
Abstract
We report an intertidal, bone-rich peat deposit on the windward (Atlantic Ocean) coast of Abaco, The Bahamas. The age of the Gilpin Point peat (c. 950–900 cal. yr BP) is based on five overlapping radiocarbon dates (one each from single pieces of wood of buttonwood Conocarpus erectus and sabal palm Sabal palmetto, and single bones of the Cuban crocodile Crocodylus rhombifer, Albury’s tortoise Chelonoidis alburyorum, and green turtle Chelonia mydas). The short time interval represented by the charcoal-rich peat suggests rapid sedimentation following initial anthropogenic fires on Abaco. The site’s diverse snail assemblage is dominated by terrestrial and freshwater species. The peat is exposed today only during exceptionally low tides, suggesting a lower sea level at the time of deposition as well as a degrading shoreline during the past millennium. Fossils from Gilpin Point represent a late-Holocene vertebrate community at the time of first human presence; only 10 of the 17 identified species of amphibians, reptiles, birds, and mammals still live on Abaco. Numerous unhealed bite marks on the inside of the thick carapaces of the green turtle attest to consumption by Cuban crocodiles, which probably scavenged turtles butchered by humans. This concept, along with the dense concentration of bones in the peat, and charring on some bones of the green turtle and Abaco tortoise, suggests a cultural origin of the bone deposit at Gilpin Point, where the only Amerindian artifact recovered thus far is a shell bead.
Introduction
The late-Quaternary record of vertebrates on West Indian islands originates mainly from two very different sources of sediment. The first (and often richest) source occurs in karst features (dry or flooded limestone caves, rockshelters, and sinkholes) that accumulate bones most often through non-cultural means such as natural trap activity or predator roosts (e.g. Pregill, 1981; Pregill et al., 1994; Steadman et al., 2007). The second source is from open cultural (archaeological) sites where bones were deposited by prehistoric peoples (e.g. Jones O’Day, 2002; Steadman et al., 1984). In this article, we describe an unusual sedimentary setting for the deposition of ancient bones on a West Indian island, namely, a coastal peat deposit discovered on Great Abaco Island in the northern Bahamas. This peat has yielded an intriguing set of molluskan and vertebrate fossils that was deposited rapidly when humans first arrived in the northern Bahamas.
Site setting
The Bahamas consists of oceanic islands that lie just off the eastern coast of Florida (Figure 1). The archipelago features 23 major islands and many smaller ones lying on shallow carbonate banks separated by deep water. All exposed bedrock in this tectonically stable archipelago are Quaternary eolianitic subtidal limestone (Hearty and Kaufman, 2000; Mylroie, 2008). The Bahamian Archipelago stretches 980 km from c. 27°N and 79°W in the northwest to c. 21°N and 71°W in the southeast and consists politically of the independent Commonwealth of the Bahamas (or ‘The Bahamas’) and the Turks and Caicos Islands, a British Protectorate.
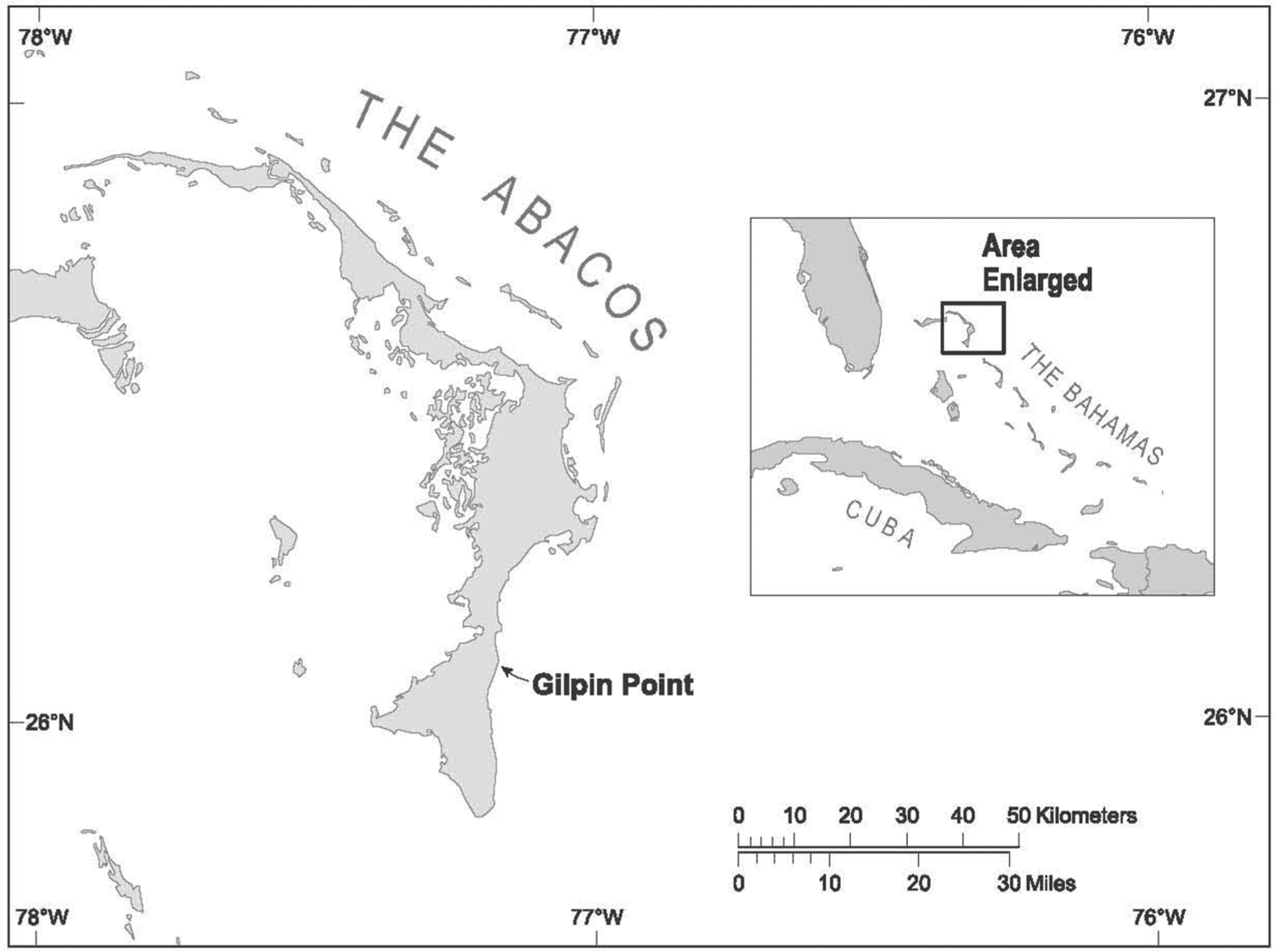
The Bahamian islands, showing the location of Abaco and other islands mentioned in the text.
