The RAPT is an implicit measure that may predict tendencies toward hostile dominance even after controlling for explicit variations in personality. To determine whether this was the case in Study 2, we asked participants to complete Goldberg’s (1999) International Personality Item Pool (IPIP) agreeableness scale, which was also administered in Study 1 (M = 4.15; SD = 0.53; α = .89).
Results
Do self-identified predators display hostile dominance?
We thought it possible that self-identified predators would seem dominant and hostile to others, like predators.
Simple regressions confirmed these hypotheses in that people with higher RAPT scores appeared more dominant, t = 2.64, p = .009, β = .21, and hostile, t = 2.87, p = .005, β = .23, in their pictures. Estimated means for these results are displayed in Figure 3.
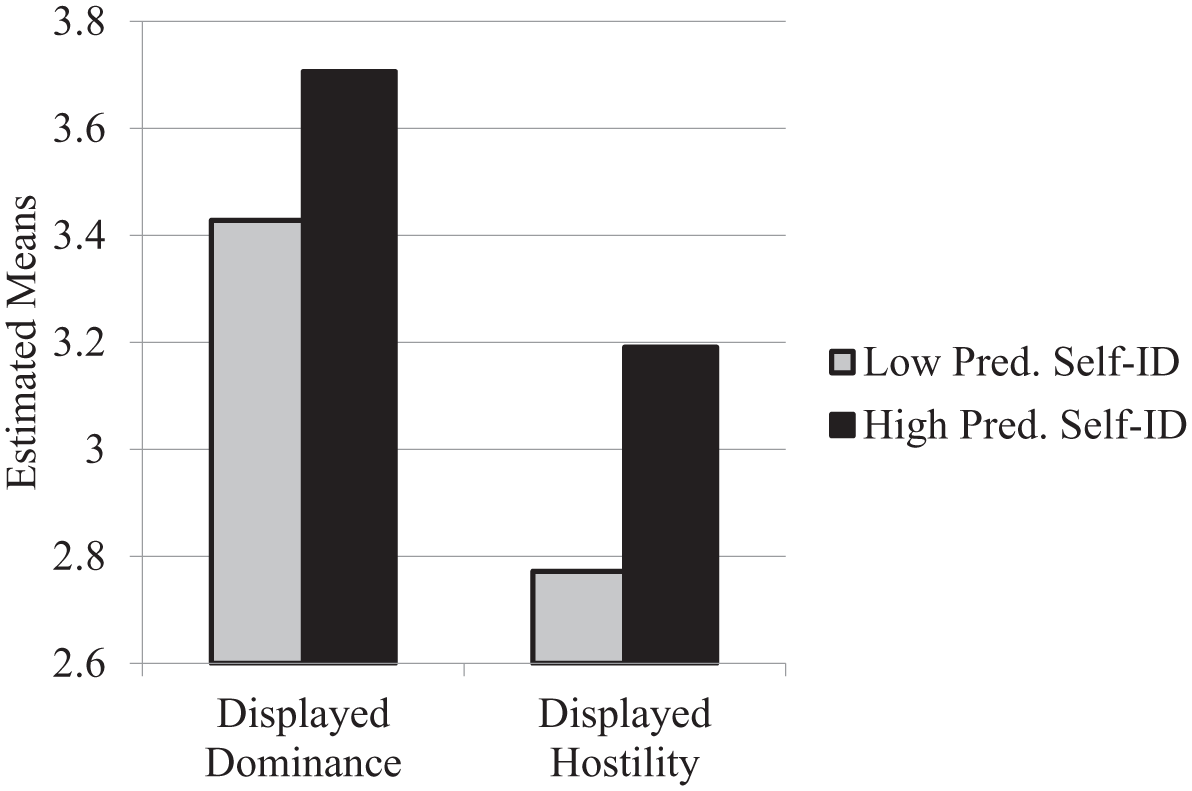
Estimated pictorial displays as a function of low (–1 SD) and high (+1 SD) levels of predator self-identification, Study 2.
