
Abstract
Recent research underscores the importance of social learning to the development of food preferences. Here, we explore whether social information about edibility—an adult placing something in his or her mouth—can be selectively tied to certain types of entities. Given that humans have relied on gathered plant resources across evolutionary time, and given the costs of trial-and-error learning, we predicted that human infants may possess selective social learning strategies that rapidly identify edible plants. Evidence from studies with 6- and 18-month-olds demonstrated that infants selectively identify plants, over artifacts, as food sources after seeing the same food-relevant social information applied to both object types. These findings are the first evidence for content-specific social learning mechanisms that facilitate the identification of edible plant resources. Evolved learning mechanisms such as these have enabled humans to survive and thrive in varied and changing environments.
For many species, plants are central to survival as potential sources of food. As omnivores whose diets are composed of a wide variety of plant and animal matter, humans are no exception. Gathered plant resources have formed the basis of human diets across evolutionary time (Cordain et al., 2000; Ungar & Sponheimer, 2011), yet many plants are poisonous, and even deadly, when ingested (Keeler & Tu, 1983). Therefore, successfully determining which plants in a local environment are edible has been an essential task throughout human evolution. There are no morphological features common to all edible or to all poisonous plants (Brill & Dean, 1994; Peters, O’Brien, & Drummond, 1992), and identifying the specific edible and poisonous plants in a given environment through trial-and-error experimentation would be extremely costly. Such circumstances favor the evolution of social learning strategies (e.g., Barrett & Broesch, 2012; Boyd & Richerson, 1985, 1988; Galef, 2001, 2009) for acquiring information about edible plants.
Despite the ubiquity of plants and the essential role they have played throughout human evolution, there has been surprisingly little investigation of the ways in which humans acquire information about plants over the course of development. A large body of work has explored young children’s knowledge of the properties that are common to both animals and plants as members of the overarching category “living things” (e.g., Carey, 1985; Inagaki & Hatano, 2002; Keil, 1989; Medin & Atran, 1999). Such research has shown, for instance, that preschoolers understand that plants, like animals, can grow, heal themselves, and reproduce (Backscheider, Shatz, & Gelman, 1993; Inagaki & Hatano, 1996; Stavy & Wax, 1989). However, studies of children’s acquisition of plant-specific knowledge have been few and far between (for exceptions, see Hickling & Gelman, 1995; Nguyen & Gelman, 2002). Recent research with adults has begun to elucidate the cognitive processes that enable humans to locate edible plant resources (Krasnow et al., 2011; New, Krasnow, Truxaw, & Gaulin, 2007), but the strategies that enable humans to identify the edible plant resources in natural environments are not known.
In fact, developmental processes concerning human food learning in any context have received surprisingly little attention, although the few existing studies underscore the importance of social learning. For example, young children prefer foods that an adult calls “yummy” (Lumeng, Cardinal, Jankowski, Kaciroti, & Gelman, 2008), and infants prefer foods offered by individuals who speak the same language as they do (Shutts, Kinzler, McKee, & Spelke, 2009). Infants also readily emulate the food preferences of both neutral and prosocial individuals, but not those of antisocial individuals (Hamlin & Wynn, 2012). In addition, social learning is a well-studied component of nonhuman animals’ food learning (e.g., Barker, Best, & Domjan, 1977; Galef, 2001; Santos, Hauser, & Spelke, 2001).
However, young children’s decisions about what to eat are, famously, not determined by simply copying adult behavior. Under certain circumstances young children will attempt to consume things that they have never seen an adult eat (e.g., sponges and imitation dog feces; Rozin, Hammer, Oster, Horowitz, & Marmara, 1986), and infants do not make consistent inferences about the edibility of certain items they witness adults consume (e.g., sugar or liquids from bowls and glasses; Shutts, Condry, Santos, & Spelke, 2009). Therefore, social learning appears to operate in concert with other processes. Thus far, research has identified factors such as sensitive periods (Cashdan, 1994, 1998) and later-emerging aversions (e.g., disgust, contamination, and neophobia; Cashdan, 1998; Rozin, 1990) as relevant to food learning.