Our study focuses on Great Abaco Island (hereafter ‘Abaco’), the second largest Bahamian island (1214 km2). Gilpin Point is an area of irregular limestone outcrops and calcareous beach sand on the windward (eastern, Atlantic Ocean) coast of southern Abaco (26.10457°N, 77.177666°W, Datum WGS84; Figure 2). The Gilpin Point site was discovered by Sabrina Bethel and landowner Perry Maillis on 8 and 10 May 2009, during a very low spring tide associated with a full moon on 9 May 2009. The dark, peaty sediment at Gilpin Point is inundated today by the ocean under normal circumstances, as well as covered by calcareous sand. Access to the sediment typically is possible only during very low tides (spring tides) associated with certain new and full moons during non-summer months (October–May). Even then, only the most exceptional spring tides are low enough to expose the sediment subaerially. PM also collected bones on 6 June 2009; 21, 22, 27, and 30 November 2009; 5 December 2009; sporadically from 28 September to 2 December 2012; 30 March 2013; and 10 May 2013. During our group’s field work on 8 and 13 November 2012 (new moon on 13 November), the deposit remained under c. 10 cm of water even during the lowest spring tide. During another round of group field work on 1 December 2012 (full moon on 28 November), the deposit was exposed subaerially for 30 min during the lowest spring tide (Figure 3).
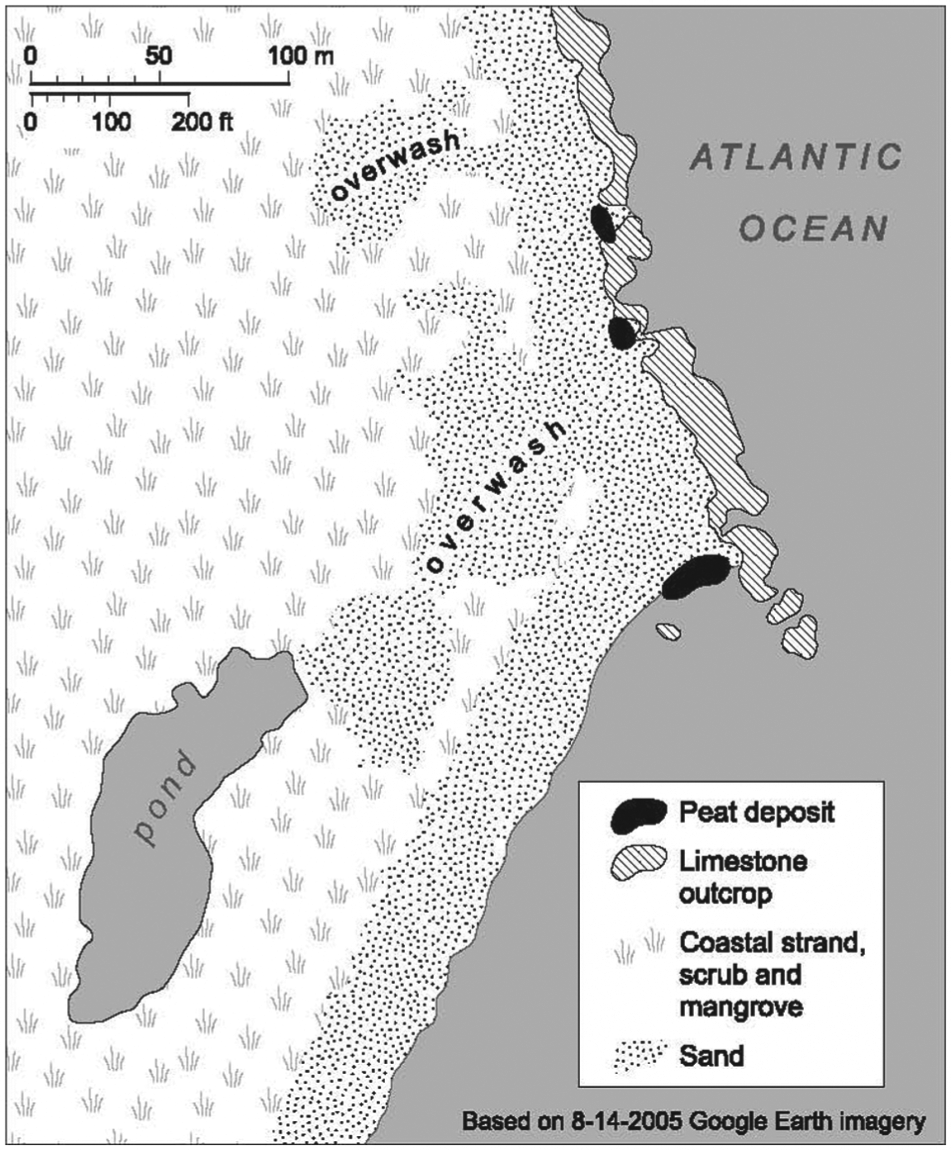
Gilpin Point, Abaco, The Bahamas, showing the location of the bone-bearing peat deposit.

Peat deposit exposed, 1 December 2012, Gilpin Point, Abaco, The Bahamas. DWS and PM pictured.
Aside from tidal cycles, another factor that limits access to the peat deposit is sand transport. For example, during our visit to Gilpin Point on 10 June 2013 (new moon on 8 June), the site was covered by sand at least 1 m deep. Prevailing easterly winds off the Atlantic Ocean tend to build up sand during the warm months, as well as inhibit extremely low spring tides. During the cooler months, when Abaco receives north winds from periodic frontal systems, sand is more often removed from this section of shoreline, thereby exposing the peat layer during extremely low spring tides. The peat can be exposed and then completely covered by sand over a single tidal cycle.
