
Abstract
In semi-arid ecosystems where lacustrine sediments are rare, bio-proxies preserved in fluvial deposits are needed to understand environmental changes. In this study, we evaluate the potential of phytoliths as a bio-proxy in the Yamé River’s deposits at Ounjougou (Mali, West Africa) covering the middle to late Holocene (7790–4000 cal. BP). In soils, phytolith assemblages result mainly from decomposition of local vegetation but in alluvial deposits, the taphonomy of phytoliths is more complex, depending on the type of transport and deposition. In order to define the spatial origin of the phytolith assemblages, either from local (valley floor) or regional vegetation (catchment area), we took the sedimentary characteristics of the deposits into account. Using a combination of general and index approaches, phytoliths of 14 modern and 29 fossil samples from the Yamé valley were analyzed. The predominant source area of represented phytoliths varies with the fluvial energy of transport. Channel deposits, carried during periods of strong fluvial activity, contain higher numbers of savanna grass phytoliths and display a lower ratio of dicotyledon versus Poaceae phytoliths (D/P) than those deposited in the floodplain where phytoliths from the local gallery forest are more strongly represented. From the 5th millennium BP onwards, higher percentages of grass short cell phytoliths (GSCP) and lower D/P values point to gradual vegetation opening due to increasing aridity. High amounts of burned phytoliths show regular fire incidence in the gallery forest and attest for human impact on the landscapes of Ounjougou from the 7th millennium BP onwards. After 4500 cal. BP, there is evidence for pearl millet cultivation.
Keywords
Introduction
Phytolith analysis offers new perspectives for reconstructing ancient vegetation in fluvial environments. Because of their siliceous structure, phytoliths are well preserved in terrestrial sediments whereas pollen preservation is mainly restricted to anoxic waterlogged deposits. In alluvium deposits, phytoliths can be found in sandy and gravelly as well as in clayey sediments. But studies of modern and fossil phytolith assemblages in alluvium deposits are scarce. In West Africa, Holocene and Late Pleistocene vegetation reconstruction is mainly based on palynological and anthracological data from lacustrine, marine sediments and archaeological sites (Ballouche and Neumann, 1995; Bouimetarhan et al., 2009; Eichhorn and Neumann, 2012; Höhn and Neumann, 2012; Lézine, 1989; Lézine et al., 2005; Maley, 1981; Salzmann and Waller, 1998; Salzmann et al., 2002).
The studies conducted at Ounjougou (Mali) for the early Holocene (Neumann et al., 2009) and in southern Cameroon for the Late Pleistocene (Sangen et al., 2011) were the first to apply phytolith analysis in fossil fluvial sediments of tropical Africa. They have shown the potential of the technique but many questions remain concerning phytolith taphonomy. Phytolith dispersal is influenced by several factors, such as wind and water transport, fire and grazing herbivores, and depends on the type of site examined (Fredlund and Tieszen, 1994; Kerns et al., 2001; Pearsall, 2000; Piperno, 2006; Prebble et al., 2002). A study on the contribution of phytoliths to the suspended load of rivers in Cameroon has shown that particulate biogenic silica (BSi) transported by rivers can be an essential component of phytoliths assemblages from the top soils (Cary et al., 2005). Thus, for a valid interpretation of phytolith assemblages from alluvial sediments, various processes of transport and deposition have to be considered.
This paper aims to (1) confirm the potential of phytoliths as bio-proxies in fluvial sediments and (2) evaluate the relationship between the stratigraphic characteristics and phytolith assemblages in the middle- to late-Holocene sediments of Ounjougou. Information on the sedimentary facies and geomorphological processes enhances the understanding of transport and depositional context of the phytolith assemblages. Sediment taphonomy is an important issue because it helps to determine the source area of phytoliths and therefore to delimit the spatial scale of vegetation reconstruction. The main aim of this study is to define, through sediment analyses, the origin of the phytolith assemblages in a Sahelo-Sudanian context, either as local (valley floor) or regional (catchment area). On the regional scale, the phytolith assemblages permit vegetation reconstruction, especially for the grass layer (which is not possible with the means of pollen analysis) and the assessment of precipitation changes (Fredlund and Tieszen, 1994). The knowledge of vegetation composition on the local scale is important for the evaluation of human impact on the landscape.
This study is based on the analysis of 14 modern and 29 fossil samples from the site complex Ounjougou. With a long and rich sedimentary record (Le Drézen et al., 2010; Lespez et al., 2011; Rasse et al., 2006) and archaeological remains giving evidence of ancient occupation and land use (Huysecom et al., 2004; Mayor et al., 2005; Ozainne et al., 2009), the Ounjougou reach is exceptional for West Africa. Palaeoenvironmental research at Ounjougou has so far mainly focused on the early- and late-Holocene periods (Le Drézen et al., 2010; Neumann et al., 2009; Ozainne et al., 2009). For the middle and the beginning of the late Holocene (7790–4000 cal. BP), anthracological data and hydrosedimentary dynamics have been described in recent publications (Eichhorn and Neumann, 2012; Le Drézen et al., 2010; Lespez et al., 2011). During this period significant climatic changes took place in the Sahara and West Africa with the establishment of drier conditions between 7500 and 2500 cal. yr BP (deMenocal et al., 2000; Gasse, 2000; Kröpelin et al., 2008; Lézine et al., 2005; Russell et al., 2003; Salzmann and Waller, 1998; Salzmann et al., 2002; Shanahan et al., 2006) and the emergence of agriculture around 4500–4000 cal. BP (Kahlheber and Neumann, 2007; Manning et al., 2011).
