
Abstract
We report a mid- to late-Holocene, non-cultural vertebrate assemblage from Garden Cave (site EL-229), Eleuthera Island, The Bahamas, with 2450 fossils representing 26 species. The chronology is based on accelerator mass spectrometer (AMS) radiocarbon (14C) dates determined directly on individual bones of the hutia (Geocapromys ingrahami), an extirpated species of rodent that dominates the bone assemblage at Garden Cave. Four AMS 14C dates from our excavation range from 1340 to 1280 cal. BP (surface of the site) to 4860 to 4830 cal. BP with depth. A hutia bone lying on the surface from elsewhere in the cave dated to 450 to 290 cal. BP, which is roughly the time of European and African contact on Eleuthera. Other extirpated species from Garden Cave are tortoise (Chelonoidis sp.), rock iguana (Cyclura sp.), skink (Mabuya sp.), parrot (Amazona leucocephala), crow (Corvus nasicus), and southeastern myotis (Myotis austroriparius). Each of these species may have survived on Eleuthera until sometime after the initial human occupation of the island (~1000 cal. BP), although we have direct AMS 14C dates for only the hutia. During the time of fossil deposition in Garden Cave, sea levels were approaching that of today, yet land areas were considerably larger than now, connecting Eleuthera to New Providence, and potentially to Exuma as well. Such relatively recent connections are important in explaining past and present distributions of terrestrial plants and animals.
Introduction
On a global scale, research on late-Quaternary vertebrate fossils from islands has revealed considerable extinction (species-level losses) and extirpation (loss of an insular population of a species that survives elsewhere). For insular fossil sites to yield evidence of lost species or populations is, by itself, no longer surprising. Beginning about a decade ago, a major focus of paleontological research on islands has been to develop chronologies for vertebrate fossil sites, most of which lacked radiometric age determinations (Steadman et al., 2005). Developing these chronologies has been essential to estimate when various insular species and populations were lost, as this is important to understand the possible causes of the losses, whether for large species such as mammoths (Martin and Stuart, 1995) and ground sloths (Steadman et al., 2005) or small species such as bats (Soto-Centeno and Steadman, 2015).
In the West Indies, as in many other island groups, bones of extinct or extirpated species occur not only in paleontological (non-cultural) sites but also in archeological (cultural) sites, thus establishing contemporaneity of lost species or populations with the arrival of people. The Bahamian islands were among the last West Indian island groups to be colonized by humans. It was only from ~700 to 1000 AD that people began to inhabit these islands (Carlson, 1999; Carlson and Keegan, 2004; Keegan et al., 2008; Newsom and Wing, 2004). Cultural sites, on average, are more likely to yield charcoal or other organic materials suitable for radiocarbon dating. For non-cultural sites, such as the one we report here, a chronology often depends on the bones (fossils) themselves retaining enough organic matter to yield radiocarbon dates.
The chronology of Holocene extinction and extirpation is well established on Abaco, a large island on the Little Bahama Bank (e.g. Hastings et al., 2014; Morgan and Albury, 2013; Soto-Centeno and Steadman, 2015; Steadman et al., 2007, 2014). Here, we report new evidence for Holocene faunal change on Eleuthera, another large island (by Bahamian standards) that lies 60 km southeast of Abaco on the Great Bahama Bank. We will focus on seven species of reptiles, birds, and mammals that no longer occur on Eleuthera.
Study area
From northwest to southeast, the Bahamian archipelago (Figure 1) extends from 90 km east of West Palm Beach, Florida (Grand Bahama) to less than 150 km from Cuba and Hispaniola (Great Inagua, Turks and Caicos Islands). The archipelago consists of 17 main islands and thousands of smaller cays. The Bahamian islands lie on four large and 13 smaller shallow water banks, all of which were exposed above the ocean during the late Pleistocene and early Holocene. The sea level–controlled expansion and contraction of land on these banks through the Quaternary (Hearty et al., 1998) had profound influences on the plant and animal communities across the island group (Steadman and Franklin, 2017).