Methods
Because the peat deposit faces the open ocean on a high-energy coast, it would be prohibitively complicated and expensive to build a break-wall to improve access to the site. Therefore, our collection methods at Gilpin Point consisted of removing blocks of the peat (working quickly because of the narrow time interval of the lowest tidal level) with a square shovel (spade; Figure 4). These blocks of sediment, each overlain and underlain by calcareous sand, measured from 14 to 20 cm wide, 10 to 16 cm long, and 10 to 22 cm deep (c. 1.5–6 L per sample). For the three largest species of vertebrates recovered (Crocodylus rhombifer, Chelonoidis alburyorum, Chelonia mydas), c. 60% of the fossils were collected in situ because of being partially exposed at the top of the peat. Fossils of the other, smaller species were collected by screen-washing c. 0.2 m3 (200 L) of sediment. We processed c. 50% of this sediment in the field through nested sieves of 12.5, 6.4, and 1.6 mm mesh; the remainder of the sediment was processed in the laboratory through five nested sieves of 12.5, 5.6, 3.35, 2.0, and 1.0 mm mesh. Snails and small bones occurred throughout the vertical sequence of sediment.

DWS collecting blocks of peat with a square spade during the lowest spring tide, 1 December 2012, Gilpin Point, Abaco, The Bahamas.
The fossils from Gilpin Point are housed in the scientific collections of the National Museum of the Bahamas in Marsh Harbour, Abaco, under the care of NAA. Some of the fossils are currently on loan to DWS at the Florida Museum of Natural History, University of Florida (UF). Identifications of the fossils are based on direct comparisons with modern specimens of mollusks and vertebrates in the UF collections. In ‘Results’ section, we provide the common, scientific, and family name for each species at its first mention.
Results
Sediment description
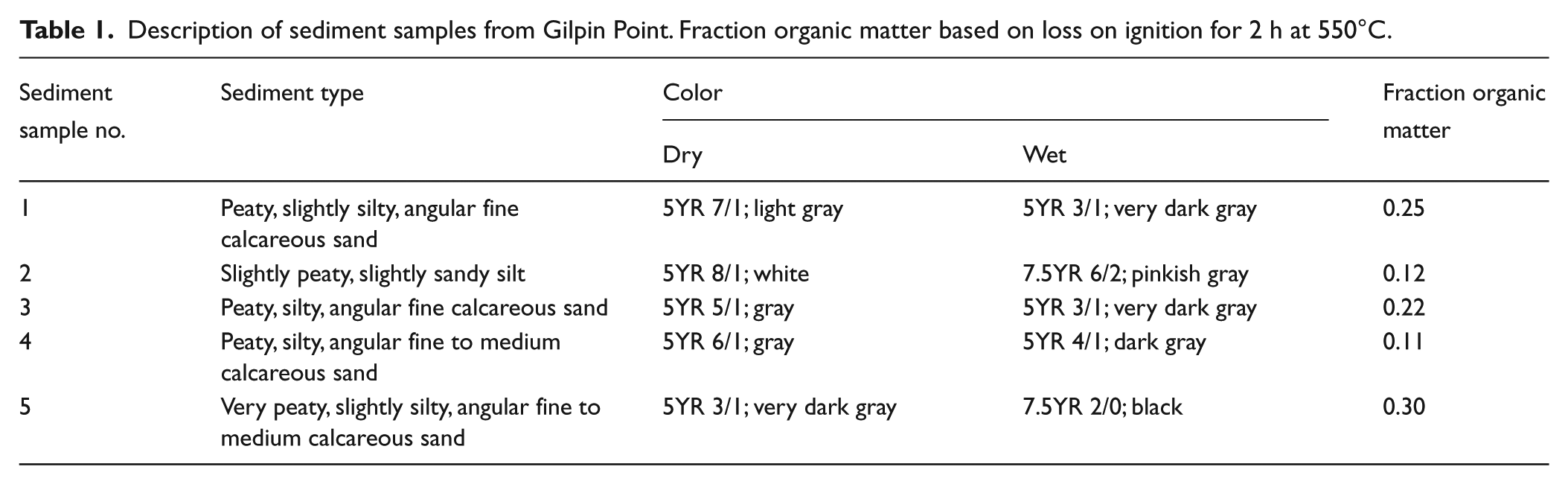
The sediment at Gilpin Point is poorly stratified, variably silty, and sandy peat (Table 1). The organic matter consists of charcoal (pieces ranging in size from microscopic to 14 mm diameter), unburned plant material, and bones. The peat deposit is 20–22 cm thick. We found no vertical trends in any characteristics of the peat. The peat deposit is much less organic and has considerably fewer mollusks and bones in the two small outcrops to the north of the main collection area, which is the largest and southernmost black area in Figure 2. All five sediment samples are from the main collection area; the two that are the least organic (samples 2 and 4) are from the northern margin of the main collection area. We did not quantify the charcoal other than to point out that it dominates the organic matter combusted to generate the loss on ignition (LOI) data (Table 1).The charcoal occurred throughout the deposit; we found no rich concentrations that would suggest a cultural feature such as a hearth. The deposit contained no high-energy overwash sediments of marine origin.
Description of sediment samples from Gilpin Point. Fraction organic matter based on loss on ignition for 2 h at 550°C.