Study area and previous work
The site Ounjougou is located 12 km east of Bandiagara and covers an area of 10 km2 in the upper section of the Yamé catchment, on the sandstone plateau (Figure 1). The Yamé River, a tributary of the Niger and the main stream of the Dogon plateau is characterized by continuous but irregular flow because of the Sudano-Sahelian seasonal rainfall regime. At Ounjougou, the Yamé corresponds to the succession of pool-riffle sequences sculpted in the sandstone bedrock and floodplain pockets enclosed in the Pleistocene formations (Lespez et al., 2011). A series of gullies incises into a complex sequence of Quaternary aeolian, alluvial and colluvial deposits. The sedimentary sequence of Ounjougou is covered by a large corpus of radiocarbon dates (Lespez et al., 2011; Ozainne et al., 2009; Rasse et al., 2006).

Geomorphological map of the Ounjougou reach. 1: sandstones; 2: main glacis; 3: upper glacis; 4: floodplain; 5: hydrographic network and permanent lakes; 6: sandy channel; 7: main gullies; 8: location of the transects; 9: location of surface samples.
The vegetation in the study area is a Sahelo-Sudanian savanna strongly altered by human activities. During the rainy season, the whole catchment area is covered by fields, mainly with pearl millet in mixed-crop systems. The persistence of water during the dry season makes Ounjougou a focal area for human activities in the Dogon country. Today, the valley floor at Ounjougou is subject to intensive gardening and is one of the last watering places used by pastoralists before the Inner Niger-Bani Delta, 120 km to the west.
The middle- to late-Holocene chronostratigraphy is divided into three units (HM2, HM4, and HR1) and two subunits (HR1A and HR1B) (Table 1). The sediment and charcoal remains indicate an environmental context different from the one during the early Holocene with more open vegetation and stronger fluvial energy. The middle- to late-Holocene sediments are characterized by the development of thin layers of silt material in the floodplain, particularly during HR1B. This indicates an active channel slightly incised into a swampy floodplain, corresponding to sandy deposits during flood flows and numerous standing water pools with fine organic sedimentation during the low-stage. Based on the exceptional preservation of organic remains and the absence of oxidation borders and desiccation cracks, permanent high water-tables in the valley floor can be inferred. This finer sedimentation can also be explained by the denser vegetation on the Dogon plateau (Lespez et al., 2008, 2011). Charcoal data indicate a gallery forest with extrazonal Sudano-Guinean elements and dense stands of the bamboo species Oxytenanthera abyssinica, resulting in lower runoff, water discharge and sediment delivery. From around 5000 cal. BP, a relatively open vegetation developed, favoured by regular bush fires but also by a marked dry season, attested by the appearance of species such as Tamarindus indica. During this period, Sudano-Guinean taxa gradually disappeared and were replaced by Sudanian and Sahelian-Sudanian species (Eichhorn and Neumann, 2012).
Chronology of the stratigraphic units (after Ozainne et al., 2009) and the samples analysed in this study.

In the charcoal assemblages, species of the local gallery forest are strongly over-represented compared with the regional vegetation (Eichhorn and Neumann, 2012; Neumann et al., 2009). Owing to regional influx, phytolith analysis at Ounjougou offers the possibility to broaden knowledge on the surrounding savannas, woodlands and agroforestry systems and to specify the grassland composition.
Material and methods
Geomorphological framework and sampling
To understand the origin of phytoliths in alluvial sediments, the geomorphological approach, according to fluvial architecture concepts, permits the interpretation of depositional conditions. These are based on the identification of granulometrical and bedding properties.
The grain-size composition of the deposits is determined by the energy of the river and related to geomorphological units such as channel and floodplain. The deposits are coarser and more abundant in the channel where fluvial activity is stronger. In the floodplain, grain size is more homogenous and finer because of the distance to the channel and decantation in shallow water. Distribution and grain size characteristics of the deposits also depend on the nature of the vegetation cover. We suggest that the predominant source area of phytoliths represented in the deposits varies with the fluvial energy of their transport. Coarse deposits carried during periods of strong fluvial dynamic activity contain a greater proportion of phytolith assemblages from the surrounding area than those deposited locally with low energy in the floodplain.
Before sampling, the fluvial architecture of the middle- to late-Holocene deposits was established. Twenty-nine stratified samples were taken from all lithofacies of the exposed profile. They range in age from the middle Holocene (7790–5320 cal. BP) to the first unit of the late Holocene HR1 (5320–4000 cal. BP) and originate from three different sections: Balanites, Detarium and Ravin Sud (Table 1, Figure 1). In addition, 14 modern samples were collected along three transects from river bed to interfluves in the Yamé valley.
Sedimentological analysis and depositional environment identification
For the fossil samples, sedimentological analyses were conducted to identify architectural elements and sediment transport types. The method uses facies assemblage identification and grain-size analysis (Lespez et al., 2011). Facies assemblages were identified and interpreted in terms of architectural elements based on Miall (1996) and comparison with modern sediments from the valley floor. Granulometry was measured by laser particle analyser (Coulter Counter LS 200), adjusted for measurements of the fraction ≤ 2 mm. Coarse material was removed and grain-size was determined using a stack of sieves ranging from 2 to 5 mm. In particular, the CM pattern (Passega, 1957, 1964) in which C is the one-percentile and M the median of the grain size distribution, was used to distinguish the depositional environments by the criteria of competence and mode of sediment transport (Lespez et al., 2011). These parameters are indicators of hydraulic conditions under which sediments were transported. Four depositional environment types were characterized for the middle-to late-Holocene samples (Figure 2; Table 2).

