
Abstract
Madagascar is home to some of the world’s most unique plants and animals. Unfortunately, anthropogenic forest loss has had a dramatic impact on both floral and faunal communities. In many regions the scale and timing of this loss remains poorly constrained. Such is the case for northwestern Madagascar. Pollen records for this region suggest that fires and grass abundance increased around 1000 calendar years before present (cal. BP), as well as c. 500 cal. BP, presumably reflecting growing human settlements. However, temporal and spatial resolution for these transitions is limited by a hiatus in the sediment record. We present a suite of carbon isotope data from 14C-dated vertebrate bones from Anjohibe Cave, northwestern Madagascar. In agreement with palynological records, isotope data indicate that the transition to open savannah may be a recent phenomenon in northwestern Madagascar. We document dramatic changes in the vertebrate community over the past 2000 years. Remains from extinct taxa are all older than 1500 cal. BP, locally extirpated taxa were in the region until c. 300 years ago, and introduced species are essentially modern. Stable carbon data suggest minimal consumption of C4 resources by extinct, locally extirpated and locally extant endemic taxa. On the contrary, introduced rats and shrews exhibit exceptionally high carbon isotope values; this may reflect a dietary shift to newly introduced grasses or consumption of seeds from C4 plants.
Introduction
It was once believed that Madagascar was densely forested prior to human arrival (e.g. Humbert, 1927; Perrier de la Bathie, 1921, 1936). However, this impression has been increasingly challenged (Bond et al., 2008; Burney, 1987; Koechlin, 1972). Savannahs may be a natural part of the island’s vegetation. Nevertheless, humans have clearly impacted forested environments, and recent forest loss through slash-and-burn agriculture has been substantial (Green and Sussman, 1990; Harper et al., 2007). Without an accurate picture of long-term local vegetation change, it is difficult to assess the degree to which modern grasslands are anthropogenically derived, or predict the future impacts of current habitat transformation.
Madagascar is recognised as one of the world’s highest-priority ‘hotspots’ for biodiversity (Myers et al., 2000). Most of its species are endemic to the island (Goodman and Benstead, 2005). Subfossils, named for their geologically young ages, provide a glimpse of the past history of modern taxa. Subfossil assemblages contain many extinct species, suggesting that modern diversity has been reduced relative to that of the recent past (Samonds et al., 2012). Madagascar suffered a late-Holocene extinction event that removed all of its large (>10 kg) native animals (Burney, 1999; MacPhee, 1986; MacPhee and Marx, 1997). While the cause of this extinction is debated, both climate change and human impacts have been suggested (e.g. Burney et al., 2004; Crowley, 2010; Godfrey and Irwin, 2007; Virah-Sawmy et al., 2009).
Our understanding of Holocene climate and vegetation shifts on Madagascar is based largely on lake sediment records (Gasse and Van Campo, 1998; Matsumoto and Burney, 1994; Virah-Sawmy et al., 2009). Around 5000 years before present (yr BP), a brief but intense drought is recorded in central and southeastern Madagascan sediments (Gasse and Van Campo, 1998; Virah-Sawmy et al., 2009). Prolonged aridity commenced around 3000 BP (Burney, 1993), peaking c. 1000 BP (Matsumoto and Burney, 1994; Virah-Sawmy et al., 2009). Vegetation changes associated with this shifting climate were mostly regional (Burney, 1987, 1993; Gasse and Van Campo, 1998; Matsumoto and Burney, 1994; Virah-Sawmy et al., 2009). However, an island-wide increase in grass pollen between 1300 and 950 BP is evident (Burney, 1987, 1993; Matsumoto and Burney, 1994; Virah-Sawmy et al., 2009). This roughly coincides with the first discernable human presence in most regions (Burney, 1987; Burney et al., 1997; Dewar and Richard, 2012; Dewar and Wright, 1993; Virah-Sawmy et al., 2009).
