
Abstract
In rich fens, unlike bogs, the key drivers structuring testate amoeba communities are related to nutrient status, suggesting the potential for transfer functions to quantitatively reconstruct changing nutrient status from palaeoecological records. Such records could be useful tools to investigate the long-term impacts of pollution and landscape change. Here, we derive and test transfer functions for pH, water-table depth, conductivity and Ca and Mg concentrations using a data set from Polish fens. Results show that transfer functions for Ca and conductivity have apparent predictive power for surface samples; these models will require further validation and testing with palaeoecological data. Testate amoeba transfer functions for fen nutrient status may be a valuable addition to the peatland palaeoecologist’s tool-kit, although further work will be required to demonstrate their usefulness in practice.
Introduction
In the last two decades, most peatland palaeoenvironment studies have focused on reconstructing Holocene climate change (Gałka et al., 2013; Swindles et al., 2009); however, reconstructing the trophic state of wetlands is also an important aim (Hájkova et al., 2012). Shifts in peatland nutrient status may be anthropogenic through pollution (Payne et al., 2012) or catchment land-use change (e.g. deforestation); autogenic (Zobel, 1988), for instance, through ombrotrophication; or related to climatic change (Ammann, 1986). However, methods for quantitative reconstruction are currently undeveloped.
Testate amoebae are protists that are widely used in palaeoecological research (Mitchell et al., 2008) because they form shells that are routinely preserved in peats (Charman, 2001). In bogs, a key control on testate amoeba communities is depth to the water-table (DWT), and testate amoebae have been widely used in climate reconstruction (Charman et al., 2002; Warner, 1990). However, compared to bogs, rich fen testate amoeba ecology has been much less intensively investigated. Recent studies in the Czech Republic (Opravilova and Hajek, 2006), the eastern Mediterranean (Payne, 2011) and Poland (Lamentowicz et al., 2011) have described species–environment relationships. Studies in Poland show that, unlike bogs, conductivity, calcium and pH are the most important environmental controls on testate amoeba communities. This suggests the potential for nutrient status–related transfer functions (TFs), but the only study to previously attempt this used a very small local training set (Dudova et al., 2013). TFs for phosphorous have been applied in lakes (Patterson et al., 2012; Roe et al., 2010) and demonstrate the potential applications of similar models in fens. In this study, we aim to (1) produce TFs for nutrient status–related factors based on a previously derived data set, (2) test TFs by cross-validation and (3) apply TFs to an existing high-resolution peat profile from N Poland (Lamentowicz et al., 2013).
Materials and methods
Eight fens in western Poland (Wielkopolska region; Figure 1) were sampled in 8–22 locations per site (n = 147). Vascular vegetation of the studied fens was composed mainly of sedges and rushes with Schoenoplectus tabernaemontani, Cladium mariscus and Carex rostrata dominant. Brown mosses were also common, including Calliergonella cuspidata and Calliergon giganteum and the relict species Tomenthypnum nitens and Paludella squarrosa; Sphagnum fallax and Sphagnum angustifolium were recorded in some acidic habitats. The data set we analyse here adds four further sites to the data previously discussed by Lamentowicz et al. (2011). Full details of sampling and analytical methods, and discussion of the autecology of species are contained in this paper.