CM image of depositional environments in middle to late Holocene and modern deposits of Ounjougou.
Sediment groups determined by grain size analysis and interpretation of the depositional environment (see Lespez et al., 2011).

(a) The coarsest sediments represent the channel deposits and are recorded only for the middle Holocene. They are composed of sandstone gravel and quartzose sand transported from the surrounding area to the river bed during high energy flood flow. (b) The laminated fine to coarse sands are interpreted as sand sheets deposited from the main channel onto the proximal floodplain during flood events. (c) The silty to fine sandy sediments rich in organic remains originated from floodplain units inundated with a lesser occurrence and velocity. This might be explained by the greater distance from the channel. (d) The dark organic silts, only recorded during HR1B (4650–4000 cal. BP), are characteristic of decantation processes in standing pools on the floodplain (Lespez et al., 2011).
Phytolith extraction, classification and analysis
15–20 g of dry sediment was processed according to the standard method described in Piperno (2006) and Neumann et al. (2009). Absolute phytolith counts range from 180 to 540 diagnostic morphotypes, depending on phytolith concentration and corrosion. Recent works have shown that prehistoric fires can be inferred from burned phytoliths (Boyd, 2002; Kealhofer, 1996; Morris et al., 2010; Piperno, 1994; Sangen et al., 2011; Schiegl et al., 2004). For each morphotype, we therefore differentiated dark opaque phytoliths from the light and coloured translucent ones in order to assess the impact of fire on the vegetation. Dark coloured but still translucent phytoliths are naturally present in some plant groups but opaque dark phytoliths result from burning of occluded organic material (Parr, 2006; Piperno, 2006). Sponge spicules, diatoms and charred fragments were also counted, but excluded from the sum, as well as the non-diagnostic morphotypes.
The classification described and illustrated in Figure 3 and Table 3 is based on Neumann et al. (2009), information from the literature and a reference collection of wood, leaf, bark, fruit and inflorescence phytoliths from modern Sahelo-Sudanian species, especially from the taxa represented in the charcoal samples. Diagnostic phytolith morphotypes were described using the ICPN classification (Madella et al., 2005) and classified into three main categories:
Sclereids and the globular morphotypes are attributed to woody and herbaceous dicotyledons (Figure 3A, B).
Grass short cell phytoliths (GSCP) are produced by Poaceae (Figure 3C). Only rarely can the morpho-types be related to plant species because of redundancy (the same morphotype occurs in different taxonomic groups) and multiplicity (several morphotypes occur in the same species) (Rovner, 1971). However, several studies have shown that phytoliths are particularly distinctive within the Poaceae and that an assignment can be made between phytoliths and grass subfamilies (Barboni and Bremond, 2009; Barboni et al., 2007; Bremond et al., 2005; Twiss et al., 1969). Bilobates and crosses occur mainly in the Panicoideae subfamily. The majority of panicoid grasses are adapted to a warm and humid climate. Chloridoideae, which produce saddles, are the dominant subfamily in the Sahelian zone today and adapted to a warm and dry climate. The subfamily Bambusoideae is characteristic of tropical warm zones. They produce diagnostic ‘collapsed saddles’ (Mercader et al., 2010; Piperno, 2006; Sangen et al., 2011). In our study, the bamboo could be identified as Oxytenanthera abyssinica, abundant in the charcoal samples (Eichhorn and Neumann, 2012). Absent from the Sahelian zone today, Oxytenanthera abyssinica is present along low energy rivers in the Sudano-Guinean zone. The rondel morphotype is diagnostic for Poaceae but not for a subfamily and is mainly produced in inflorescences. We included rondels and other grass short cell phytoliths into the undifferentiated Poaceae class.
Three family specific morphotypes are produced by non grass monocotyledons (Arecaceae, Commelinaceae and Cyperaceae), and one by the dicotyledonous Podostemaceae (Figure 3D). Palms (Arecaceae), producing the globular echinate morphotype, are widespread in West Africa today. Cyperaceae occur in wetlands but some species are also found in fields on the interfluves. The Commelinaceae are herbal species occurring in West Africa either in cultivated fields, along rivers, in the undergrowth of gallery forest (Floscopa, Murdannia) or on sandstone crusts (Cyanotis lanata) (Eichhorn et al., 2010). The Podostemaceae are riverweeds attached to rock surfaces in swiftly flowing water. They are a good indicator of fluvial activity (Piperno, 2006).

Phytolith morphotypes in Ounjougou samples. Scale bar 10 µm. Woody dicotyledons; (A) Sclereids; (B) Globular phytoliths; (C) Grass short cell phytoliths; (D) Family specific. (A): (a) cylindric faceted; (b) cylindric geniculate psilate; (c) irregular psilate; (d) polyhedral faceted; (e) irregular branched thick; (f) ellipsoid with outgrowths. (B): (a–d) decorated indiff.; (e) psilate; (f) subglobular dark core; (g–h) decorated compound. (C): (a–d) bilobate; (e–f) cross; (g) bilobate nodular; (h) trilobate; (i) polylobate; (j) Oxytenanthera saddle; (k) burned Oxytenanthera saddle; (l, m) squat saddle; (n, o) tall saddle; (p, q) rondel. (D): (a) Cyperaceae; (b) Podostemaceae; (c) globular echinate (Arecaceae); (d) burned globular echinate (Arecaceae); (e–h) Commelinaceae; (e) Cyanotis lanata type; (f) Commelina type; (g) Floscopa/Murdannia type; (h) Commelina forskaolii type.