Northwestern Madagascar preserves some of the earliest large-scale human settlements on the island (Dewar and Wright, 1993). Today, the vegetation is predominantly palm savanna with isolated patches of riparian forest (Koechlin, 1972; Matsumoto and Burney, 1994). Pollen spectra previously collected from speleothems at Anjohibe Cave (Figure 1) indicate that regional vegetation has been similar for at least 40,000 years (Burney et al., 1997). Nevertheless, this area has undergone extensive human modification, and was at least partially forested in the recent past (Burney et al., 1997; Harper et al., 2007). Charcoal, especially from graminoid sources, increases dramatically both in speleothem surface accretions and surficial cave sediments (Burney et al., 1997). Ages for these deposits are poorly constrained, but radiocarbon and uranium-series dating indicate they are late Holocene. Pollen records from Lake Mitsinjo (16.03°S, 45.83°E), 125 km to the southwest, indicate that a woodland mosaic was present from the mid Holocene until c. 1000 BP (Matsumoto and Burney, 1994). Grass pollen and charcoal both increase in lake sediments c. 1000 BP, and again around 500 BP (Matsumoto and Burney, 1994). Unfortunately, a hiatus in sediment deposition coincident with the inferred vegetation shift c. 1000 BP complicates constraining the timing and extent of this shift (Matsumoto and Burney, 1994). Here we use carbon isotope values from radiocarbon-dated bones to investigate if and when the vertebrate community at Anjohibe Cave was affected by a late-Holocene increase in C4 grasses.
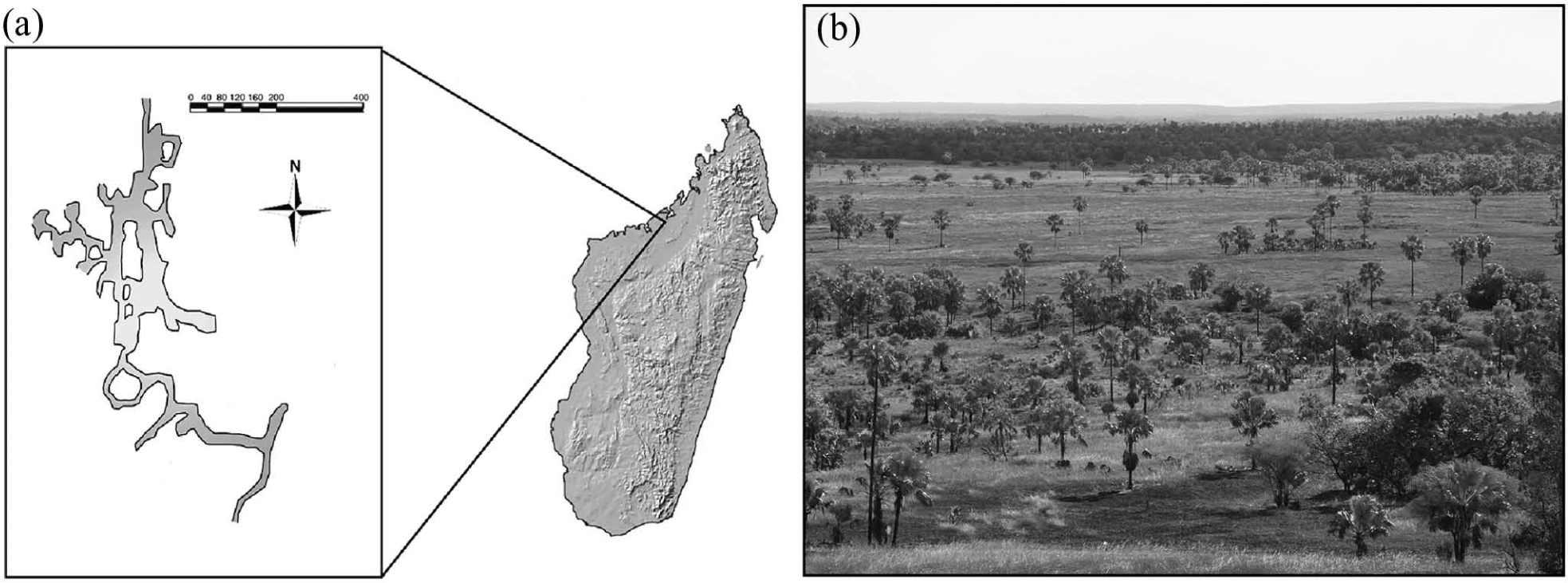
(a) Map of Anjohibe Cave, northwestern Madagascar (adapted from Samonds, 2007). Map scale in meters. (b) Palm savannah photograph taken from the cave entrance.