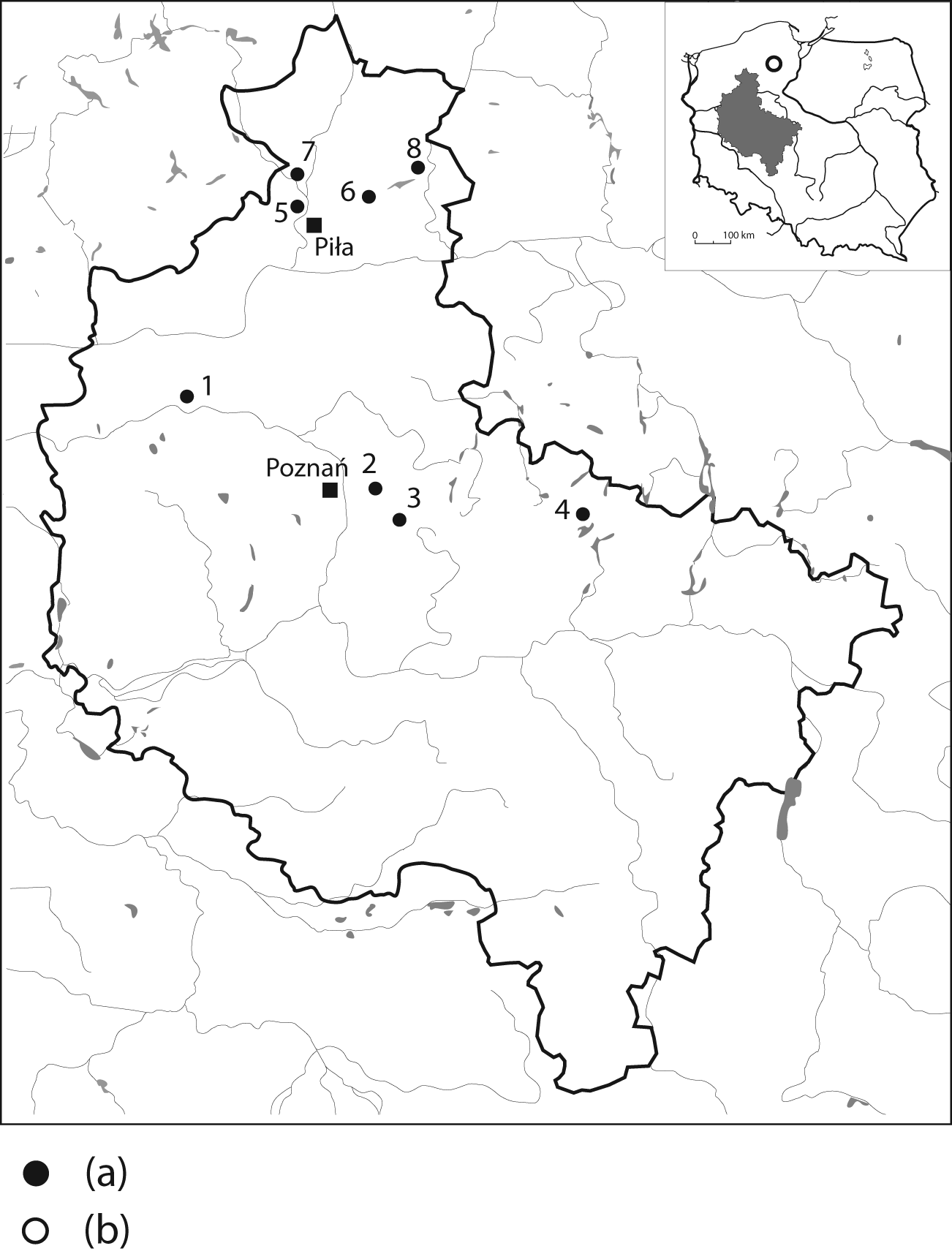
Location of the study sites: (a) transfer function and (b) peat sampling (1: Makąty; 2: Kazanie; 3: Wagowo; 4: Czarne; 5: Kuźnik Olsowy i Kuźnik Bagienny; 6: Czarci Staw; 7: Rurzyca and 8: Wierzchołek).
The upper 5 cm of mosses were removed and agitated in water with the fraction >20 µm and <300 µm retained for analysis (Booth et al., 2010). A total of 150 tests were identified per sample (Payne and Mitchell, 2009) based on the established taxonomic literature (Clarke, 2003; Decloitre, 1978, 1979; Grospietsh, 1958; Mazei and Tsyganov, 2006; Ogden, 1980; Ogden and Fairman, 1979; Ogden and Hedley, 1980). Three replicate groundwater subsamples were collected in the field and analysed as described in Lamentowicz et al. (2011).