Morphotype classes used in this study.
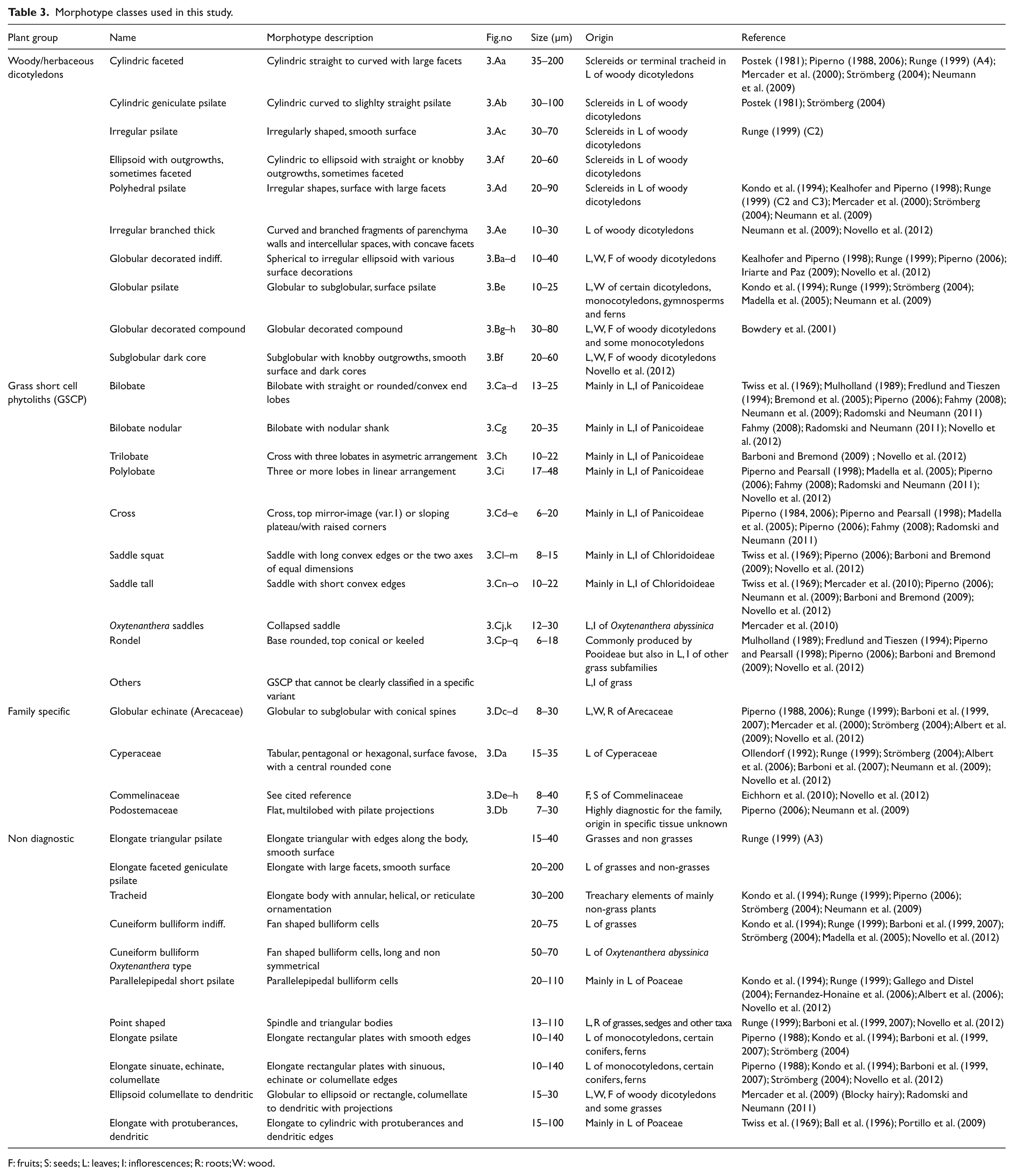
F: fruits; S: seeds; L: leaves; I: inflorescences; R: roots; W: wood.
In the analysis of the data, the index and general approaches were applied (Neumann et al., 2009). The indices describe the relationship of the vegetation with climatic parameters such as temperature and degree of aridity. In our study, the Iph and the D/P indices were calculated. The Iph index, first defined by Diester-Haass et al. (1973), is the ratio of Chloridoideae phytoliths (saddles) versus total Chloridoideae plus Panicoideae phytoliths (i.e. saddles plus lobates) and indicates the dominance of short-grass or long-grass savannas along an aridity–humidity gradient. According to Bremond et al. (2005), high Iph values (> 20 ± 1.4%) indicate vegetation dominated by Chloridoideae, i.e short-grass savanna, and point to dry climatic conditions whereas low Iph values (<20 ± 1.4%) indicate tall-grass savanna with dominating Panicoideae, suggesting humid climatic conditions or high available soil moisture. The D/P, first proposed by Alexandre et al. (1997), describes the tree cover versus open grass vegetation and is the ratio of globular decorated phytoliths, produced in woody dicotyledons, versus GSCP (Bremond et al., 2008). This index, already discussed by Strömberg (2004), cannot be applied for vegetation types including Bambusoideae, because their collapsed saddles represent relatively closed vegetation. In vegetation with Bambusoideae, the D/P underestimates the tree cover. Therefore, we excluded the Oxytenanthera saddles from the sum of the GSCP in the D/P calculation.