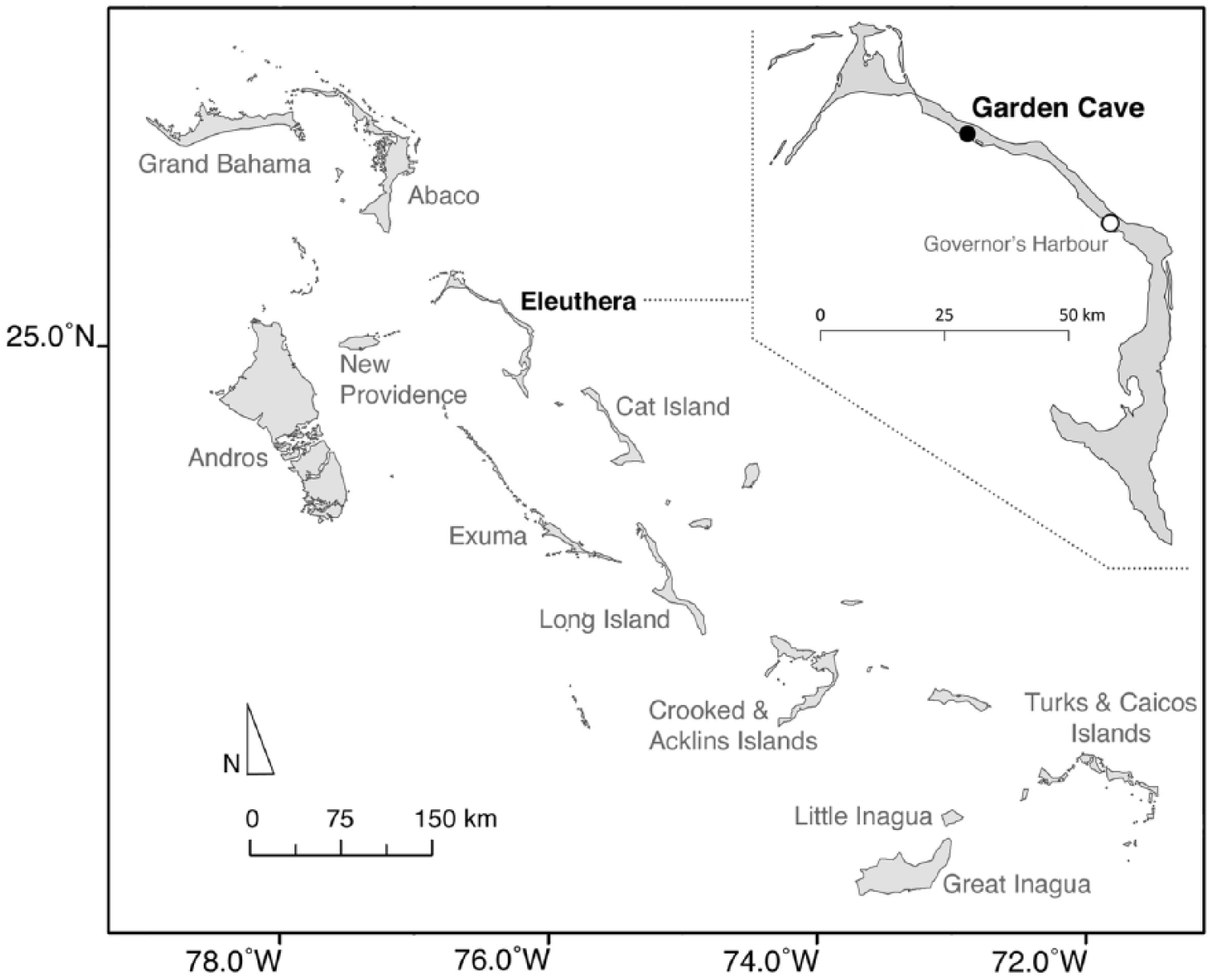
Map of the Bahamas archipelago. Eleuthera Island is identified in boldface. The location of Garden Cave is the black circle.
Located in the Central Bahamas, Eleuthera is a long (~160 km) and narrow (generally <3- to 4-km wide) island that trends NW to SE (Figure 1). Most of the land on Eleuthera (currently 471 km2) is below 10-m elevation, with the highest point only 35 m in elevation. The karst landscape of Eleuthera features many caves and other limestone solution features. The vegetation is a broadleaf dry forest and shrubland locally called ‘coppice’ (Franklin et al., 2015). Much of the coppice has been cleared for agricultural, commercial, and residential uses.
Local setting, materials, and methods
While exploring Eleuthera in March 2011, we came upon a shallow flank margin cave (see Mylroie and Mylroie, 2013) developed in an inland but seaward-facing cliff in the region of Hatchet Bay (N25°21.923′, W76°31.181′; Figure 1). In March 2012, we mapped this site called Garden Cave (site EL-229) using a Brunton compass and tape. Excavations were done with trowel in the soft sediment of Room 1, in arbitrary levels of ca. 10 cm each (measured as cm below surface or cmbs), following natural stratigraphic breaks when feasible. All excavated sediment was sieved through nested screens of 12.5-, 6.4-, 3.3-, and 1.6-mm mesh. The materials excavated from EL-229 are properties of the National Museum of The Bahamas, currently on loan to the Florida Museum of Natural History, University of Florida (UF). After being picked from the sediment, the bones were sorted by DWS into broad taxonomic categories and then dispersed to the appropriate specialists for identification (JIM and DWS for reptiles, JAS-C and DWS for mammals, and DWS for birds). The modern skeletons used in our osteological comparisons are from UF collections in the divisions of Herpetology, Mammalogy, Ornithology, and Vertebrate Paleontology. All radiocarbon dating and associated stable isotope analyses were done at Beta Analytic, Inc. Calibration data set follows IntCal13.