Background on stable isotopes
Stable isotope ratios are presented as δ values, where δ = [(Rsample/ Rstandard)−1] × 1000, R=13C/12C, and the standard is Pee Dee Belemnite. Values are presented as parts per thousand (‰) deviations from the standard. Stable carbon (δ13C) isotope values in an animal’s bones reflect an individual’s long-term diet, for example the type of plants consumed. Most terrestrial plants rely on either C3 or C4 photosynthesis. Whereas most trees, shrubs and herbs are C3, most tropical grasses are C4 plants (Bond et al., 2008). Endemic succulents are relatively uncommon in northwestern Madagascar (www.tropicos.org, 2012). Physiological differences in how CO2 is converted to sugar and where carbon is fixed within a leaf result in isotopic differences between C3 and C4 plants (reviewed in Marshall et al., 2007). Reported δ13C ranges for C3 and C4 plants are −20 to −37‰ and −12 to −16‰, respectively (Kohn, 2010; Marshall et al., 2007). These isotopic differences are passed on to animal consumers. Collagen δ13C values in herbivores are roughly 5‰ higher than those in consumed plants, and faunivores may have slightly higher δ13C values (c. 1–2‰) than sympatric herbivores (e.g. Bocherens et al., 1995; Fox-Dobbs et al., 2007; Hyodo et al., 2010).
Site description
Anjohibe Cave (15.55°S, 46.89°E, Figure 1) is located in the southern part of the Mahavo Plains dry lowland savannah in northwestern Madagascar (Decary, 1934). Anjohibe is a large karst system formed in an Eocene limestone plateau (Besairie, 1969; Burney et al., 1997; de Saint-Ours and Paulian, 1953). Remains from more than 60 species of higher vertebrates have been previously reported at the cave (Burney et al., 1997; Godfrey and Jungers, 2003; MacPhee, 1986; Ravoavy, 1986; Samonds, 2007; Samonds et al., 2010). Ages for these animals range from modern to about 80,000 BP (Samonds, 2007). In addition to endemic species still found in the region today, extirpated and extinct taxa have also been recovered. The majority of these species were likely forest dependent, suggesting that their disappearance is at least partly correlated with a reduction in forest habitat near the cave (Burney et al., 1997; Green and Sussman, 1990). Remains from introduced rats (Rattus sp.) and shrews (Suncus murinus) are also present.
Methods
Sample collection and preparation
We collected bones from loose surface sediments, shallow pools, and accumulations near entrances, beneath skylights, or in sunken forests where the cave ceiling collapsed. Comparative measurements for identification were made with 500-172 Mitutoyo digital calipers to 0.01 mm, or with a MA285 Meihi Techno optical reticule X, 1/100 (0.01 mm).
We isolated collagen from bones by decalcification in EDTA, gelatinization in weak HCl, and filtration through 1.5 µm glass-fiber filters (outlined in Crowley, 2012). Samples were analysed using a Thermo-Electron (Finnigan) Delta XP Continuous Flow System connected to an Elemental Analyser at the UC Santa Cruz Stable Isotope Laboratory. Precision based on nine IAEA Acetanilide standards was 0.1‰. Collagen preservation is relatively poor at Anjohibe. Nearly one-third of the bones we attempted to analyse yielded no collagen or atomic C:N ratios far outside of the acceptable range (2.9–3.6). This was particularly true for tortoise (Aldabrachelys spp.) scutes. None of the tortoise specimens we attempted to analyze yielded collagen. We excluded these poorly preserved specimens from further analysis.