Chronology
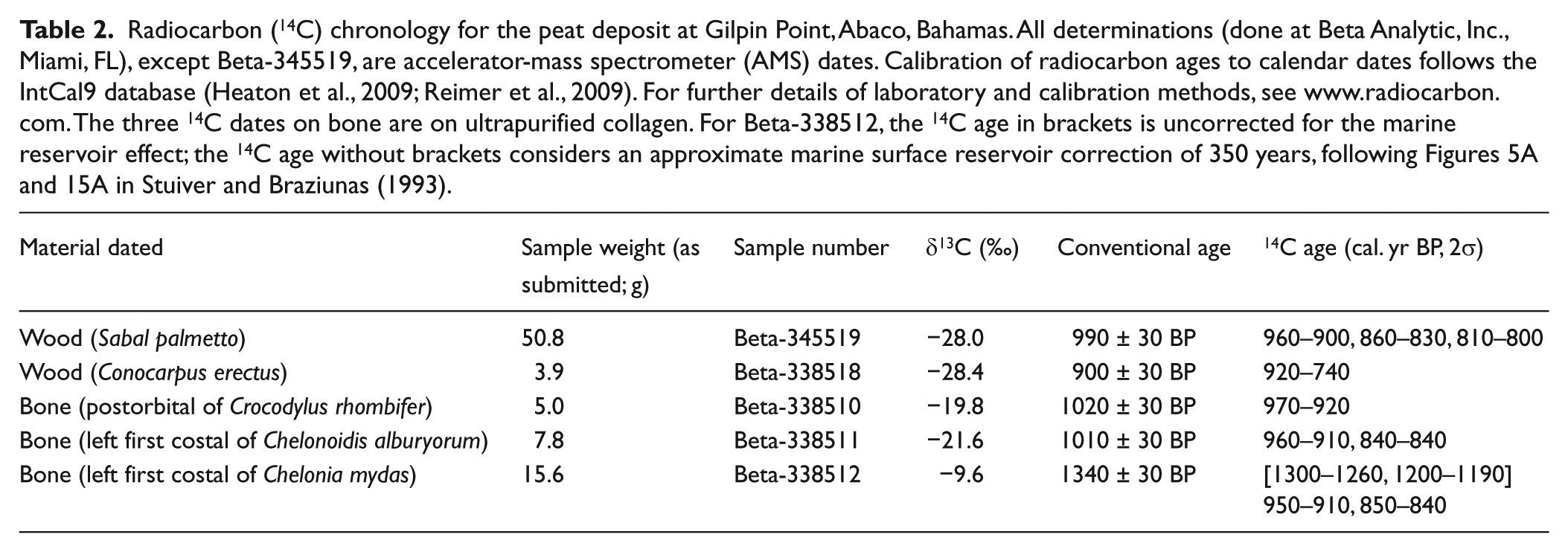
Our age estimate for the peat deposit at Gilpin Point is based on five radiocarbon (14C) dates from individual specimens of plants and animals identified to species (Table 2). The four accelerator-mass spectrometer (AMS) 14C dates consist of one on wood of the buttonwood Conocarpus erectus (Combretaceae) and three on purified collagen from individual bones of the Cuban crocodile Crocodylus rhombifer (Crocodylidae), the extinct Albury’s tortoise Chelonoidis alburyorum (Testudinidae), and green turtle Chelonia mydas (Cheloniidae). The single conventional 14C date is from a large sample of wood of the sabal palm Sabal palmetto (Arecaceae). The two 14C dates on wood overlap (at 2σ) at 920–900, 860–830, 810–800 cal. yr BP, whereas the three 14C dates on bone overlap (at 2σ) at 950–920 cal. yr BP. Both the buttonwood and palm were found in an upright growth position (Figure 5), suggesting that they may have continued to live after deposition of the bones. This mechanism would explain the slightly younger age determinations for the wood samples than the bone samples.
Radiocarbon (14C) chronology for the peat deposit at Gilpin Point, Abaco, Bahamas. All determinations (done at Beta Analytic, Inc., Miami, FL), except Beta-345519, are accelerator-mass spectrometer (AMS) dates. Calibration of radiocarbon ages to calendar dates follows the IntCal9 database (Heaton et al., 2009; Reimer et al., 2009). For further details of laboratory and calibration methods, see www.radiocarbon.com. The three 14C dates on bone are on ultrapurified collagen. For Beta-338512, the 14C age in brackets is uncorrected for the marine reservoir effect; the 14C age without brackets considers an approximate marine surface reservoir correction of 350 years, following Figures 5A and 15A in Stuiver and Braziunas (1993).

Black trunks of Conocarpus erectus in growth position in water on left side, originating in peat deposit, 13 November 2012, Gilpin Point, Abaco, The Bahamas.
With a 14C age of 970–920 cal. yr BP, Beta-338510 is the youngest direct age determination for the Cuban crocodile in the Bahamas and therefore helps to pinpoint the time of its extirpation on Abaco. The earliest 14C age previously determined directly on a Cuban crocodile bone in the Bahamas was 2880–2750 cal. yr BP (Beta-225508) from Sawmill Sink blue hole, Abaco (Steadman et al., 2007). (A blue hole is a flooded cave or sinkhole.) Nevertheless, crocodile bite marks exist on an Albury’s tortoise specimen from Sawmill Sink that has a 14C age of 970–920 cal. yr BP (Beta-298219; Hastings et al., in press), which is identical to our age determination for the crocodile from Gilpin Point. With a 14C age of 960–910 and 840–840 cal. yr BP (Beta-338511), the Albury’s tortoise from Gilpin Point overlaps with the two youngest previous age determinations for this species, namely, 970–920 cal. yr BP (Beta-298219; Sawmill Sink) and 920–780 cal. yr BP (Beta-298220; Lost Reel Cave, Abaco; Hastings et al., in press).
The ‘marine reservoir effect’ reflects the fact that marine organisms and ocean waters are depleted in 14C (i.e. contain ‘dead’ carbon) and therefore may yield 14C ages that are older than those based on contemporary terrestrial organic materials (Goodwin, 1993). The precise range of the surface marine reservoir correction for the Bahamas is not known, so we used an approximation of 350 years (from global estimates in Stuiver and Braziunas, 1993) to correct the AMS 14C date for the green turtle.
Stable isotopes
The carbon stable isotope data (δ13C values) of the 14C-dated samples (Table 2) are reasonable for each species, given what is known about their biology. Like most woody plants, the buttonwood and sabal palm follow the C3 (Calvin) photosynthetic pathway; their determined δ13C values (−28.0‰ and −28.4‰, respectively) are as expected for C3 plants (average −28.5‰, with a typical range of −37‰ to −20‰; Kohn, 2010; O’Leary, 1981, 1988).