We calculated the area of exposed land (i.e. a.s.l.) at different time periods relative to present as in the work by Steadman and Franklin (2015). Late-Holocene relative sea level (RSL) estimates were taken from the literature (Milliken et al., 2008; Milne and Peros, 2013; Toscano and MacIntyre, 2003). Each RSL elevation was applied to a topographic grid at 1-m intervals (Smith and Sandwell, 1997), creating an output raster grid for land area above the specific elevation threshold connected to present-day Eleuthera. Then, the total number of land grid cells was summed and multiplied by the area of each grid cell to get a total land area estimate for the paleo-land mass encompassing modern Eleuthera.
Results
Stratigraphy and chronology
Based on our 1-m2 excavation, the sediments in Room 1 of Garden Cave (site EL-229) were developed as three fairly distinctive layers, designated as IA + IB, II, and III, with the difference between IA and IB readily discernible only in profile (Table 1; Figure 2). The upper level (0–10 cmbs) was dominated by Layer IA, with a minor component of IB. The next level (10–20 cmbs) was dominated by Layers IB and II, with a minor component of Layer IA. The next deepest level (20–30 cmbs) consisted mainly of Layer II, with some Layer III. The deepest level (30–40 cmbs) was almost entirely from Layer III, which lacked fossils and was much more indurated than the upper 30 cm of sediment. No cultural materials or charcoal were found at any level. In Table 3, the level designated as ‘10–20, 20–30 cm’ is part of the south wall of the excavation that collapsed, combining levels 10–20 and 20–30 cmbs.
Sediments excavated in Room 1, Garden Cave (site EL-229), Eleuthera Island, The Bahamas.

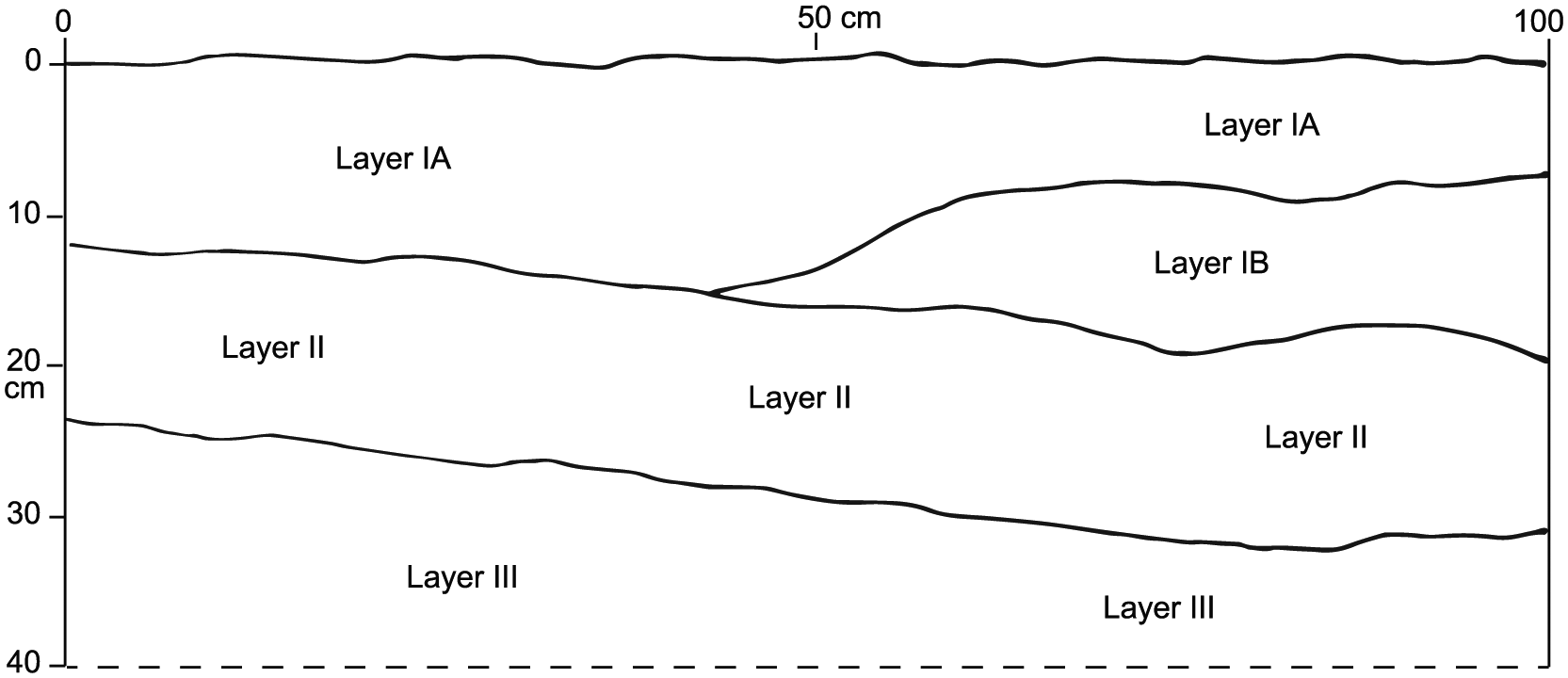
Profile of sediments in the north wall of Excavation Unit 1, Garden Cave (site EL-229), Eleuthera Island, The Bahamas.