Five variables explaining most variance in the species data are considered here: DWT, pH and conductivity measured in the field and Ca and Mg concentrations by atomic absorption spectrometry (AAS) in the laboratory (Hermanowicz et al., 1999). A 1-m core was extracted from the northern part of the main Stążka Basin using a Wardenaar sampler (Wardenaar, 1987) and subsampled every centimetre. Detailed description of the core and multiproxy results are provided in Lamentowicz et al. (2013).
TFs were developed using two established methods which are known to perform well with testate amoeba data (Mitchell et al., 2013; Payne et al., 2011): Weighted averaging (WA; Ter Braak and Barendregt, 1986) and weighted average partial least squares (WA-PLS; Birks, 1998). Recently, Juggins and Birks (2012) have introduced a new method which is tested here for the first time with testate amoeba data. Locally weighted–weighted averaging (LWWA) selects a local training set of size k using the distance criterion of the modern analogue technique (MAT) and then applies weighted averaging to this subset. In this case, we selected k = 30 after initial trials and use squared chord distance. TF performance was assessed using leave-one-out (LOO; also termed jack-knifing) boot-strap and leave-one-site-out (LOSO) cross-validation, recently suggested as a more robust approach to account for within-site clustering of samples (Payne et al., 2012). To account for the possible impact of unevenly sampled gradients, we also applied the segment-wise root mean square error of prediction (RMSEP) approach advocated by Telford and Birks (2011a). TFs were applied to the palaeodata set, and significance testing carried out using random TF (Telford and Birks, 2011b). All analyses were carried out in R 12.2.1 with the packages rioja (Juggins, 2012) and palaeoSig (Telford, 2013).