For a precise reconstruction of the paleovegetation at a local scale, the index approach is not sufficient. The general approach, including all morphotypes from all size classes, is generally considered to be restricted by redundancy and multiplicity. However, recent studies have recognized new morphotypes from plant taxa with clear ecological preferences that can be useful to characterize local vegetation changes and human impact (Eichhorn et al., 2010; Strömberg, 2004). Taxa with specific phytolith morpho-types such as Arecaceae, Cyperaceae, and Commelinaceae yield information on water availability, and can indicate pioneer and anthropogenic plant communities (Eichhorn et al., 2010).
Results and interpretation
Modern assemblages
Fourteen phytolith surface samples were studied from three transects along the Yamé valley at Ounjougou, Doucombo (20 km downstream), and Sampara (located at the confluence of the Yamé with the Niger) (Figure 4). Owing to the proximity to water, the tree cover is denser on the floodplain than on the interfluves, with their savannas strongly altered by humans.

(a) Phytolith percentage diagram from modern samples (in grey are the percentages of burned phytoliths). Abundances lower than 1% are figured by a dot; (b) transects and location of surface samples.
Channel deposits
Three samples originate from the Yamé river bed (2, 7, 12) but the size of the catchment area contributing to the sediment and water flow is wider at Sampara (4400 km2) and Doucombo (650 km2) than at Ounjougou (200 km2). They consist of detritic sediment generated by flows with high velocity and channel avulsions at the valley bottom during the period of intensive rainstorms in the wet season. The assemblages are dominated by phytoliths produced by Poaceae (50, 67 and 64%) while globulars and sclereids occur with lower values (40, 28 and 25%). Low D/P indices suggest relatively open vegetation. The other channel bed sample (5) comes from Ravin Sud, a small seasonal tributary of the Yamé with sediments originating from a small area on the sandstone plateau covered by a traditional park savanna with Butyrospermum parkii, Adansonia digitata and Borassus aethiopum. Owing to the more important tree cover, the phytolith assemblage has more globular decorated (40%) and Arecaceae phytoliths (12%) while GSCP reach only 30%. Thus, the D/P is relatively high (1.3).
Floodplain deposits
Owing to their topographic position, two types of floodplain deposits can be identified: one which is seasonally flooded (1 and 14), and one other, more elevated, only inundated during exceptional events (3, 6, 8, 11). The assemblages from the regularly inundated floodplain are similar to those from the channel and field deposits, probably because the sediment had been transported seasonally from the catchment area. The phytolith assemblages from the second type of floodplain differ markedly. Because of pedogenesis and low occurrence of flood flows, the sediments contain mainly phytoliths produced by the local vegetation. In these samples, phytoliths from woody dicotyledons and Arecaceae are over-represented (63–70%). Sclereids reach 20–25% and globular decorated fluctuate between 33 and 40% whereas Poaceae GSCP are weakly represented (24–32%). At Ounjougou, the presence of Oxytenanthera saddles can be explained by erosion of the neighbouring middle-Holocene sediments which are deposited on the floodplain.
Field sediments and pond deposits
Phytolith assemblages from millet fields (4, 9) and a pond (13) are dominated by grass phytoliths (58–71%). The most conspicuous feature of these samples is the high percentages (26–35%) of bilobates. They probably originate from pearl millet in the fields and from panicoid wetland species such as Echinochloa stagnina in the pond.
Dry soils on sandstone
Dry soils located on the sandstone interfluves were sampled at Doucombo (10). Owing to their skeletal soils, mainly taxa such as Balanites aegyptiaca and Combretum spp. are growing there and cultivation is not possible. The phytolith assemblage is characterized by a low percentage of phytoliths from woody dicotyledons (28%). Grass phytoliths reach a high value (66%), particularly the Chloridoideae saddles (29%), representing grasses adapted to poor soils and aridity.
Middle- to late-Holocene deposits: From phytolith assemblages to vegetation reconstruction
Several stratigraphic units have been sampled for the middle to late Holocene corresponding to different transport and deposition patterns and catchment area origin. Sediments from the Balanites and Detarium sections originate from the Yamé catchment area while the Ravin Sud deposits come from a smaller catchment area of a tributary of the Yamé. These factors have to be considered in the interpretation. Six phytolith assemblage zones (OUN-1 to OUN-6) have been recognized using constrained cluster analysis (CONISS) (Grimm, 1987) (Figure 5, Table 4).

Phytolith percentage diagram from middle- and late-Holocene deposits of Ounjougou (in grey are the percentages of burned phytoliths). Abundances lower than 1% and D/P index lower than 1 are figured by a dot.
Absolute counts of phytoliths in fossil and modern samples from Ounjougou.

Site identification: B: Balanites; D: Detarium; RS: Ravin sud; OUN: Ounjougou; DC: Doucombo; SP: Sampara.