We reasoned that, in addition, there might be cases in which social information about edibility is selectively tied to certain types of entities. Given that humans have relied on gathered plant resources across evolutionary time, and given the costs of trial-and-error learning, we hypothesized that human infants may possess selective social learning strategies that rapidly identify edible plants. Similar cases in which social information is preferentially tied to certain types of entities have been established in other domains (e.g., learning about danger; Barrett & Broesch, 2012; Cook & Mineka, 1990; DeLoache & LoBue, 2009). To test this hypothesis, we examined whether infants would preferentially identify a plant, over an artifact, as a food source after they saw an adult place objects from both entities in his mouth. We conducted four experiments, recruiting 6- and 18-month-olds from the greater New Haven, Connecticut, area and testing them in the Infant Cognition Center at Yale University.
Experiment 1
Method
Participants in our first experiment were 32 healthy, full-term 18-month-olds (16 female, 16 male; mean age = 18 months 17 days, range = 18 months 4 days to 18 months 30 days) who heard English spoken more than 50% of the time. Eleven additional infants were tested but excluded: because of failure to make a choice during testing (n = 5), failure to look at both options before choosing (n = 2), fussiness (n = 1), procedural error (n = 1), or video loss due to equipment failure (n = 1), or because the behind-ear action that was part of the procedure was used for hearing-aid training in the participant’s home (n = 1).
Our stimuli were a plant and an artifact, each with four removable objects (dried apricots or dried plums) attached. The plant consisted of realistic-looking artificial leaves and branches arranged in a clay pot. The artifact was designed to control for many of the individual perceptual features of the plant: It was made from the same leaves and branches as the plant, but they were spray-painted silver, stuck into a beige base, and encased in a glass cylinder. Thus, it had individual parts of the same shape, texture, and number as the plant, but did not itself look like a plant. The dried fruits were arranged to look like fruits growing on the plant; they were placed in the same spatial configuration on the artifact (for details, see Supplementary Methods and Figure S1a in the Supplemental Material). Which type of dried fruit (apricot or plum) was placed on the plant and which was placed on the artifact was counterbalanced across participants.
Infants sat on their parents’ laps to watch a show, and parents were instructed not to direct their infants’ attention. The show began with two introduction events during which a curtain opened to reveal Experimenter 1 sitting between the plant and artifact; directly in front of him was a board with two white squares. Experimenter 1 said, “Hi, [infant name]!” while waving, and then the curtain was lowered.
During the next two events, Experimenter 1 performed the same action on the plant and artifact, in a counterbalanced order. Half of the infants saw a food-relevant in-mouth action: The curtain opened, and then Experimenter 1 said, “Hi, [infant name],” turned toward the plant or artifact, removed a dried fruit, paused, and placed the topmost part of the fruit in his mouth while saying “Hmmm” and looking down. He did not make eye contact with the infant after saying “Hi” and acted as if he was tasting the fruits for himself rather than explicitly demonstrating an action. The curtain lowered during the end of his vocalization. This procedure was then repeated a second time (with the plant if the experimenter had performed the action on the artifact in the first event, and with the artifact if he had performed the action on the plant). The other half of the infants saw a food-irrelevant behind-ear action, which was the same as just described except that the experimenter placed the fruit behind his ear. Previous investigations used similar in-mouth and behind-ear actions as examples of food-relevant and food-irrelevant actions, respectively (Santos et al., 2001; Shutts, Condry, et al., 2009). In the final event sequence, the curtain rose, and Experimenter 1 said, “Hi, [infant name],” removed the remaining dried fruits from the plant and artifact, and placed each set of fruits on the white square closest to the object from which they came (see Video S1 in the Supplemental Material).
Finally, parents closed their eyes, and Experimenter 2, who was blind to which fruits came from which object, offered infants a choice (see, e.g., Feigenson & Carey, 2003; Hamlin, Wynn, & Bloom, 2007). Experimenter 2 ensured that the infants saw both fruit types, asked, “Which one can you eat?” in the in-mouth condition and “Which one can you use?” in the behind-ear condition, and brought the board within the infant’s reach (see Video S1). Two independent coders, who were blind to which fruits came from which object, coded each infant’s choice as the first fruit type he or she touched (intercoder agreement = 100%) after verifying that the infant looked at both options prior to making the choice (intercoder agreement = 97%).