Results and discussion
Results show that TFs for water-table depth (WTD) and Mg have very little predictive power (Table 1): with LOO cross-validation R2 values are <0.3, whereas when the more conservative LOSO cross-validation is applied, R2 is <0.15 and RMSEP is greater than standard deviation (SD). This result for water-table contrasts with many studies in bogs and reinforces the fundamental differences in testate amoeba ecology between bogs and fens (Payne, 2011).
Performance of transfer functions for the different variables using various cross-validation methods. Best performing variables shown in bold and best results in terms of predictive power have grey background.
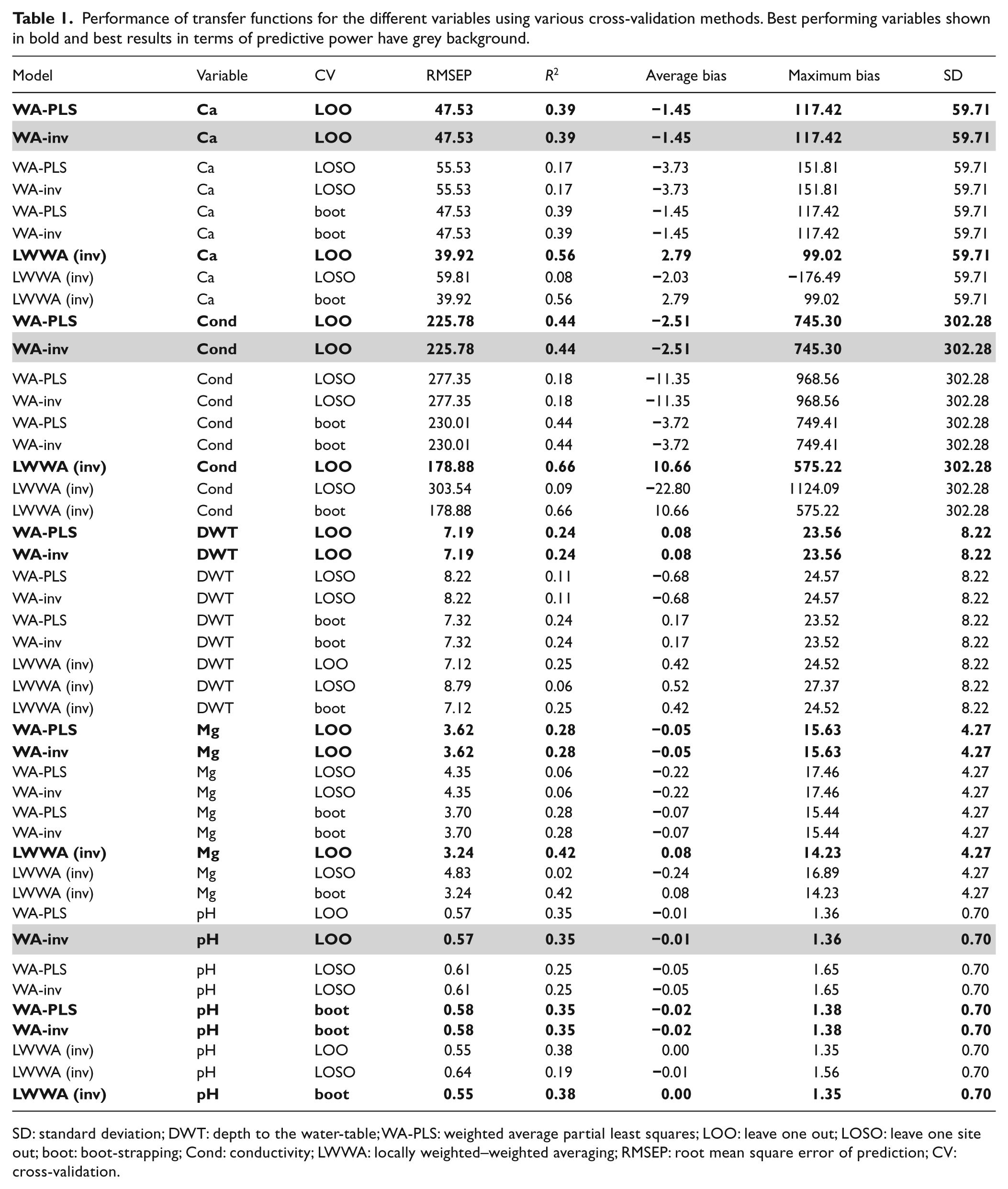
SD: standard deviation; DWT: depth to the water-table; WA-PLS: weighted average partial least squares; LOO: leave one out; LOSO: leave one site out; boot: boot-strapping; Cond: conductivity; LWWA: locally weighted–weighted averaging; RMSEP: root mean square error of prediction; CV: cross-validation.
Results were more promising in the cases of Ca, pH and conductivity (Table 1). In all these cases, multiple WA-PLS components did not improve on WA (inverse deshrinking). LWWA showed initial promise with standard cross-validation methods; however, when LOSO was used to cross-validate, the models lost all predictive power (R2 < 0.1 and RMSEP > SD). This is almost certainly due to the clustering problem discussed by Payne et al. (2012): in LOO and boot-strapping, it is likely that the majority of the 30 preferentially selected analogues will be from the same site as the test sample(s) and therefore model performance will be over-estimated. This is likely to be a general problem for LWWA in peatland TF studies. Due to these problems, we selected WA (inverse deshrinking) as the preferred model for pH, Ca and conductivity.