What is the interpersonal signature of the RAPT?
In Study 1, people who wanted to be predators more frequently characterized their interpersonal behavior as more dominant, hostile, and arrogant. The same was true in Study 2. Specifically, higher RAPT scores were linked to higher IAS-R scores for the dimensions of dominance, r = .23, p = .006; hostility, r = .19, p = .018; and arrogance, r = .27, p < .001. The strongest correlation was for arrogance, which can be thought of as a form of dominance that is hostile rather than friendly (Wiggins & Trapnell, 1996). Consistent with this line of thinking, the angular displacement (Wiggins & Broughton, 1991) for the RAPT placed it in the hostile-dominant portion of the circumplex, just as in Study 1 (see Figure 2).
Additional analyses
As in Study 1, men (M = .85) received higher RAPT scores than women (M = .61), F(1, 149) = 30.92, p < .001,
Discussion
Study 2 results replicated those of Study 1. At high levels of predator self-identification, people wanted to be predators nearly 100% of the time; at low levels, by contrast, the predator–prey distinction did not appear to be particularly salient. Furthermore, people receiving higher RAPT scores tended toward higher levels of hostility, dominance, and arrogance and these relationships tended to remain significant when controlling for agreeableness. Moreover, we were able to place the RAPT in the same region of the circumplex space (see Figure 2): The RAPT captures a version of dominance that is hostile rather than friendly.
The circumplex location of the RAPT is consistent with the psychopathy construct (Rauthmann & Kolar, 2013). In addition, one can use this circumplex location to make other predictions about high RAPT scorers. By circumplex logic (Gurtman, 2009), high RAPT scorers should receive higher scores on measures of antisocial personality, they should be status-seeking, and they should be prone to aggression (Wiggins & Broughton, 1991). By contrast, they should not be deferent and they may lack interpersonal self-control (Wiggins & Broughton, 1991). Finally, their interpersonal problems will be due to self-enhancement and manipulation rather than self-effacement and a lack of assertiveness (Horowitz, 2004). These considerations add to the likely profile of RAPT-linked predators and they suggest useful areas of future research.
In addition, a major purpose of Study 2 was to determine whether predator-choosing people appear hostile and/or dominant to others. We examined such questions using nonposed pictures because there is evidence that people display features of their personality in this context (Naumann, Vazire, Rentfrow, & Gosling, 2009) and because the context removes any verbal or spoken cues (Ambady & Rosenthal, 1992). Under these conditions at least, we found a positive relationship between RAPT scores and inferred hostility as well as a positive relationship between RAPT scores and inferred dominance. Future research might focus on the specific facial (Oosterhof & Todorov, 2008) or postural (Gifford, 1991) cues responsible for these inferences. Regardless, these Study 2 results suggest that self-identified predators seem to present themselves in ways that signal hostile dominance.
General Discussion
Psychodynamic clinicians have often-used versions of an APT to study themes of object relation among their clients (Arad, 2004; Tuber, 2012). The genius of the test is that it is make-believe (e.g., one will never become a rabbit or a shark) and yet responses can reveal core interpersonal motivations that the self has (Rojas & Tuber, 1991). Still, the APT is typically given in an unstructured, intentionally projective way, rendering its suitability for large-scale personality assessment largely unknown.
In this context, we sought to create a structured version of the APT termed the RAPT. The RAPT is structured in the sense that one animal of each pair is a predator of the other and all choices could therefore indicate something about the person’s desires to be like predators—that is, fierce, dominant, and aggressive (Rojas, 1981). The test is projective, though, in that it involves make-believe and does not ask people to report on their traits or behaviors (McClelland, 1987). Still, the stimuli are not particularly ambiguous, unlike some other projective tests (Bornstein, 2002). Altogether, then, the RAPT could be thought of as a projective/objective hybrid with the potential to capture important motivational dynamics toward hostile dominance.