OUN-1: From early to middle Holocene (before 7425–7261 cal. BP)
The oldest sample, from a floodplain deposit, has a high proportion of woody dicotyledons phytoliths, reaching 80%. The main feature is the high percentage of sclereids (67%) mainly represented by the polyhedral psilate morphotype (Table 4). The composition of the assemblage is similar to those from the early Holocene with high values of sclereids and low values of globular decorated phytoliths (Neumann et al., 2009). However, they are different in regard to their GSCP. Although the proportion of grass phytoliths is very low for the middle-Holocene sample (17%), the presence of Oxytenanthera saddles (12%) points to a distinct vegetation change compared with the early-Holocene samples in which this morphotype is absent. Similarly, the very low percentage of the other GSCP (5%) and a D/P of 2.3 indicate a closed gallery forest on the valley floor with Oxytenanthera abyssinica. The Iph is 25%.
OUN-2: The middle Holocene at Balanites/Detarium (7425–6000 cal. BP)
The most conspicuous feature of these samples is the increase of the Oxytenanthera saddles (27–47%). Generally, values of globular decorated are higher in comparison with the preceding period (around 20% versus 13%) and sclereids decrease (from 67 to 30% on average). Diagnostic GSCP (without Oxytenanthera saddles) show only low values around 10%. Phytoliths produced by woody dicotyledons are better represented in the floodplain deposits (49–70%) than in the channel deposits (29% and 52%) and the percentages of Oxytenanthera saddles are higher in the coarser deposits. Probably the bamboos were growing at the outer margins, about 10 to 100 m from the main channel. Today, Oxytenanthera abyssinica does not grow along rivers with high energy flow but rather along those with calm water. Chloridoideae saddles are also more abundant in the coarse deposits, but with low percentages. Family-specific morphotypes are not well represented; Commelinaceae and Cyperaceae phytoliths occur with low values (<1%). The D/P index shows lower values for the coarse deposits (2.3 and 0.7) while the assemblages from the floodplain deposits have D/P indices from 1.6 to 11.5. From the abundance of woody dicotyledons phytoliths and the high D/P, relatively closed vegetation can be inferred for the floodplain. The Iph index reaches percentage from 20 to 100% but cannot be used because of the low numbers of GSCP. Burned phytoliths account for 11–29% and occur with higher percentages in the finer layers. They are more common among sclereids and Oxytenanthera saddles, indicating that the local vegetation was affected by fires.
OUN-3: The middle Holocene at Ravin Sud (6200–5940 cal. BP)
Sclereids reach higher values in the fine layers (40%) than in the coarser sediments (14%). The proportion of the globular decorated (44%) and the Arecaceae type (8%) is more important in the channel deposits in comparison with the Detarium sediments but also with the floodplain deposits of the same section. This indicates that the vegetation in the Ravin Sud catchment area was more closed than that of the Yamé. Sclereids are clearly representative of the gallery forest while the higher values of globular decorated phytoliths in the coarse layers suggest that they had been produced by savanna trees. The values of Oxytenanthera saddles are higher in the floodplain deposits. The lower energy of the Ravin Sud stream in comparison with the Yamé can explain the better representation of Oxytenanthera saddles from the local vegetation. Other GSCP are almost absent from the phytolith assemblages, indicating a closed environment on the interfluves and the valley floor. The D/P indices of the two samples are very high (17.9 and 17.7) because there were only few GSCP except Oxytenanthera saddles. High numbers of burned sclereids and Oxytenanthera saddles indicate that fire occurred mainly in the vegetation of the floodplain.
OUN-4: The transition of the middle to late Holocene at Ravin Sud (5300–5060 cal. BP)
The phytolith assemblage is similar to that from the Detarium section (OUN-2). Sclereids are much more common in the floodplain deposits (31–32%) than in the coarser sediment (12%). Conversely, GSCP reach higher values in the coarse (17%) than in the finer deposits (2–7%). As indicated by the D/P indices (1.7 in channel sediments to 3.5 and 13 in floodplain deposits), the vegetation in the catchment area was more open probably because of regular bush fires. Indeed, burned phytoliths reach high values of 14% in the coarser sediment and 23–33% in the local deposits.
OUN-5: The beginning of the late Holocene at Balanites (5310–4220 cal. BP)
Although from the same period and the same sedimentation pattern as Ravin Sud, the assemblages indicate more open vegetation. Phytoliths produced by woody dicotyledons do not show significant differences between sandy and silty deposits. Most conspicuous are decreasing values of sclereids (around 10%) and higher values of GSCP in the sandy layer (32%), particularly bilobates (15%). GSCP values of 15 and 17% in the finer deposits indicate more open vegetation for the Yamé catchment area also on the valley floor. As for the preceding period, the D/P index is lower for the channel deposits (0.7) compared with the floodplain deposits (1.3 and 1.7). The values of burned phytoliths (7–10%) are decreasing.
OUN-6: The late Holocene at Ravin Sud and Balanites (4580–4220 cal. BP)
Panicoideae and Chloridoideae GSCP have significantly higher percentages than in the older samples. For the first time, bilobates reach 20%. Arecaceae, Cyperaceae and Commelinaceae phytoliths are also better represented. By contrast, Oxytenanthera saddles and undifferentiated GSCP phytoliths are less represented. Sclereids decrease and globular decorated stay stable. At the bottom of the Ravin Sud stratigraphy, the assemblages are more differentiated. Samples from the channel deposits have higher percentages of saddles and bilobates in comparison with the silty layer in between where globular decorated, Bambusoideae saddles and the Arecaceae morphotype are better represented.