The Cuban crocodile was a terrestrial carnivore in the Bahamas (Morgan and Albury, 2013; Steadman et al., 2007); the δ13C value for crocodile from Gilpin Point (−19.8‰) falls within the values reported elsewhere for modern and prehistoric terrestrial carnivores (c. −21‰ to −17‰; Bocherens and Drucker, 2003; Coltrain et al., 2004) and the values for Holocene Cuban crocodile fossils from other sites on Abaco (−20.9‰ to −16.4‰, N = 8; Hastings et al., in press). The Abaco tortoise was a terrestrial herbivore believed to have consumed mainly C3 plants (Franz and Franz, 2009; Steadman et al., 2007); the δ13C value for tortoise from Gilpin Point (−21.6‰) falls within the values reported elsewhere for modern and prehistoric terrestrial herbivores (c. −24‰ to −19‰; Bocherens and Drucker, 2003; Coltrain et al., 2004), including Holocene tortoise fossils from elsewhere on Abaco (−22.9‰ to −21.1‰, N = 5; Hastings et al., in press).
In the Greater Caribbean Region, the green turtle is a marine herbivore as an adult, although juveniles can be omnivorous or carnivorous (Bjorndal, 1997). Adult green turtles (females only) come ashore only to nest on sandy beaches. The δ13C value for the dated green turtle bone from Gilpin Point (−9.6‰) falls within the values reported for modern specimens from Long Island, Bahamas (−12.2‰ to −6.4‰; Vander Zanden et al., 2013).
Plants
The peat at Gilpin Point contains abundant wood, often in nearly vertical growth position, of buttonwood (C. erectus). The tops of dead buttonwood branches were also exposed in growth position on the sandy beach near the present shoreline c. 25 m north of where the peat was excavated (Figure 5), suggesting that the peat deposit extends at least this far northward. Buttonwood has lower tolerance to soil salinity than other mangroves (Medina, 1999; Rada et al., 1989). In the Bahamas, buttonwood typically is found inland from the three other species of mangroves (black mangrove Avicennia germinans, Acanthaceae; white mangrove Laguncularia racemosa, Combretaceae; red mangrove Rhizophora mangle, Rhizophoraceae) and is associated with slightly brackish or freshwater wetlands (Areces-Mallea et al., 1999).
The stump of the 14C-dated sabal palm (S. palmetto) also was found in growth position. In the Bahamas, sabal palm tolerates poorly drained soils and is associated with ephemeral freshwater ponds and areas with subterranean freshwater lenses near the surface (Areces-Mallea et al., 1999; Sullivan-Sealey et al., 2002). Sabal palm is prone to mortality when coastal flooding from sea-level rise exposes it to increasing salinity (DeSantis et al., 2007; Perry and Williams, 1996). Such flooding may have killed the individual that was 14C-dated, suggesting that this inundation took place no later than c. 800 cal. yr BP.
Mollusks
The sediment at Gilpin Point contains abundant remains of small gastropods as well as much rarer small bivalves. The molluscan assemblage was analyzed by JDS and JIM in detail from a single 3-L sediment sample taken from the area of densest bone concentration. Of 36 taxa recognized, 12 are terrestrial, 4 are freshwater, 1 is freshwater/estuarine, 2 are estuarine, 4 are estuarine/marine, and 13 are marine (Table 3). The majority of the most commonly occurring species are snails that require terrestrial or freshwater habitats. We interpret this trend in habitat preference to mean that the main sources of snails in the peat deposit originated inland (washing in with the charcoal) or in situ (in the freshwater or slightly brackish swamp).
Snails identified from a single sediment sample at Gilpin Point, Abaco, The Bahamas. B: bivalve; all others are gastropods. Macrohabitat categories – E: estuarine; F: freshwater; M: marine; T: terrestrial.

The species of mollusks in Table 3 are minute and do not include the large, edible, marine species (such as conchs, whelks, and top shells) that often are abundant in Amerindian middens in the Bahamas and southern coastal Florida (Jones O’Day, 2002; Keegan et al., 2008; Marquardt, 2010a, 2010b). We note, however, that a single specimen of Queen conch (Strombus gigas, Strombidae) and two of the West Indian top shell (Cittarium pica, Tegulidae) were recovered in situ alongside bones of the green turtle, only c. 2 m south of the snail sediment sample. These three specimens, each recovered in a broken condition typical of Amerindian middens (see Jones O’Day and Keegan, 2001), constitute the only mollusks from the Gilpin Point site that are likely to have been consumed or used as tools by prehistoric peoples.
Vertebrates
We identified 20 species of vertebrates from the fossils at Gilpin Point (Table 4). The five identifiable fish bones represent three species, the bonefish Albula vulpes (Albulidae; 2 vertebrae), bar jack Caranx ruber (Carangidae; palatine), and midnight parrotfish Scarus coelestinus (Scaridae; atlas, hypural). Each of these edible estuarine or marine species occurs regularly in West Indian archaeological contexts (Newsom and Wing, 2004).
Species of vertebrates identified from late-Holocene fossils at Gilpin Point, Abaco, Bahamas. ‘Status on Abaco’ categories – EA: extirpated on Abaco, but still exists elsewhere; EG: extinct globally; SE: still exists on Abaco. ‘Habitat preference’ categories: E: estuarine; F: freshwater; M: marine; T: terrestrial. Chelonia mydas and Pterodroma cahow are regarded as both marine and terrestrial because they most must come ashore to nest. Numbers in brackets represent the minimum number of individuals represented by the fossils of the five most common species. The species names for Leiocephalus, Cyclura, Chilabothrus, and Cubophis are based on the modern species that inhabit Abaco or nearby islands; the fossils from Gilpin Point for these three genera are diagnostic to genus but not to species.

Of the 17 species of amphibians, reptiles, birds, and mammals found in the Gilpin Point material, we first will document the seven that no longer occur on Abaco. The Cuban crocodile Crocodylus rhombifer is represented by c. 60 cranial and post-cranial bones (e.g. Figure 6) that depict two individuals (an adult and a sub-adult). Albury’s tortoise Chelonoidis alburyorum is represented by c. 60 post-cranial bones that are dominated by pieces of the carapace and plastron of a single adult individual with apparently healed bite marks from the Cuban crocodile on the outside of its carapace (Figure 6). Five post-cranial elements are referred to the rock iguana Cyclura cf. carinata (Iguanidae), another rather large, edible species (Carlson and Keegan, 2004; Keegan and DeNiro, 1988).