Consistent with this reasoning, we found that people who chose predator animals on a greater proportion of trials were more dominant and hostile in their interpersonal style and they tended toward interpersonal arrogance, which combines features of dominance and hostility. Moreover, they scored higher in primary psychopathy and appeared more hostile and dominant in their nonverbal displays. These findings begin to make a case for the idea that there are motivational factors in psychopathy and that the RAPT captures some of them. In the General Discussion, we focus on implications, questions/limitations, and future directions.
Implications and Analysis
The RAPT does not ask people what traits they have or even which animal better symbolizes the self. Rather, the RAPT is projective: People choose animals that they want to be, independent of reality. Under these circumstances, what people choose may reflect motivations that the self has, particularly within its interpersonal context (Tuber, 2012). That the RAPT predicted hostility, dominance, and psychopathy therefore suggests that people who have psychopathic traits may wish to have such traits, at least to a certain extent. Or, more specifically, they may wish to see themselves as fierce and dominant in their interactions with others, who can be metaphorically likened to prey (Hare, 1996). The present findings provide unique evidence for this motivational perspective of psychopathy.
At the same time, the RAPT related more substantially to primary psychopathy in Study 1, at least when controlling for overlapping variance. These results make sense. Envisioning the self as a predator should more intuitively capture the interpersonal features of psychopathy (Factor 1) relative to lifestyle factors (Factor 2). Or, stated another way, the predatory mind-set may be more relevant to hostile than to impulsive forms of psychopathy (Miller & Lynam, 2015). Thus, there are some limitations to a RAPT-based view of psychopathy.
Even so, an advantage of the RAPT is its implicit nature. Because the test does not explicitly ask about hostility or dominance, it is likely to predict a number of outcomes even after controlling for explicit measures (Cogswell, 2008). We supported this idea by controlling for agreeableness in the present studies. Furthermore, given that the RAPT targets motivations without explicitly asking about motivations, the RAPT may be likely to predict the sorts of outcomes highlighted in the implicit motivation literature (McClelland, 1987). These include spontaneous behaviors and affective reactions to interpersonal events (Schultheiss, 2008). We provided some support for these ideas in the form of spontaneous nonverbal behaviors in the nonposed picture task (Study 2), but further data would be welcome.
Questions, Limitations, Applications, and Future Research
Juxtaposing RAPT responses with the psychopathy literature reveals an interesting dissociation. Many people wanted to be predators on the RAPT, but tendencies toward psychopathy are not normative (Hare, 1996; Miller & Lynam, 2015). A useful way of thinking about this dissociation is that the RAPT engages make-believe rather than reality. In their visions for the self, many people, and especially boys or men, may fantasize themselves as “tough” if not exploitative or aggressive (Konijn, Bijvank, & Bushman, 2007; Rojas & Tuber, 1991). In reality, though, people tend to temper behavioral trends of this type (Horowitz, 2004; Wiggins & Broughton, 1991). Regardless, people who are more interested in being predators, relative to those who are less interested, are more prone to hostile dominance or psychopathy.
Although we focused on psychopathy, the RAPT could have some relevance to other members of the Dark Triad. This may be so given that the RAPT aligns itself with the regions of the interpersonal circumplex common to all Dark Triad members (Jones & Paulhus, 2011). Even so, we are inclined to think that the RAPT may have some particular relevance to psychopathy relative to narcissism or Machiavellianism. Narcissism is primarily a form of self-enhancement (Sedikides & Gregg, 2008) and this trait is not a robust predictor of predatory forms of aggression (Bobadilla, Wampler, & Taylor, 2012). Machiavellianism, furthermore, has some submissive elements to it that seem inconsistent with a predatory approach to the social environment (Rauthmann & Kolar, 2013). Accordingly, predator self-identifications may relate more strongly to psychopathy than to other members of the Dark Triad, though this suggestion would clearly benefit from additional research.