The D/P values decrease in comparison with the preceding periods at Ravin Sud (OUN-4) and only reach 0.8–1.1 in the channel deposits. The Iph index is also low because of the increase of bilobates which might suggest Sudanian grasslands, but low D/P indices point to more open vegetation. Low percentages of burned phytolithes, around 10%, indicate that bush fires were less intensive in comparison with the middle Holocene, probably owing to a lower biomass production.
Discussion
Taphonomy of the phytoliths
Analysis of the modern samples shows that the taphonomy of the phytoliths has to be considered in the interpretation and reconstruction of vegetation dynamics. In contrast to pollen, the phytolith assemblages in soils result mainly from decomposition of the local vegetation. In the samples from cultivated fields, bilobates are over-represented, whereas at a distance of only 150 m, the samples from the sandstone interfluves are dominated by saddles, indicating poor and dry soils. In the alluvial deposits, the origin of phytoliths is more complex, depending on the type of transport and the area of origin. The channel and sand sheet deposits transported from the interfluves during the rainy season contain numerous phytoliths originating from the grasses in the surrounding savannas. By contrast, the finer sediments of the floodplain and the ponds deposited with low energy are dominated by phytoliths from the local vegetation.
The taphonomic model of modern deposition can also be applied to the fossil sediments. If assemblages from the same site and time period are compared with each other, samples from the channel and those from the floodplains and ponds can be clearly distinguished, just as the modern ones. Each sedimentary unit has its own phytolith signature, indicating the area of origin: the finer floodplain sediments contain remains of the gallery forest, whereas the assemblages in the coarser deposits from the channel and proximal floodplain represent a mixture of phytoliths from different vegetation types on the interfluves and also from the gallery forest.
The results show that the applied methodology, including geomorphological study of the deposits, is effective for characterizing different modes of vegetation representation by phytoliths, depending on the origin and type of deposition, and for a spatial reconstruction.
The use of the general and index approaches in fluvial records
Based on the taphonomic results, the two approaches can be discussed with regards to vegetation reconstruction and history of alluvial deposits. The indices Iph and D/P describe the relation of vegetation types and climatic parameters, such as temperature or degree of aridity (Alexandre et al., 1997; Barboni et al., 2007; Bremond et al., 2005, 2008). The D/P, here calculated by excluding the Bambusoideae morphotype, is a good tool for characterizing the woody plant cover. Higher D/P values have been found in modern and fossil samples from the floodplain, pointing to quite dense arboreal vegetation. Lower values in the channel samples indicate more open vegetation in the surrounding area. Consequently, the D/P can differentiate vegetation communities, such as gallery forest and savanna, on a local scale. It is thus essential to consider taphonomic processes and the possibility of local variability in palaeoecological reconstructions. This implies that for application of the D/P in transregional studies, only data from comparable sediments should be used.
The index Iph is mainly applied in Africa for the differentiation of bioclimatic zones. In the fossil Holocene samples, Iph values are extremely variable, fluctuating between 20% and 100% for the same sediments and time period. Values <20%, usually indicating long grass savannas with dominating Panicoideae under warm and humid climatic conditions, have mainly been found in the late-Holocene samples although a shift towards drier climate took place during this period (Kröpelin et al., 2008). In contrast, samples with values > 20%, pointing to grasslands dominated by Chloridoideae and hence the prevalence of warm and dry climate, are dated to the middle Holocene when climatic conditions were more humid. It can be concluded that the Iph should not be utilized for the reconstruction of different vegetation types on a regional scale in the fluvial record because it highly corresponds with the local vegetation. Moreover, elevated percentages of bilobates in the modern samples from cultivated fields and ponds might be attributed either to pearl millet (Pennisetum glaucum) and associated panicoid field weeds or to wetland grasses.In this case, high Iph values cannot be interpreted as representing Sudanian tall-grass savannas. This shows that the significance of the Iph as an indicator of rainfall is distinctly reduced in landscapes of tropical Africa where cultivation of panicoid cereals can be assumed. But consideration of depositional characters can help to resolve the difficulties in index interpretations.
The general approach, including all morphotypes, yields more precise information about particular vegetation communities (Blinnikov et al., 2002; Boyd et al., 1998; Runge, 1999; Strömberg, 2004). Our data show that certain morphotypes can be indicators of plant communities, especially the gallery forest in the river valley and savannas in the surrounding areas. Sclereids are better represented in the samples from modern and fossil floodplains, while the globular decorated morphotype is rather correlated with the surrounding savanna samples especially in the middle Holocene. In the fossil samples, Arecaceae globular echinate phytoliths are more abundant from the 6th millennium cal. BP onwards. Today, the most prominent palm species in the Yamé valley is Borassus aethiopum. According to Giffard (1967), Borassus aethiopum is a light-demanding species and at least as full grown individuals highly resistant to fire. In this case, the increase of globular echinate phytoliths can be linked to vegetation opening and increased fire activity. Commelinaceae and Cyperaceae are important elements of wetland communities but provide also information about human disturbance because they invade cultivated fields (Eichhorn et al., 2010) and form important components of secondary vegetation (Piperno, 1988). The presence and abundance of Oxytenanthera saddles in fossil assemblages can be useful to determine the proximity to a perennial water source. Oxytenanthera abyssinica is associated to calm and wet environments such as swampy floodplains and perennial ponds. In summary, it can be concluded that the general approach with a large number of different morphotypes in all size classes, in combination with the index approach, allows for a better understanding of small-scale vegetation dynamics (Strömberg, 2004).