The following were counterbalanced across participants: the fruit type on the plant (i.e., dried apricots on the plant and dried plums on the artifact, or vice versa), the plant’s position during the show and choice (right or left side), the object Experimenter 1 acted on first (plant or artifact), and the object from which Experimenter 1 removed the remaining fruits first (plant or artifact).
Results and discussion
Coders blind to the target of the actions (plant vs. artifact) rated the affect Experimenter 1 displayed in all of his in-mouth and behind-ear actions and also recorded their durations. They found no differences between actions directed at the plant and those directed at the artifact (for these analyses for Experiments 1 through 3, see Supplementary Analyses, Section 1.1, in the Supplemental Material).
We predicted that if infants preferentially identify plants as food sources, participants who viewed the in-mouth action would choose the fruits from the plant over those from the artifact, even though they witnessed the same action demonstrated with both. Infants who observed the behind-ear action, in contrast, were predicted to choose both fruit types equally often. As predicted, infants who saw the in-mouth action chose the fruits from the plant (13 of 16 infants; binomial p = .02 1 ). Infants who saw the behind-ear action chose randomly (7 of 16 infants chose the fruits from the plant; binomial p = .80), a significantly different pattern of response than observed in the in-mouth condition (Fig. 1), Pearson χ2(1, N = 32) = 4.80, p = .03. Thus, despite seeing the same social information (the in-mouth action) applied to both object types, infants preferentially identified the plant as a food source.
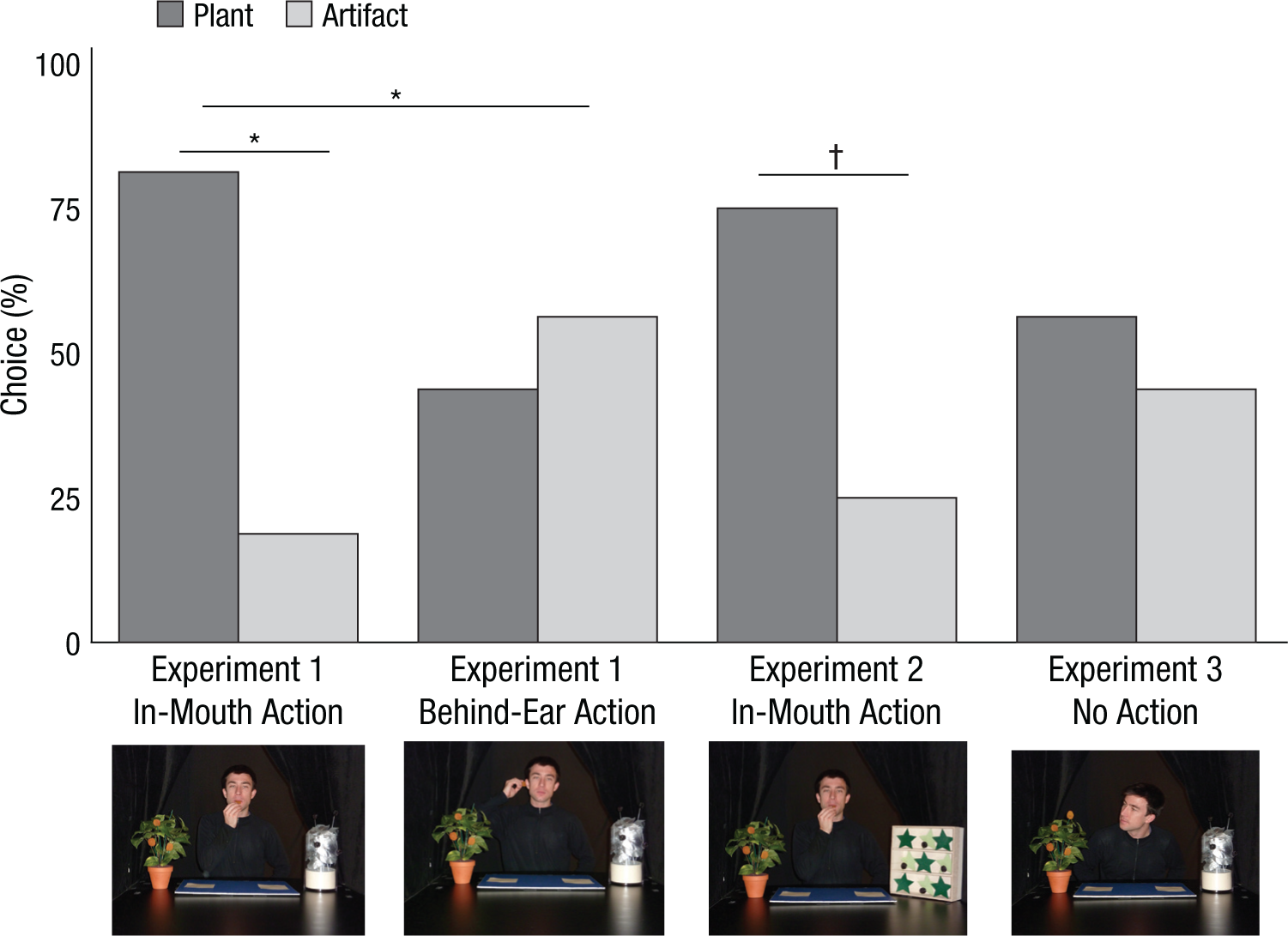
Choices of 18-month-olds in Experiments 1 through 3. Asterisks indicate significant differences (†p < .08, two-tailed; *p < .05, two-tailed).
Experiment 2
The results for the behind-ear condition rule out the possibility that infants generally prefer objects from plants over those from artifacts. However, our plant stimulus was likely more familiar to infants than our novel artifact. To rule out the possibility that infants prefer items from familiar sources over items from novel sources in food contexts specifically, we tested a new group of 18-month-olds using a more familiar artifact.
Method
Participants were 16 healthy, full-term 18-month-olds (8 female, 8 male; mean age = 18 months 15 days, range = 18 months 2 days to 19 months 2 days) who heard English spoken more than 50% of the time. An additional 3 infants were tested but excluded for failure to choose (n = 2) and fussiness (n = 1).