(a and b) Specimens of Cuban crocodile (Crocodylus rhombifer) and (c) Albury’s tortoise (Chelonoidis alburyorum) from Gilpin Point, Abaco, The Bahamas. (a) Lateral view of right maxilla (NMB.AB62.12), (b) dorsal view of frontal and parietal (NMB.AB62.11), and (c) dorsal view of nuchal and left peripheral 1 (NMB.AB62.1).
The Gilpin Point material of Bermuda petrel Pterodroma cahow (Procellariidae) consists of a nearly complete coracoid and two fragmentary humeri. These specimens agree with skeletal elements of P. cahow rather than those of the confamilial Audubon’s Shearwater Puffinus lherminieri (extant in the Bahamas) in these characters: larger overall size; medial surface of humeral end of coracoid singly concave (doubly concave in P. lherminieri); in medial or lateral aspect, procoracoid less flared dorsally; and humeral shaft less compressed (more circular in cross-section). The length of the coracoid (23.06 mm) agrees with that of fossils of P. cahow from Bermuda (22.5–23.7 mm, N = 15; Olson and Hilgartner, 1982).
The two pedal phalanges of white ibis Eudocimus albus (Threskiornithidae) from Gilpin Point represent nestlings or very young fledglings of a species that no longer breeds in the Bahamas. Eight fossils (sternum, radius, two carpometacarpi, four pedal phalanges) of the Cuban crow Corvus nasicus (Corvidae) occur at Gilpin Point. They represent the first record of C. nasicus from the Little Bahama Bank and the northernmost occurrence of species today confined to Cuba and the Caicos Islands. Finally, among the extirpated species, the Bahamian hutia Geocapromys ingrahami (Capromyidae) is recorded from Gilpin Point by a single molar. Common as a late-Quaternary fossil on many Bahamian islands (Morgan, 1989), G. ingrahami now survives only on several very small islands (see ‘Discussion’ section).
In all, 10 species of terrestrial vertebrates found in the Gilpin Point material certainly or probably still exist on Abaco. A single humerus represents the Cuban tree frog Osteopilus septentrionalis (Hylidae). The green turtle C. mydas is the most commonly occurring species of vertebrate in the deposit, represented by c. 80 bones of the carapace, plastron, and limbs from two very large individuals. Because unhealed bite marks from a crocodile are found on the inside (rather than outside) of the carapace of one individual (Figure 7), we interpret the bite marks to represent scavenging of a green turtle that people had butchered. Being so large, adult green turtles likely were butchered near the beach prehistorically and then shared among multiple households so that the bones of a single individual might be deposited in more than one midden (Newsom and Wing, 2004). With further exploration, perhaps more remains of fish and shellfish will be found at Gilpin Point.

Specimen of green turtle (Chelonia mydas; NMB.AB62.7) from Gilpin Point, Abaco, The Bahamas, in dorsal (a) and ventral (b) aspects.
A single caudal vertebra has the morphology of a gecko (Gekkonidae) but is not diagnostic beyond family level. A single genus of geckos (Sphaerodactylus) occurs on Abaco today (Schwartz and Henderson, 1991). A complete tibia is referred to the curly tailed lizard Leiocephalus carinatus (Leiocephalidae). This fossil tibia is 19.83 mm long, compared with 17.33 mm for the tibia of an adult modern specimen of L. carinatus from Cuba with a snout-vent length (SVL) of 98 mm and total length (TL) of 226 mm. Thus, the prehistoric curly tailed lizard from Gilpin Point had a SVL of c. 112 mm and a TL of c. 259 mm.
Four specimens are identified as anoles (Anolis, Polychrotidae), which are diverse and widespread in the West Indies, including the Bahamas (Henderson and Powell, 2009). One dentary and two maxillae of Anolis cf. sagrei are recorded from Gilpin Point by these characters: central cusp low, wide, and not excessively higher than marginal cusps; dentary robust and wide. This species exists in Abaco today. Another dentary is identified as Anolis cf. distichus because the diagnostic sharply pointed central cusp is distinctly higher than the well-pointed marginal cusps (see Pregill, 1982). Buckner et al. (2012) questioned whether or not A. distichus was historically introduced to Abaco; the Gilpin Point dentary is evidence that A. distichus is indeed indigenous to the island.
A single caudal vertebra and an edentulous fragment of dentary from a boa Chilabothrus cf. exsul (Boidae) are noticeably larger than that in the trope (dwarf boa) Tropidophis (Tropidophiidae). A mid-trunk vertebra is needed for more precise identification. The most commonly recovered small reptile from Gilpin Point is the snake (Bahamian racer) Cubophis cf. vudii (Dipsadidae; 12 specimens), identified by mid-trunk vertebrae with long, thin neural spines, elliptical prezygapophyses, and a thin, straight hemal keel with slight swelling and overhang at the condyle.
The yellow-crowned night-heron Nyctanassa violacea (Ardeidae) is a common resident species in the Bahamas, both today and as a fossil (Steadman et al., 2007; White, 1998). Our identification of a small rail (Rallidae sp.) is based on a fragmentary tibiotarsus of a size that may represent either the sora Porzana carolina or Virginia rail Rallus limicola; the specimen lacks diagnostic species-level characters. Both species presently occur in the Bahamas as non-breeding birds in freshwater and estuarine habitats.
Discussion
Landscape change
The setting and contents of the coastal peat deposit at Gilpin Point demonstrate how dynamic the Bahamian physical landscape can be over geologically short time frames. At both local and regional (inter-island) scales, these low islands can be affected dramatically by changes in sea level. The estimated sea level in the eastern Atlantic at 950 cal. yr BP (when humans arrived on Abaco) was ~20 cm lower than at present but was rising in response to the ‘Medieval Warm Period’ (Curtis et al., 1996; Keigwin, 1996; Kemp et al., 2011; Walker, 2013). Within a century or so, the shorelines degraded, inundating any coastal habitation sites that had been located on beaches seaward from the buttonwood and sabal palm wetlands at Gilpin Point. This wetland was also affected by the encroaching saltwater in at least two ways. First, the increased salinity and direct contact with the ocean would have made the wetland uninhabitable by buttonwood and sabal palm. Second, the peat deposit would have begun to get covered by beach sand.