About 3 m northeast of the excavation, we also excavated and sieved an area of extremely loose, unstratified sediment against the rear wall of Room 1, retrieving hundreds of bones. We called this place the ‘Crabby Pocket’ because of abundant remains of land crabs that had thoroughly churned the sediment.
The chronology for Garden Cave is based on five accelerator mass spectrometer (AMS) radiocarbon (14C) dates on individual, unburned bones of the extirpated hutia Geocapromys ingrahami, a rodent that once was the largest indigenous non-marine mammal on Eleuthera and elsewhere in the Bahamas. The youngest AMS 14C date is 450 to 290 cal. BP, from the unexcavated surface of Room 2 (Table 2). At the excavation in Room 1, a hutia femur on the sediment surface was nearly 2000 years older (2350–2300, 2240–2180 cal. BP). The soft sediment in Garden Cave has been conducive to vertical mixing of fossils, with the AMS 14C date from 0 to 10 cmbs (1340 to 1280 cal. BP) being roughly a millennium younger than the surface date. Furthermore, the oldest AMS 14C date from the excavation (4860–4830 cal. BP) was from 10 to 20 cmbs rather than 20 to 30 cmbs (Table 2). The loose, non-cultural sediment accumulated slowly in Garden Cave.
Accelerator mass spectrometer (AMS) radiocarbon (14C) dates on bones of hutia (Geocapromys ingrahami) from Garden Cave (site EL-229), Eleuthera Island, The Bahamas.

Stable isotopes
The stable isotope values (13C/12C) for organic carbon from hutia collagen from Garden Cave fossils vary from −20.1‰ to −18.9‰ (mean −19.5%; Table 2), which overlaps considerably with those reported from hutias in both cultural and non-cultural contexts from caves on Crooked Island (−20.7 to −19.1, mean −20.2; Steadman et al., 2017). For nitrogen (15N/14N), the five hutia specimens from Eleuthera are more positive (+6.1‰ to +7.5‰; mean +6.9‰) than in the four samples from Crooked Island (+4.7‰ to +6.9‰; mean +5.5‰; Steadman et al., 2017). The nitrogen data suggest a slightly smaller marine-derived component in the prehistoric terrestrial food web on Eleuthera than on smaller Crooked Island.
Vertebrate fauna
Bones are common in the three upper levels at Garden Cave (0–10 through 20–30 cmbs) but drop off to nearly zero by 30–40 cmbs (Table 3). We identified 26 species of vertebrates from the 2450 bones of indigenous, non-fish species recovered at EL-229. The fauna consists of one extant amphibian (8.2% of all bones), 12 species of reptiles (43.7% of bones, with the tortoise Chelonoidis sp., rock iguana Cyclura sp., and skink Mabuya sp. being extinct or extirpated), eight species of birds (0.4% of bones, with the parrot Amazona leucocephala and crow Corvus nasicus being extirpated), and five species of mammals (47.7% of bones, with the hutia Geocapromys ingrahami and the bat Myotis austroriparius being extirpated). The dominance of hutias in the bone assemblage is typical for sites on the Great Bahama Bank that presumably were deposited primarily by the large extinct barn owl Tyto pollens (Olson and Hilgartner, 1982), although no fossils of Tyto pollens (or the extant Tyto alba) were recovered. We regard the absence of species in the fossil assemblage to be of little significance because so many of the species we found are represented by one or only a few bones.