The procedure was identical to that for the in-mouth condition of Experiment 1, except that a set of shelves was substituted for the novel artifact (see Supplementary Methods and Fig. S1b in the Supplemental Material). This afforded a strong test of familiarity in food contexts: Not only have urban and suburban Western infants been exposed to shelves (see Supplementary Methods for confirmation that our set of shelves would be more familiar to infants than the novel artifact), but in their direct experience, most food comes off of shelves (e.g., in a cupboard or refrigerator) that contain objects of various shapes and colors. Thus, we put other items, stars, on our set of shelves; the stars were different shades of green, like the plant leaves. Infants saw Experimenter 1 perform the in-mouth action with dried fruits taken from the plant and from the set of shelves, and Experimenter 2 asked them, “Which one can you eat?” As before, two independent blind coders determined infants’ choices (intercoder agreement = 100%) and verified that infants saw both options before choosing (intercoder agreement = 100%).
Results and discussion
Again, as predicted, infants chose the fruits from the plant, replicating our finding from Experiment 1 (Fig. 1). The number of infants choosing the fruits from the plant over the fruits from the shelves (12 of 16 infants; binomial p = .077) was nearly identical to the number of infants choosing the fruits from the plant over the fruits from the novel artifact in Experiment 1 (13 of 16 infants), notwithstanding infants’ familiarity with shelves and lack of familiarity with the novel artifact used in Experiment 1. Despite a lifetime of experience with food coming from shelves, after viewing the in-mouth action, infants identified the items from the plant, over those from the shelves, as edible.
Experiment 3
One explanation for our findings in Experiments 1 and 2 is that infants view all plants as food sources, or conversely, apply a rule that items from artifacts are generally inedible. If so, then when asked, “Which one can you eat?” infants would have selected fruits from the plant even without seeing a food-relevant in-mouth action. But because many plants are poisonous, unpalatable, or difficult to digest, viewing every plant as edible without some positive evidence that it is safe—such as seeing an adult put it in his or her mouth—would be costly. We therefore hypothesized that in the absence of a social cue of edibility, infants would no longer preferentially identify a plant as a food source.