This line of thinking points to further outcomes that should vary with the RAPT. People scoring high in psychopathy are self-centered, meaning that they will take advantage of others when opportunities arise (Jones & Paulhus, 2011). Similarly, we would hypothesize that self-identified predators would exhibit self-serving behavior in realms such as the dictator game or paradigms designed to capture social value orientation (van Lange, 1999). Relatedly, people scoring high in psychopathy tend to be callous as well as low in empathy. Robinson et al. (2017) provided some evidence that self-identified predators experience empathy less frequently, but further data of this type would be valuable. Finally, we were able to link the RAPT to hostile dominance, which is a robust predictor of aggression (Wiggins & Broughton, 1991), but more direct investigations of aggressive behavior should be conducted. In this connection, though, it would be particularly useful to differentiate proactive and reactive aggression because the RAPT is likely to predict the former more than the latter (Glenn & Raine, 2009).
We also admit that ours was basic personality research. What this means, among other things, is that further research will be necessary to determine whether and how the present individual differences matter for serious forms of violence or criminality. What we can say here is that basic probes of personality possess value within forensic contexts (Sellbom, 2011) and that violence researchers have called for new assessments of the present implicit type (Skeem & Monahan, 2011). Implicit measures are useful because they can bypass concerns related to social desirability while identifying key motivational processes that contribute to interpersonal behavior (Bornstein, 2002). Our sense is that the RAPT should have value for such reasons. Among people who are prone to violent or antisocial behavior, the RAPT might predict the severity and/or nature of such behavior. For example, high RAPT scorers should be more prone to risk-taking behavior and to crimes that are often conducted for instrumental reasons, such as theft or robbery (Glenn & Raine, 2009). They may also get into fights more often, particularly when dominance contests are involved (Terburg & van Honk, 2013).
Furthermore, we can mention some potential areas of application. As an educational tool, the RAPT could be used to highlight tendencies toward hostile dominance and their probable basis in fantasy. Indeed, even young children should be able to grasp the significance of their projective identifications (Tuber, 2012). Occupationally, the RAPT could be used to identify applicants whose motivational strivings could be problematic within a particular job, such as police person, prison guard, or soldier (Paulhus, 2014). In forensic settings, the RAPT could be useful in identifying trends of a psychopathic type, particularly when combined with other information like interviews, records, and third-party reports (Hare, 1996). And the RAPT could have a variety of applications within therapy and counseling settings (Arad, 2004; Tuber, 2012). Indeed, a fairly large number of psychological and behavioral problems follow from interpersonal factors (Horowitz, 2004) and people often lack insight into the ways in which the self contributes to such problems (Westen, 1991). In this connection, the RAPT could serve as a basis for self-insight, an important catalyst of therapeutic change (Karoly, 2012).
To realize some of these outcomes, sample diversity should be expanded. As mentioned, there are reasons for thinking that the RAPT should predict similar outcomes among children, adolescents, and younger adults (Tuber, 2012). And our sense is that the RAPT would function similarly among people of different races or socioeconomic backgrounds. However, we are uncertain whether the RAPT would function similarly among older adults, particularly given its basis in make-believe, and we know of no relevant research using samples from countries other than the United States. Accordingly, we recognize the limitations of the present samples and encourage future research whose focus includes generalizability questions.
Finally, it may be useful to wonder about the direction of the relationships observed. For example, does predator self-identification predispose one toward psychopathy or do psychopaths gravitate toward predator animals? A third possibility, and one that we favor, is that people who harbor competitive, dominant strivings score higher in psychopathy and also prefer predator to prey animals on the RAPT. From this perspective, the RAPT likely predicts psychopathic traits and tendencies because it assesses sources of motivation that contribute to these traits and tendencies (McClelland, 1987; Tuber, 2012).
Conclusion
In a structured version of an APT, people reliably differed in the extent to which they wanted to be predatory (fierce, dominant) or nonpredatory animals, independent of reality constraints. Of more importance, predatory self-identification rates predicted interpersonal arrogance, primary psychopathy, and nonverbal behaviors consistent with hostile dominance. We conclude that a structured projective test—the RAPT—can assess motivational factors pertinent to the psychopathy construct.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.