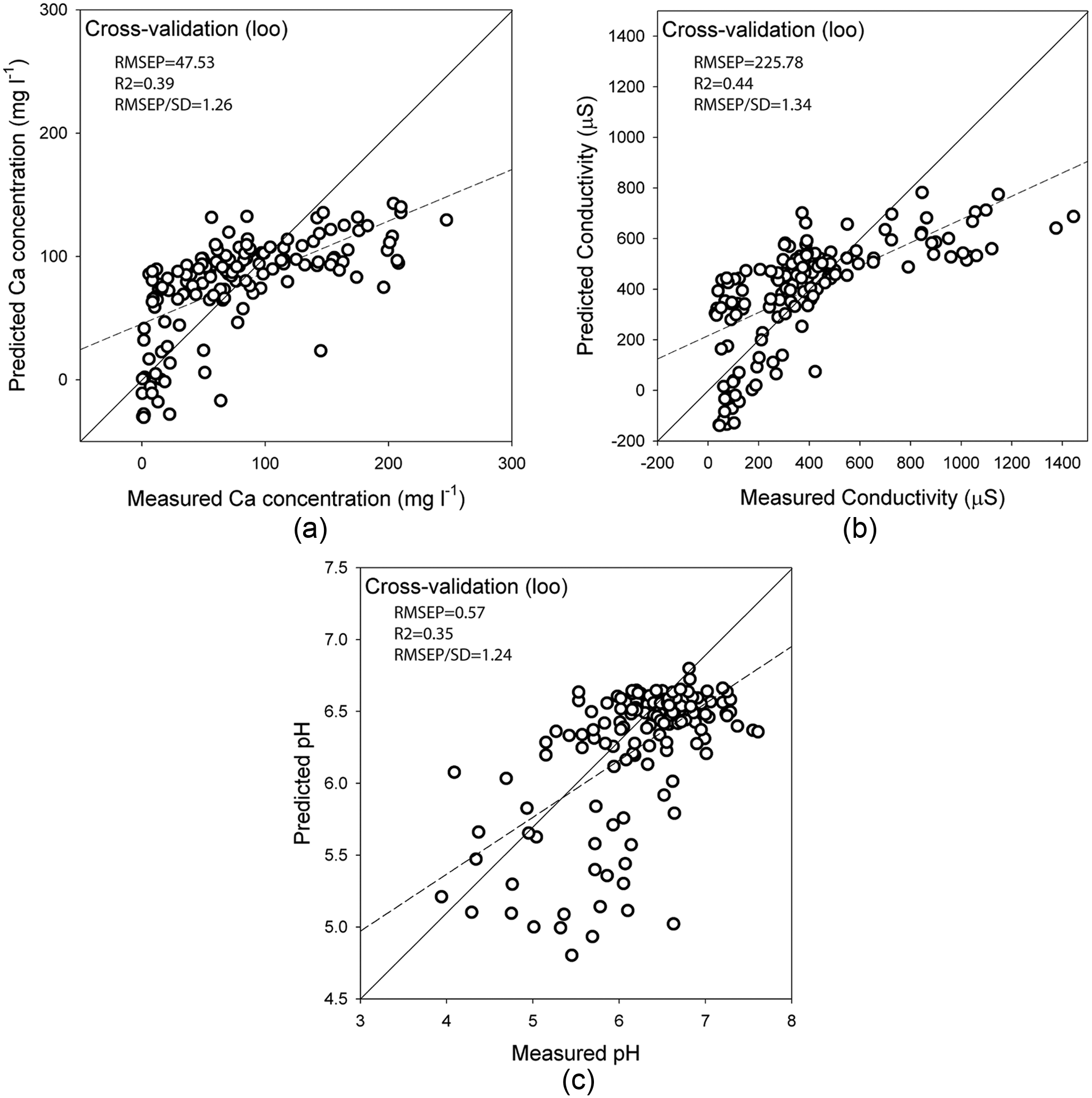
Performance of the transfer function of the particular variables: (a) Ca, (b) COND and (c) pH.
Reconstructions of pH, Ca and conductivity show very similar downcore trends (Figure 2). This is unsurprising as all represent the nutrient status/base-richness gradient, and there are strong correlations in the training set. Reconstructions support our previous qualitative interpretations (Lamentowicz et al., 2013) particularly in relation to deforestation and subsequent conifer afforestation in 1850–1900 (Giętkowski, 2011). However, when reconstruction significance is tested using the random TF approach, non-significant results are returned for all variables (Figure 3). While such a non-significant result does not mean that our interpretation of the record is incorrect, it does call into question the usefulness of the TF output; this appears to be a general issue with testate amoeba TFs and will be discussed more elsewhere.