Method
Participants were 16 healthy, full-term 18-month-olds (7 female, 9 male; mean age = 18 months 20 days, range = 18 months 0 days to 19 months 1 day) who heard English spoken more than 50% of the time. An additional 3 infants were tested but excluded for failure to choose.
The stimuli and procedure were the same as in Experiment 1, except that infants saw Experimenter 1 simply look pointedly at the plant and the novel artifact in turn while keeping his arms at his sides and saying, “Hmmm”; the curtain lowered during the end of each of his vocalizations. In the next phase, the curtain rose, and Experimenter 1 removed the dried fruits from both the plant and the artifact, placing them on the same board used in the previous experiments. Experimenter 2 then presented the board to the infants and asked, “Which one can you eat?” Two independent blind coders again determined infants’ choices (intercoder agreement = 100%) and verified that infants saw both options before choosing (intercoder agreement = 100%).
Results and discussion
In this case, infants chose randomly between the plant and the artifact (Fig. 1; 9 of 16 infants chose fruits from the plant; binomial p = .80). Without viewing the in-mouth action, infants no longer identified the plant as a more likely food source.
Experiments 1 through 3 demonstrate that by the age of 18 months, infants preferentially identify a plant as a source of food on the basis of a single brief exposure to an adult putting an object from the plant into his mouth. Unlike some nonhuman primates who view any object placed in the mouth by an experimenter as food (Santos et al., 2001), human infants use additional criteria for determining edibility. The combination of the action and the ontological status of the object—in this case, that it is a plant—appears to be critical.
For Experiments 2 and 3, parents completed a questionnaire assessing their infants’ experience with plants, including whether the infants had ever eaten fruit directly off a plant before or had previously eaten dried apricots or plums. None of the variables measured were related to infants’ choices (see Supplementary Analyses, Section 1.2, in the Supplemental Material). Nevertheless, 18-month-olds have had several months of experience with solid foods. In Experiment 4, we used a looking-time paradigm to test whether much younger infants—6-month-olds, who typically are only just starting to eat mashed-up baby foods and have very limited (if any) experience with solid food items—preferentially identify plants as sources of food.
Experiment 4
We used a violation-of-expectation paradigm, in which infants tend to look longer at unexpected or surprising events (e.g., Baillargeon, Spelke, & Wasserman, 1985; Thomsen, Frankenhuis, Ingold-Smith, & Carey, 2011). If 6-month-olds selectively identify plants as potential food sources, they should look longer when the in-mouth action is performed with fruits from the artifact than when it is performed with fruits from the plant, but should look equally long when the behind-ear action is performed with fruits from the plant and when it is performed with fruits from the artifact.
Method
Participants were 32 healthy, full-term 6-month-olds (16 male, 16 female; mean age = 5 months 28 days, range = 5 months 15 days to 6 months 15 days). Sixteen additional infants were tested but excluded because of failure to watch more than one pair of Experimenter 2 events (n = 6), failure to watch Experimenter 1 events (n = 1), procedural error (n = 6), fussiness (n = 2), and looking-time differences between plant and artifact trials greater than 2.5 standard deviations from the mean (n = 1).
Infants were evenly divided across two conditions: the in-mouth condition and the behind-ear condition. All infants sat on their parents’ laps throughout a two-phase presentation; parents were instructed not to direct their infants’ attention. Each phase was preceded by a single introduction event in which the experimenter said, “Hi, [infant name]!” and waved. Phase 1 was identical to Experiment 1, except that Experimenter 1 remained motionless at the end of each event while the infant’s looking time was recorded: Experimenter 1 performed either the in-mouth or the behind-ear action sequence with one dried fruit taken from the plant (Event 1) and one taken from the novel artifact (Event 2; in a counterbalanced order), then removed the remaining fruits from their sources and placed them on two white squares on the board (Event 3; see Video S2 in the Supplemental Material).