Forty-nine specimens yielded acceptable collagen data. Published radiocarbon dates exist for some of these specimens (Samonds et al., 2010). Previously undated specimens were sent to the Center for Accelerator Mass Spectrometry at Lawrence Livermore National Laboratory. Most, but not all, of these bones yielded 14C dates. We augmented this data set with published radiocarbon and δ13C values from Burney et al. (1997). We converted conventional radiocarbon years (14C BP) to calendar years (cal. BP) in Calib 5.01 using a 20 yr moving average and the Southern Hemisphere calibration curve SHCal (McCormac et al., 2004; Stuiver and Reimer, 1993). We used mean calibrated ages in all statistical analyses.
For all subfossils older than 150 cal. BP, we corrected δ13C values by −1.2‰ to account for post-industrial isotopic shifts in atmospheric CO2 (following Crowley et al., 2011a). Global radiocarbon levels were artificially increased by nuclear testing in the 1950s (Reimer et al., 2004). ‘modern’ specimens (0 14C BP) may, therefore, be up to 60 calendar years old. We assumed that specimens that yielded acceptable C:N ratios did not contain enough carbon to measure 14C ages are >50 years old and corrected their δ13C values accordingly. We refer only to corrected carbon isotope data throughout the remainder of this paper.
Quantitative analyses
Based on the timing of potential vegetation shifts, we divided our specimens into two time bins: Modern–1000 cal. BP, and >1000 cal. BP. As this division excluded specimens that did not yield dates, we also divided our specimens into four groups: Extinct, Locally Extirpated, Locally Extant, and Introduced. Extinct taxa include the hippo Hippopotamus madagascariensis, the lemur Archaeolemur cf. edwardsi and the elephant bird Mullerornis sp. Locally extirpated animals include fruit bats, the nile crocodile (Crocodylus niloticus; C Brochu, personal communication), and several species of lemur that are currently restricted to eastern Madagascar. Locally extant taxa include tenrecs, a turtle, and several species of lemur and bats. Introduced species include rats, shrews, and a giant frog (Hoplobatrachus cf. tigrinus). All of these taxa are thought to have arrived over the past two millennia (Samonds et al., 2013; Vences et al., 2003). We recognize that our extinct taxa are not fully representative of the diversity of organisms that once inhabited the region surrounding Anjohibe Cave. Remains from extinct elephant birds are rare in northwestern Madagascar. Fragmented eggshell (attributed to Mullerornis sp.) has been recovered from Lavakasaka, a rockshelter that lies roughly 5 km southwest of Anjohibe (Burney et al., 1997). Stable carbon isotope data are not available for this material. To the best of our knowledge, this is the full extent of elephant bird material that has been found in the region. Remains from extinct carnivorans are also rare. The extinct tortoise (Aldabrachelys spp.) specimens that we sample did not yield collagen.
We do not have stable isotope data for plants from Anjohibe. However, the mean δ13C value for C3 plants collected at Ankarafantsika National Park, which lies 100 km south of Anjohibe, is −28.1 ± 1.4‰ (Crowley et al., 2011b). Carbon isotope values have not been measured for C4 plants from this region. We assume that they are similar to the global average (−14‰; Marshall et al., 2007). Using a fractionation of c. 5‰ between collagen and plants (Koch, 1998; Krueger and Sullivan, 1984), animals with pure C3 diets at Anjohibe should, on average, have δ13C values ≤ −23.1‰, whereas animals consuming some C4 foods should have higher δ13C values. The highest δ13C value recorded for a C3 plant at Ankarafantsika is −25.6‰ (Crowley et al., 2011b). We therefore infer that subfossil individuals with corrected collagen δ13C values higher than −20.6‰ must have consumed some C4 resources (direct consumption of C4 plants or insects, or consumption of arthropods or small vertebrates that consumed C4 plants).