We found no high-energy overwash sediment within the peat deposit, that is, no coarse sandy to pebbly to cobbly storm-surge facies such as those found in sediment cores in saline lakes on San Salvador (Park, 2012). The short interval of time represented by the peat is compatible with rapid sedimentation from inland areas following initial anthropogenic fires on Abaco. The time of deposition of peat at Gilpin Point corresponds with when people first arrived on Abaco (Steadman et al., 2007). Prehistoric deforestation, accompanied by fire, may have increased the rate of sediment deposition in Abaco’s wetlands, which has been reported independently on Abaco from the sediments of Emerald Pond blue hole (Slayton, 2010). The charcoal at Gilpin Point also corresponds in time with charcoal reported in sediments from a blue hole on Andros (Kjellmark, 1996). Similar increases in charcoal-rich sediment influx, believed to be associated with human activity, have been reported on other Caribbean islands and even the Atlantic coast of North America (e.g. Burney et al., 1994; Hilgartner and Brush, 2006; Horn et al., 2000; Lane et al., 2009). At Gilpin Point, the rapid deposition of peat (in less than a century, from c. 950 to 900 cal. yr BP) was followed by breaching of the former coastline (at least 200 m east of the modern one) and deposition of beach sand over the peat.
Aside from a lower sea level, deposition of the peat was enhanced by the limestone outcrop that exists just seaward from the peat deposit. This formerly continuous outcrop, which once would have served as a dam for the charcoal-rich sediment washing in from inland during deposition of the Gilpin Point site, has been broken and breached in several places in the past two decades during storms (e.g. Figures 5 and 8; PM, personal observation).

Section of limestone ridge broken by wave action, 13 November 2012, Gilpin Point, Abaco, The Bahamas. Note as well the black trunks of Conocarpus erectus.
Faunal change
With a late-Holocene age of ~950–900 cal. yr BP, the fossils from Gilpin Point represent a vertebrate community at the time of initial Amerindian (Lucayan Taíno) presence; 7 of its 17 species (41%) of amphibians, reptiles, birds, and mammals no longer live on Abaco. Among well-dated assemblages of West Indian vertebrates similar in age to those of Gilpin Point, much lower percentages of extirpated species are found on islands where humans arrived in the middle Holocene. For example, at the Trouing Jean Paul site in Hispaniola (14C-dated from ~1600 to 600 cal. yr BP), only 1 of 23 species (4%) of birds identified by Steadman and Takano (2013) no longer occurs on the island. Amerindians had arrived in Hispaniola four or five millennia earlier and already had extirpated the most vulnerable species of vertebrates (Steadman et al., 2005). A similar situation took place on some Pacific islands, where prehistoric human colonization led to huge losses of species within a century or two (Steadman et al., 2002). The species that survived the initial shock of human presence were likely to persist into modern times.
Four of the seven extirpated species (Cuban crocodile, Albury’s tortoise, white ibis, and Bahamian hutia) have been reported previously from other Holocene sites on Abaco (Franz and Franz, 2009; Franz et al., 1995; Morgan and Albury, 2013; Steadman et al., 2007). The three other extirpated species (Bahamian rock iguana, Bermuda petrel, Cuban crow) are new records for the island.
The morphology, systematics, and paleoecology of the Cuban crocodile and Albury’s tortoise on Abaco have been reviewed recently (Franz and Franz, 2009; Morgan and Albury, 2013; Steadman et al., 2007). Until their extermination shortly after human arrival, these two species were the largest terrestrial carnivore and herbivore on Abaco, respectively. The AMS 14C date on Cuban crocodile from Gilpin Point is the youngest direct age determination on the species and provides additional evidence that the species existed on Abaco when Amerinidians colonized the island. The Bahamian rock iguana is unknown on Abaco in modern times. We suspect that it was a food item for Amerindians on Abaco, as has been well documented on Grand Turk Island (Carlson and Keegan, 2004). Late-Quaternary fossils of rock iguana are also reported from Banana Hole, New Providence Island (Pregill, 1982).
The Bermuda petrel breeds today only on Bermuda, where it is common as a fossil (Olson and Meylan, 2009). This seabird was believed to have been extinct for several centuries until its rediscovery in 1951 and subsequent increase through intensive conservation efforts (Howell, 2012: 176–178; Wingate, 1972). An endangered species, the Bermuda petrel, does not occur in the Bahamas today, although six prehistoric bones were reported from Crooked Island (Olson and Hilgartner, 1982).
The white ibis has been recorded uncommonly in small numbers on several Bahamian islands (but not Abaco) in modern times but is not known to breed in the island group (White, 1998: 234). Both white ibis fossil specimens (pedal phalanges) from Gilpin Point represent nestlings or very young fledglings. Fossils of nestling and fledgling white ibises also occur in the late-Holocene sediments of Sawmill Sink on Abaco (Steadman et al., 2007), further substantiating that this large aquatic bird once bred on the island.
Nowadays in the Bahamian archipelago, the Cuban crow lives only in the Caicos Islands, far to the south of Abaco (Buden, 1987). Its main geographic range is on Cuba and Isle of Pines. The Cuban crow is known from late-Quaternary fossils on New Providence Island, Little Exuma Island, Crooked Island, and Eleuthera Island (Olson and Hilgartner, 1982; Steadman, unpublished data); the fossils from Gilpin Point represent its northernmost occurrence and its first record from any island on the Little Bahama Bank. They also demonstrate that the Cuban crow was still widespread in the Bahamas during the late Holocene.
The Bahamian hutia also was formerly widespread in the Bahamas but survives on tiny East Plana Cay (28 km east of Acklins Island) with introduced populations on certain islands in the Exuma Chain (Borroto-Páez et al., 2012; Morgan, 1989; Olson and Pregill, 1982). As with the Cuban crow (above), the range contraction of the Bahamian hutia was mostly a late-Holocene phenomenon.
Among extant species, the green turtle is a widespread marine herbivore that is well represented at archaeological sites throughout the West Indies (Newsom and Wing, 2004: 63). Adult female green turtles are vulnerable to predation by people when they come ashore to nest on sandy beaches. Our identification of A. cf. distichus is evidence that this anole was not introduced to Abaco in modern times but is indigenous to the island (contra Buckner et al., 2012). The Bahamian boa and Bahamian racer occur on Abaco today (Buckner et al., 2012; Krysko et al., 2013). Both of these snakes also were recovered from the late Pleistocene owl roost deposit in Sawmill Sink, Abaco, with the boa C. exsul reported as Epicrates striatus and the racer C. vudii reported as Alsophis sp. (Steadman et al., 2007; our updated nomenclature follows Hedges et al., 2009, and Reynolds et al., 2013).