The plant and artifact were removed from the stage for Phase 2, and Experimenter 2, who was blind to which fruits came from which object, appeared seated behind the board. After the single introduction event, there were six alternating test trials during which Experimenter 2 performed either the in-mouth or the behind-ear action sequence with the dried fruits on the board, alternating between the two squares (three times each; see Video S2). After each action sequence, Experimenter 2 remained motionless as infants’ looking times were recorded. Parents’ eyes were closed during the test trials.
Each looking-time trial ended when the infant looked away for 2 consecutive seconds or 30 s elapsed. Looking times were determined by an on-line coder, who was blind to event type (i.e., whether the dried fruit came from the plant or artifact), using the jHab computer program (Casstevens, 2007). A second blind coder evaluated the trials of 25% of the participants; intercoder agreement was 99%. Counterbalancing was the same as in Experiments 1 through 3, with the addition of counterbalancing of the fruit type Experimenter 2 acted on first.
Results and discussion
As in Experiments 1 through 3, blind coders rated the events and found no differences between actions directed at the plant and at the artifact (see Supplementary Analyses, Section 2.1, in the Supplemental Material).
A repeated measures analysis of variance examined the effects of object type (plant vs. artifact) and presentation order (plant vs. artifact acted on first) on infants’ looking times (for details, see Supplementary Analyses, Section 2.2, in the Supplemental Material). As predicted, infants looked longer at in-mouth actions when they were performed with fruits from the artifact than when they were performed with fruits from the plant (Fig. 2); this was true for the in-mouth actions of both Experimenter 1, F(1, 14) = 7.21, p = .018, η p 2 = .34, and Experimenter 2, F(1, 14) = 12.63, p = .003, η p 2 = .47. These results suggest that preverbal infants expect plants, relative to artifacts, to be sources of food. In contrast, infants who viewed the behind-ear action looked equally at the plant and artifact events performed by Experimenter 1, F(1, 13) = 0.84, p = .38, η p 2 = .06, and Experimenter 2, F(1, 14) = 0.002, p = .97, η p 2 = .00; thus, there was no general tendency for infants to look longer at any action performed with the novel artifact.