All statistical tests were performed using JMP 5.0.1. We tested for homogeneity of variances using Bartlett tests for equal variances. When variances were equal, we used student t-tests and one-way Analysis of Variance coupled with posthoc Tukey Honestly Significant Difference (HSD) tests. When variances were unequal, we used Welch t-tests. All probability levels are two-tailed, and the significance for all tests was set at α < 0.05.
Results
All data are presented in Table 1. Comparing animals older and younger than 1000 cal. BP, we find a significant difference in δ13C values. Animals >1000 cal. BP have significantly lower δ13C values than younger specimens (student t=5.366, d.f.=37, p<0.0001). The vast majority of individuals in this older group have δ13C values <−23.1‰ and all have δ13C values <−20.6‰. Conversely, all individuals in the younger group have δ13C values >−23.1‰, and many have δ13C values >−20.6‰.
Radiocarbon and δ13C data for all specimens included in this study. Radiocarbon sources: 1: Samonds et al. (2010); 2: this paper; 3: Burney et al. (1997). Institution codes: UA and UAP: University of Antananarivo, Madagascar; UMASS: University of Massachusetts, Amherst. Field/Acquisition numbers are included when available for uncatalogued specimens. Dietary Categories: H: Herbivore; F: Faunivore; O: Trophic Omnivore.

Specimen from Anjohikely, a nearby cave which is associated with the same cave system (15.33°S, 46.53°E). Based on the continued presence of turtles in the area, we have called this individual ‘locally extant’.
Uncorrected δ13C values.
Carbon isotope values corrected −1.2‰ to account for post-industrial isotopic shifts in atmospheric CO2.
Previously unpublished carbon isotope value provided by David Burney.
Eggshell from Lavakasaka, a rock shelter which is associated with the same cave system (15.57°S, 46.85°E).
Extinct animals are all >1500 cal. BP (Table 1). These dates are much older than those for the locally extirpated and locally extant fauna (Figure 2). Remains for locally extirpated fauna have 14C ages <500 cal. BP. Crocodylus niloticus is 385 ±75 cal. BP and fruit bats (Eidolon dupreanum) are c. 300 cal. BP. Neither of the bamboo lemurs (Prolemur simus) specimens yielded 14C dates. With the exception of one Eulemur fulvus (UMASS A29.2 N2 Left Branch), all dates for locally extant animals are historical (<500 cal. BP). All introduced taxa (Hoplobatrachus cf. tigrinus, Rattus sp., Suncus murinus) are essentially modern (Table 1).

Boxplots comparing 14C ages for four groups. Ages are calendar years BP.
Comparing δ13C values in introduced, locally extant, locally extirpated and extinct taxa, we find significant differences among groups (F3,46=42.084, p<0.0001). Posthoc tests indicate that extinct taxa have the lowest δ13C values, locally extirpated and extant taxa have intermediate values, and introduced taxa have uniquely high δ13C values (Figure 3). Extinct taxa consumed minimal C4 resources. Most individuals exhibit δ13C values that are <−23.1‰, and all are <−20.6‰ (Table 1). Locally extirpated taxa also consumed minimal amounts of C4; only the crocodile has δ13C values >−20.6‰. All Eidolon dupreanum have δ13C values <−20.6‰, and Prolemur simus have δ13C values <−23.1‰. Locally extant taxa consumed moderate amounts of C4 resources. Most individuals have δ13C values >−23.1‰. Only two Eulemur fulvus have δ13C values <−23.1‰. These include the uncatalogued individual mentioned above (UMASS A29.2 N2 Left Branch), and an undated individual (UMASS 12/8/96) which conceivably lived prior to 1000 BP. Horseshoe bats (Hipposideros sp.), tenrecs (Tenrec ecaudatus), and the unidentified turtle have δ13C values >−20.6‰ (Table 1). All of the introduced taxa have δ13C values that are substantially >−20.6‰. These individuals must have consumed some C4 resources.