Cultural origin of the bone deposit
Numerous unhealed bite marks on the thick carapaces of the green turtle are evidence of consumption by Cuban crocodiles. The fact that the bite marks are well developed on the insides of the carapaces but not evident on the outer surfaces, suggests that the crocodiles scavenged butchered green turtles rather than preying on living turtles. Humans are the only terrestrial predators in the Bahamas capable of splitting open the shell of an adult green turtle to expose the meat and organs inside the carapace and plastron. Human involvement in deposition of bones at Gilpin Point is supported further by the dense, midden-like concentration of large bones (crocodile, green turtle, and tortoise) in the peat, and the fact that some bones of both the green turtle and Abaco tortoise are charred (burned). Berman et al. (2013) suggested that Lucayans likely cooked/roasted sea turtles on their back in a hearth, which is compatible with the charring on the outside of the carapaces.
Other species in the assemblage, such as the three fish, the rock iguana, petrel, and crow, are also edible. The few large marine shells (queen conch, West Indian top shell) are broken in a way characteristic of prehistoric human processing. Nevertheless, the only unequivocal Amerindian artifact recovered thus far is a single shell bead (Figure 9). This round, well-polished specimen is 3.4 mm wide, 1.4 mm deep, with a drilled hole 1.5 mm wide. Similar shell beads have been reported from the Coralie Site on Grand Turk Island (Keegan, 1997: Figure 8.2). We note as well that the charcoal occurs throughout the sediments rather than in local concentrations suggestive of hearths.

Ground and polished shell bead from Gilpin Point, Abaco, The Bahamas.
In spite of using fine-mesh sieves (1.6 mm in the field, 1.0 mm in the laboratory), fossils of the three largest species of vertebrates (crocodile, tortoise, green turtle) far outnumber those of the 16 much smaller species at Gilpin Point (c. 200 vs 46 specimens). While prehistoric West Indian peoples typically preferred larger species when available (Carlson and Keegan, 2004), they also often deposited high numbers of small species in their middens (Newsom and Wing, 2004). Among the 16 smaller species in the deposit, the scarcity of fish in particular suggests that we have not yet sampled the densest concentrations of bones in the site because fish typically dominate the bone assemblages from prehistoric archaeological sites in the region (e.g. Jones O’Day, 2002; Wing, 2001).
The fossil sample from Gilpin Point probably is based on the prey remains left behind by humans as well as the Cuban crocodile. We found, nevertheless, very little non-boney evidence of prehistoric human activity at the site, such a pottery or shell midden (often dominated by conch shells Strombus spp.), both of which occur routinely and often in abundance in subsurface archaeological sites in the Bahamas and coastal Florida (Jones O’Day, 2002; Keegan, 2007; Keegan et al., 2008; Marquardt, 2010a, 2010b). Our failure to find any pottery or rich shell midden at Gilpin Point might be due to inadequate sampling; if the site extends inland beneath the beach ridge (which seems likely), then the peaty sediment that we have observed would represent less than 1% of the entire site. It could be that c. 950–900 years ago the Lucayans lived (at least seasonally) on a barrier beach located on the seaward side of the current beach (and the prehistoric peat deposit). Wave breaks at high tide may have contributed some relatively gentle overwash sand to the peat deposit (and brought small marine mollusks with them), but as noted, evidence of high-energy overwash sediment is lacking. The snails recovered in the deposit are consistent with sheet-wash transport of local terrestrial leaf litter and associated sediments into the peat layer that was forming in the wetland.
Conclusion
The Gilpin Point site is a late-Holocene (c. 950–900 cal. yr BP) intertidal, snail-rich, bone-rich peat deposit on the Atlantic coast of Abaco, Bahamas. The site is significant from several standpoints. The fossil plants and mollusks suggest that the site was a freshwater or somewhat brackish coastal wetland at the time the peat was deposited. The short time interval represented by the peat is compatible with rapid sedimentation following initial human-set fires on Abaco. The peat is exposed today only during exceptionally low tides, suggesting a lower sea level at the time of deposition as well as a degrading shoreline during the past millennium.
As a sample of a late Holocene vertebrate community at the time of first Amerindian presence, the fossils from Gilpin Point demonstrate that Abaco has undergone major biotic changes during the past c. 950 years of human occupation. Only 10 of the 17 identified species of non-fish vertebrates still live on Abaco. The loss of seven species of reptiles, birds, and mammals included Abaco’s largest terrestrial carnivore (Cuban crocodile) and its three largest terrestrial herbivores (Albury’s tortoise, rock iguana, and hutia). Unhealed crocodile bite marks on the inside of the carapaces of green turtles suggest scavenging of turtles that had been butchered by Lucayans.
The dense concentration of large bones in the peat, the charring on some bones of the green turtle and Albury’s tortoise, and the discovery of a shell bead, all point to a cultural origin of the bone deposit at Gilpin Point. A challenge now is to search the beaches of Abaco’s windward side to begin to learn whether the Gilpin Point site is truly unique or merely represents a more common situation that heretofore has been overlooked. During the lowest spring tides of the coming winter, we look forward to this search.
Footnotes
Acknowledgements
We thank the National Museum of the Bahamas/Antiquities, Monuments, and Museums Corporation (Michael Pateman, Keith Tinker), Abaco Friends of the Environment (Michael Albury, Olivia Patterson, Kristin Williams), and the Bahamas National Trust (Eric Carey, Lynn Gape, David Knowles) for furnishing permits, logistical support, and many other courtesies. Sabrina Bethel, Richard Franz, and Gary Morgan helped with preliminary collections. William Kenny (UF Department of Geology) kindly provided the loss on ignition data for our sediment samples. For comments that improved the manuscript, we thank Karen Bjorndal, Alan Bolton, Sally Horn, and Michelle LeFebvre.
Funding
Financial support was provided by the US National Science Foundation (grant BCS-1118369 to DWS, and BCS-1118340 to JF). This work represents the findings of the authors, and does not necessarily reflect the opinion of the sponsors.