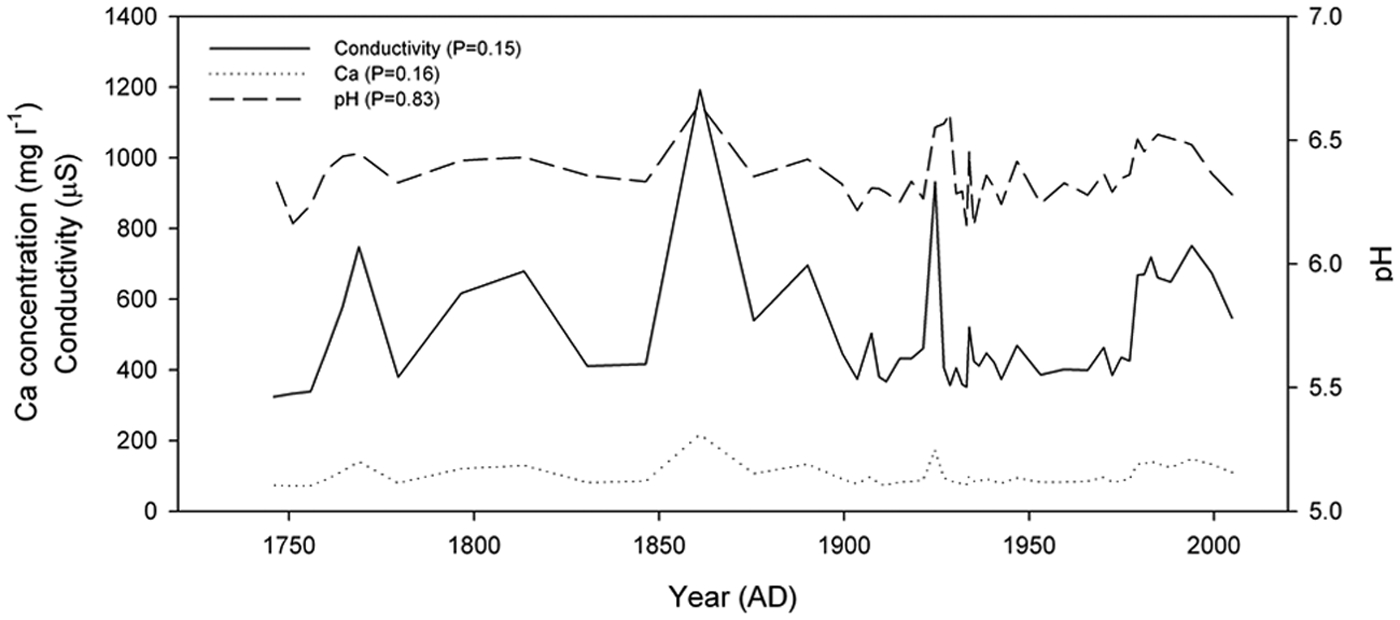
Quantitative reconstruction of Ca, pH and COND from Bagna nad Stążką peat core published by Lamentowicz et al. (2013).
Overall, our results show that TFs for conductivity and Ca concentration appear to have predictive power for surface samples in fens, while addressing the usefulness of such TFs in palaeoreconstruction will require further research. We believe that testate amoeba TFs for fen nutrient status may be a useful addition to the peatland palaeoecologist’s tool-kit.
Footnotes
Acknowledgements
We are grateful to Ralf Meisterfeld for the help in identification of the difficult testate amoeba taxa. We thank to Maciej Gąbka for the fruitful discussion and cooperation.
Funding
This study was financed by a scientific grant from the Polish National Science Centre (NCN; No. 305077936, Principal Investigator: Łukasz Lamentowicz). Financial support was also received from the grant PSPB-013/2010 from Switzerland through the Swiss Contribution to the enlarged European Union.