Vegetation dynamics during the middle to late Holocene: Climatic change or human impact?
If phytolith data are linked with geomorphological processes, the vegetation at the middle- to late-Holocene transition (7790–4000 cal. BP
During this period, wetlands with hygrophilous vegetation developed in Ounjougou in a regional context of dense savannas and woodlands (Eichhorn and Neumann, 2012). The presence of well-watered environments generally corresponds to the ‘Holocene Humid Optimum’ in northern tropical Africa leading to the ‘Green Sahara’ and distinctly higher lake-levels from the Sahara to the Guinean Zone (Ballouche and Neumann, 1995; Gasse, 2000; Hoelzmann et al., 2004; Kröpelin et al., 2008; Salzmann and Waller, 1998; Salzmann et al., 2002; Servant, 1983), and also locally in the Yamé Valley (Lespez et al., 2011). At Ounjougou, the phytoliths indicate closed vegetation at the valley bottom and also on the interfluves. Oxytenanthera abyssinica, appearing in the middle-Holocene phytolith and charcoal samples, points to a major vegetation change, mainly regulated by climatic factors. In the sediments dated to 7248–5060 cal. BP, around 20% of the phytoliths are burned, especially those from the gallery forest, among them sclereids and Oxytenanthera saddles. It is difficult to find natural causes of regular burning events in a hygrophilous gallery forest; anthropogenic fires for hunting or better access to the water provide a much more plausible explanation. In addition, the presence of humans at Ounjougou is attested by a lithic industry between the 8th and 6th millennium BP (Kouti and Huysecom, 2007).
At the beginning of the late Holocene (HR1A, 5320–4650 cal. BP), the vegetation became more diversified in the Yamé catchment area. In the two sites of this period (Balanites and Ravin Sud) the samples from the channel deposits are comparable but those of the floodplains show differences. Higher values of sclereids in the samples from Ravin Sud point to a denser woody vegetation, while higher GSCP percentages in the Yamé River valley samples indicate more open environments, most probably due to local conditions. However, for the second part of the late Holocene (HR1B, 4650–4000 cal. BP) vegetation transformation is visible all over the Yamé catchment area, attested by higher percentages of GSCP (especially bilobates), Commelinaceae and Cyperaceae and lower D/P values. The opening of the vegetation at Ounjougou after 4580 cal. BP can be correlated to a shift to drier conditions in West Africa. This aridification trend is also recorded between 4700 and 3300 cal. BP in several lakes of the Nigerian Manga Grasslands (Salzmann and Waller, 1998; Wang et al., 2008). Decreasing sclereids in the floodplain deposits might be correlated to a change in the gallery forest where Sudano-Guinean taxa were gradually decreasing (Eichhorn and Neumann, 2012).
The late Holocene is also a period of major cultural change and the emergence of agriculture in West Africa, first evidenced by the appearance of domesticated pearl millet (Pennisetum glaucum) dated to 4569–4416 cal. BP in the Tilemsi Valley, 250 km north of Ounjougou (Manning et al., 2011). From 4500 cal. BP onwards, increasing aridity and the dwindling of suitable grazing grounds and available water resources forced Saharan pastoralists to migrate southwards into the Sahel (Breunig and Neumann, 2002; Hassan, 2002; Kuper and Kröpelin, 2006). At Ounjougou several sites of the Late Neolithic indicate more intensive settlement activities in the course of the 5th millennium cal. BP (Ozainne et al., 2009). Pottery of Saharan affinity indicates that Ounjougou was situated along the southward migration way from the Tilemsi Valley (Ozainne et al., 2009). Probably these populations brought the knowledge of millet cultivation with them. The increase of bilobates in the channel deposits, originated from the surrounding areas and dated to 4580–4290 cal. BP, can be explained as first indirect evidence of pearl millet cultivation at Ounjougou, several hundred years before the appearance of its charred grains in the archaeological record around 3700–3500 cal. BP (Ozainne et al., 2009).
Conclusions
The combination of the general and the indices approaches (mainly the D/P index) coupled with geomorphological studies is a good tool to reconstruct the vegetation in fluvial and anthropogenic environments and offers new perspectives for areas lacking other bio-proxies. Even if in situ decay is the dominant factor for the depositional pattern of the phytoliths, an influx from the regional vegetation can be recorded in the coarser deposits. At Ounjougou, general patterns of the middle- to late-Holocene vegetation dynamics were already known by previous anthracological studies. Phytolith analysis confirms the openness of the environment around the 5th millennium BP through higher percentage of GSCP and lower D/P values. The most important contribution of phytolith analysis at Ounjougou is the detection of human impact on landscapes already during the middle Holocene. There is a continuous fire record for the gallery forest from the 7th millennium cal. BP and evidence for agriculture from 4500 cal. BP.
Footnotes
Acknowledgements
We thank our Malian colleagues for the support of our work in the field and the members of the project ‘Human populations and paleoenvironment in West Africa’ for good and helpful cooperation, especially Eric Huysecom, Aziz Ballouche and Yann Le Drézen. We are grateful to Jennifer Markwirth for processing the phytolith samples and Richard Byer for language editing.
Funding
This study is part of the ANR-DFG project ‘Archéologie du paysage en Pays Dogon’ (APPD) and of the PhD Dissertation of Aline Garnier funded by Région Basse-Normandie.