Boxplots comparing δ13C values for four groups. Groups that share the same superscript letter are statistically indistinguishable (Tukey HSD; α = 0.05).
Discussion and conclusion
Radiocarbon and stable carbon data reveal a picture of faunal turnover and vegetation change at Anjohibe Cave. These data provide some important insights into the recent paleoecology of northwestern Madagascar.
Whereas all dated extinct taxa are >1500 cal. BP, the majority of dated subfossils from locally extant and locally extirpated fauna are <1000 cal. BP. While it is possible that this pattern reflects a taphonomic bias, we believe that this is unlikely for two reasons. First, many small animal bones are found in caves; in fact the oldest published date for Madagascar’s subfossil record is a bat from Anjohibe (Samonds, 2007). Instead, this phenomenon probably reflects the fact that Madagascar’s large and charismatic ‘megafauna’ have historically been the focus of paleontological research. Smaller associated species have only been identified and described relatively recently (e.g. Muldoon, 2010; Muldoon et al., 2009; Samonds, 2007; Samonds et al., 2010). Second, the disappearance of large-bodied taxa c. 1500 cal. yr BP is not a local phenomenon. Radiocarbon dates suggest that between 1000 and 2000 years ago, populations of endemic taxa >10 kg experienced a dramatic decline across the entire island (Crowley, 2010).
Animals living in the vicinity of the cave consumed negligible amounts of C4 prior to 1000 cal. BP, and endemic mammals continue to eat only small amounts of C4. Stable carbon data indicate that all extinct taxa included in this study relied on C3 resources, including the dwarf Hippopotamus madagascariensis, which presumably was a grazer. Although none of the tortoise remains from Anjohibe yielded collagen, we were able to obtain data from a carapace attributed to the extinct Aldabrachelys spp. from Amparihingidro, a marshy locality approximately 40 km southwest of Anjohibe. This individual yielded a radiocarbon age of 7135 ± 135 cal. BP and a corrected δ13C value of −23.2‰, indicating consumption of C3 plants. These results are consistent with previously published isotopic data from southern and southwestern Madagascar, where δ13C values for extinct elephant birds, tortoises, hippopotamuses and most lemur species indicate minimal reliance on C4 resources (Clarke et al., 2006; Crowley and Godfrey, 2013). Only one extinct species, the lemur Hadropithecus stenognathus has δ13C values consistent with habitual consumption of C4 resources. Remains from this species have not been recovered in Northwest Madagascar.
Locally extant and extirpated individuals from Anjohibe Cave exhibit variable δ13C values, indicating negligible to moderate consumption of C4 resources. Herbivorous lemurs Propithecus coquereli, Eulemur fulvus, and the fruit bat Eidolon dupreanum incorporated nominal amounts of C4 into their diets. However, several faunivorous taxa have δ13C values >−20.6‰. This pattern may result from trophic enrichment in δ13C values. Faunivores have slightly higher δ13C values than sympatric herbivores (c. 1‰) in many ecosystems (e.g. Bocherens et al., 1995; Fox-Dobbs et al., 2007; Hyodo et al., 2010). Additionally, this pattern may reflect the bias in collagen δ13C values towards dietary protein (Jim et al., 2004). Whereas most, if not all, dietary protein for browsers and frugivores derives from C3 trees and shrubs, dietary protein for faunivores derives from arthropods and small vertebrates that could feed on a mix of C3 and C4 resources. The omnivorous mouse lemur (Microcebus sp.) and insectivorous Tenrec ecaudatus have δ13C values that are comparable with the more herbivorous lemurs (Table 1). However, the crocodile, turtle, and two of the three horseshoe bats (Hipposideros spp.) have slightly higher δ13C values than the herbivorous taxa. Nevertheless, collagen δ13C values for these taxa still suggest predominantly C3-based diets.