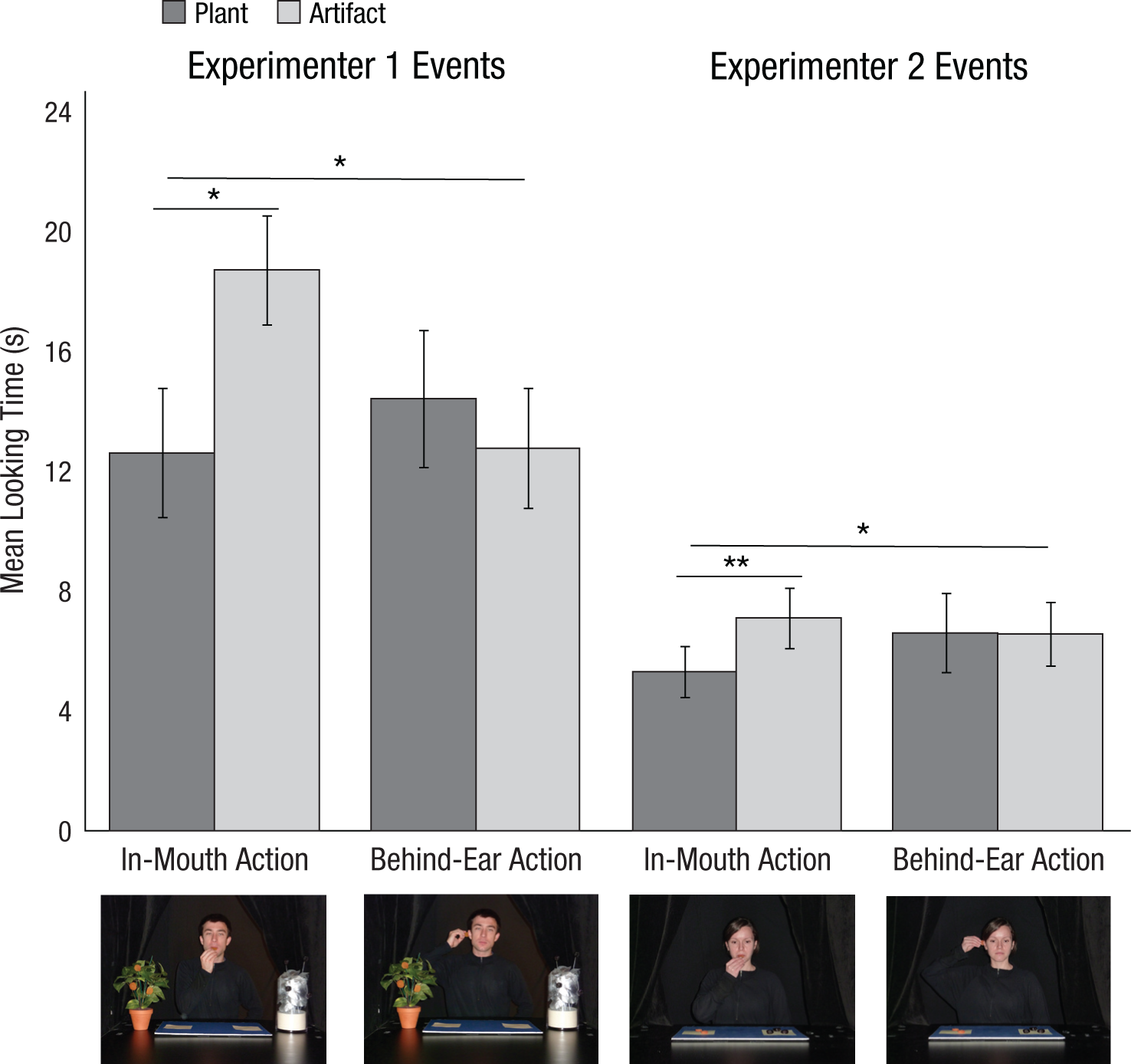
Mean looking times of 6-month-olds in Experiment 4. Error bars represent ±1 SEM. Asterisks indicate significant differences (*p < .05, two-tailed; **p < .01, two-tailed).
Overall, infants’ looking patterns differed significantly between the in-mouth and behind-ear conditions. The Object Type × Condition interaction was significant for both Experimenter 1 events, F(1, 27) = 7.39, p = .01, η p 2 = .22, and Experimenter 2 events, F(1, 28) = 4.54, p = .04, η p 2 = .14. These results confirmed that preverbal infants’ differential expectations about plants were constrained to the food-relevant in-mouth condition. Prior to extensive experience with solid foods, 6-month-old infants appear to expect plants to be food sources.
General Discussion
The findings reported here indicate that after viewing the same food-relevant social information—an adult placing something in his or her mouth—applied to a plant and an artifact, 6- and 18-month-old infants preferentially identify the plant as the food source. A series of control conditions and questionnaire information ruled out alternative explanations—including prior experience eating fruit directly from plants—and indicated that viewing the food-relevant social information was necessary for identifying plants as food sources.
These findings add to the growing literature highlighting the role of social learning in the development of food preferences (e.g., Hamlin & Wynn, 2012; Lumeng et al., 2008; Santos et al., 2001; Shutts, Kinzler, et al., 2009). However, our results show that infants do not use social information alone, but also take into account other relevant contextual information—in this case, the kind of object acted upon. This type of selective social learning has been demonstrated before (e.g., Barrett & Broesch, 2012; Cook & Mineka, 1990; DeLoache & LoBue, 2009), and is similar to classic cases of prepared learning (Garcia & Koelling, 1966; Seligman, 1970), but to our knowledge, this is the first demonstration that human infants make selective inferences based on social information about edibility.