Introduced taxa, including rats (Rattus sp.), shrews (Suncus murinus), and the giant frog Hoplobatrachus cf. tigrinus have remarkably high δ13C values, implicating substantial ingestion of C4 foods (Table 1, Figure 3). Isotope data suggest that these taxa are exploiting a distinctive niche that differs from any of the native taxa, extinct or extant. Increased δ13C values may reflect consumption of a new dietary resource that did not previously exist, such as an introduced C4 grass species. Alternatively, the opportunistically granivorous rats and shrews may selectively consume seeds from C4 grasses that have been in the region for thousands of years.
Combined, these data suggest that open savannah dominated by C4 plants is a relatively recent phenomenon at Anjohibe. A reduction of forest cover or an increase in C4 grasses at the expense of C3 grasses, such as bamboo, could both explain the observed patterns in δ13C values. We are not suggesting that regional vegetation was once dense forest, nor are we arguing that the transition to open savannah was dramatic. Indeed, the prevalence of native C4 species attests to the antiquity of C4 grasses in Madagascar (Bond et al., 2008), and pollen records at Anjohibe and nearby Lake Mitsinjo indicate that graminoids have been present for thousands of years (Burney et al., 1997; Matsumoto and Burney, 1994). Nevertheless, the presence of forest-dwelling extinct and locally extirpated fauna strongly suggests that the region surrounding the cave was more wooded than it is now, possibly as recently as 300 years ago. In addition to the species we successfully analyzed, remains from extinct arboreal sloth lemurs Palaeopropithecus kelyus, and Babakotia radofilai have been reported from Anjohibe (Burney et al., 1997; Godfrey and Jungers, 2003). Remains from the locally extirpated euplurid carnivoran Galidia elegans were also reported from Anjohibe (Samonds et al., 2010). This species is currently restricted to more forested regions of Madagascar (Goodman, 2003, 2012).
Our results are also consistent with the documented late-Holocene increases in charcoal and grass pollen at Anjohibe and nearby Lake Mitsinjo (Burney et al., 1997; Matsumoto and Burney, 1994). Radiocarbon dates suggest that populations of the large, extinct fauna may have been in decline prior to the inferred expansion of C4 grasses c. 1000 cal. BP (Crowley, 2010; Matsumoto and Burney, 1994). However, because of a hiatus in sedimentation, the timing of this vegetation change in the sediment record is not well constrained. It is possible that C4 grass abundance was on the rise prior to 1000 cal. BP. Increases in the abundance of C4 grasses during the Holocene may have corresponded to the regional decline of locally extirpated species. Although we were unsuccessful in obtaining 14C dates for bamboo lemurs (Prolemur simus), fruit bats (Eidolon dupreanum) and Crocodylus niloticus were still present 300 years ago. Today, P. simus is restricted to forested regions in eastern Madagascar (Mittermeier et al., 2010). Eidolon dupreanum is still widely distributed across the island, but it is susceptible to human pressures, particularly hunting (Jenkins and Racey, 2008). Endemic lemurs and tenrecs that remain in the region tend to concentrate in small isolated patches of woodland (Samonds, personal observation).
In agreement with previous studies, stable isotope data reveal details of vegetation and faunal turnover in Northwestern Madagascar. C4 grasses have increased, forest dwelling species have dwindled, and introduced taxa are exploiting a distinctive niche. This paleoecological perspective will have useful implications for predicting how Madagascar’s vertebrate community will respond to future climate change and anthropogenic impacts.
Footnotes
Acknowledgements
We thank LR Godfrey, SM Goodman and an anonymous reviewer for providing feedback on earlier versions of this paper. Fieldwork was performed under two collaborative agreements between the Département de Paléontologie et d’Anthropologie Biologique, Université d’Antananarivo and (1) DA Burney, LR Godfrey and WL Jungers, and (2) K Samonds. Thanks to H Andriamialison, A Rasomiaramanana, and the late G Randria, for the opportunity to collaborate. Thanks also to the Madagascar Institut pour la Conservations des Ecosystèmes Tropicaux, D Andreasen, S Conway, M Cravens, M Getty, T Hughes, J Karr, P Koch, M Loewen, and N Teakle.
Funding
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.