To be clear, we are not claiming that plants are the only entity treated this way. For example, given the long history of humans processing food prior to consumption (Wrangham, 2009), it is likely that cues of preparation, such as chopping or cooking, may also be privileged. Similarly, extensive ontogenetic experience of certain objects being associated with food (e.g., plates, spoons, etc.) may privilege items associated with those objects in certain contexts. This is an area that is ripe for future exploration. Another important avenue for future research will be investigating the ways in which the content of the social information and the ontological status of the object interact. The actors in the current experiments deliberately conveyed neutral information (i.e., they said, “Hmmm” instead of “Mmmm”). Understanding how infants react to negative information applied to an entity in a privileged category and positive information applied to other entities will be essential to mapping the development of food preferences.
Of course, selective social learning processes like those identified here will not underlie all aspects of human food learning. Human food-learning processes are likely to be complex combinations of many underlying factors, including other types of social learning (e.g., Hamlin & Wynn, 2012; Lumeng et al., 2008; Santos et al., 2001; Shutts, Kinzler, et al., 2009), avoidance processes (e.g., Cashdan, 1998; Rozin, 1990), sensitive periods (e.g., Cashdan, 1994), and pedagogical demonstrations of food consumption and preparation (e.g., Csibra & Gergely, 2011).
Limitations and alternative explanations
Because this is the first investigation of selective social learning of plant edibility, there are several outstanding issues that remain to be addressed. For example, the plant we used in the current experiments was a prototypical leafy green plant with fruits growing on it. It remains to be seen whether the selective social learning we observed will generalize to other types of plants, and what combination of features infants use to identify an object as a plant in the first place. Some of our other work provides insight into the latter issue by suggesting that infants do not simply use single features such as green color, leaf-shaped objects, or parts moving relative to the whole to identify plants (Wertz & Wynn, 2014). Instead, it seems that infants rely on a probabilistic combination of features that has yet to be fully uncovered.
Similarly, although we have ruled out several alternative explanations for the current findings, including a general attraction to plants, bias toward familiarity in food contexts, and certain ontogenetic experiences (e.g., general experience with solid foods and specific experiences with plants, such as fruit picking), other plausible alternatives remain. For example, we have characterized this selective social learning system as resulting from the ancestrally recurrent problem of identifying edible plants, but it is possible that it stems instead from other processes operating over either phylogenetic or ontogenetic time, or that it is instead part of a broader learning system that includes many different types of entities.
In any case, the current results leave open the question of how this kind of system develops over the first few months of life. Although we have ruled out certain experiences, there must be relevant environmental inputs; even “maturational” processes require the appropriate environmental conditions for development to occur (e.g., Maurer & Lewis, 2001). Some of these inputs might be counterintuitive, and in principle, a learning system like this could be built (either entirely or in part) via earlier learning processes. The task going forward will be to identify and characterize the relevant inputs and describe how they contribute to the ontogenetic development of this selective social learning system.
Conclusions
Our current results demonstrate that, holding social information constant, infants preferentially identify plants (over artifacts) as food sources. This tendency is present early in ontogeny, prior to any formalized instruction, and mirrors the ancestrally recurrent problem humans faced with respect to identifying edible plant resources. Our other work shows that, absent any social information from adults, infants selectively refrain from touching plant leaves (Wertz & Wynn, 2014), which suggests that specialized responses to plants extend beyond inferences about candidate food items. Although more work is needed to characterize the nature of the cognitive mechanisms underlying these results, they are consistent with the existence of evolved learning mechanisms.
Generally speaking, these types of learning mechanisms enable the accumulation of cultural knowledge across many generations (Boyd & Richerson, 2006) and explain how a universal cognitive architecture can give rise to systematic cultural and individual differences (Tooby & Cosmides, 1992). For example, every individual may be equipped with the same social learning rule for identifying edible plants, but the specific knowledge banks individuals develop will depend on the types of plants and the existing cultural context that they encounter across their lifetimes. Cognitive adaptations such as these have enabled humans to survive and thrive in varied and changing environments.
Footnotes
Acknowledgements
We thank D. Pietraszewski, A. Spokes, K. Duffy, and members of the Infant Cognition Center at Yale University for their help in data collection. We also thank P. Bloom, F. Keil, and K. Hinde for helpful comments on earlier versions of this manuscript.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by National Science Foundation Grant BCS-0715557 and National Institutes of Health Grant R01 MH 081877 to K. Wynn.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.