Stratigraphic summary of bones from Garden Cave (site EL-229), Eleuthera Island, The Bahamas.
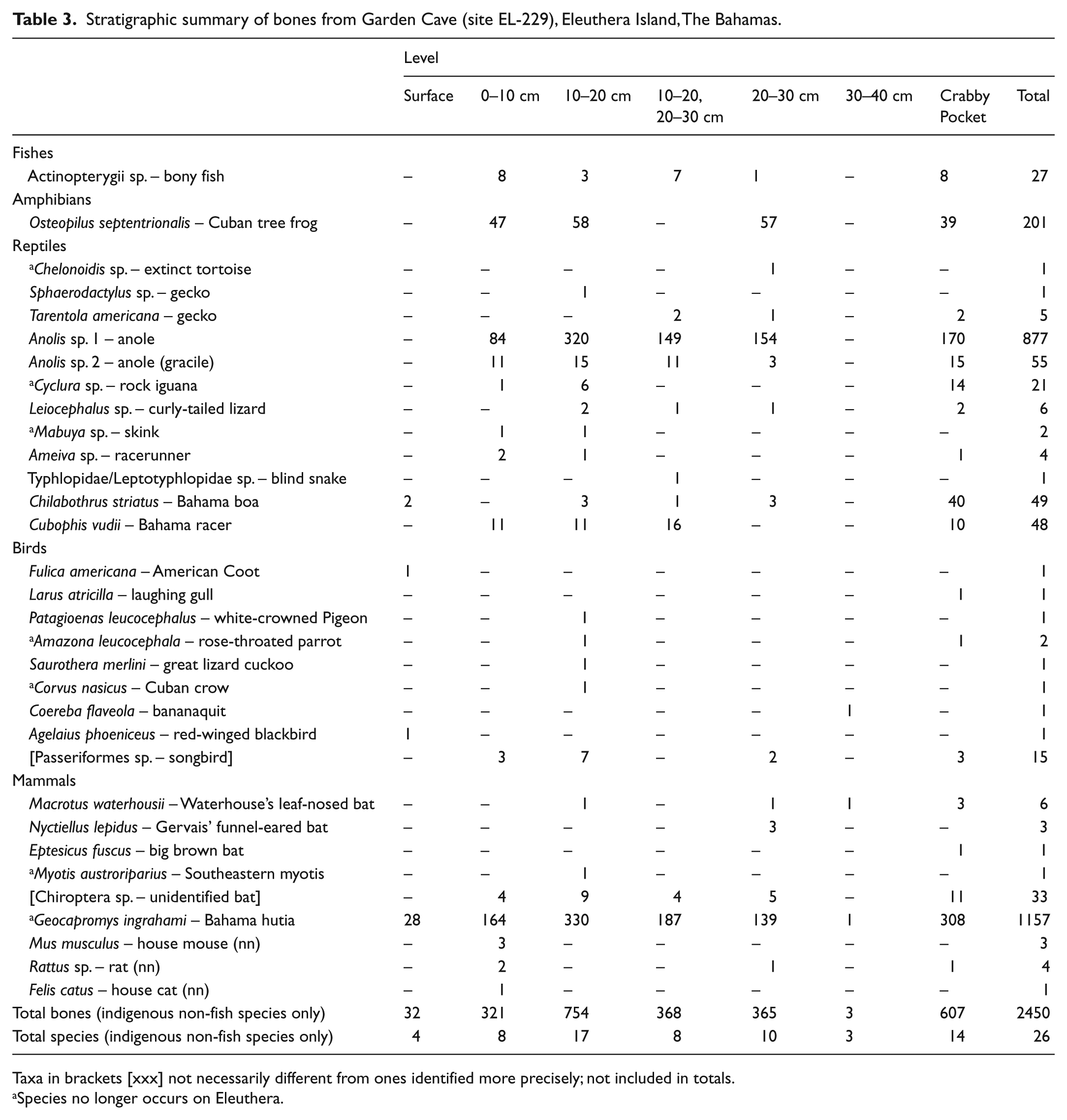
Taxa in brackets [xxx] not necessarily different from ones identified more precisely; not included in totals.
Species no longer occurs on Eleuthera.
Bones of three non-native mammals (Mus musculus, Rattus sp., Felis catus) were found in the upper strata at EL-229, as was the case at McKay’s Bluff Cave (Crooked Island), another site with loose, dry sediments (Steadman et al., 2017). The non-native mammal bones are consistently lighter in color and more modern in appearance than the bones of indigenous species. The two non-native rodents are common prey of modern barn owls (Tyto alba) on West Indian islands (Buden, 1974). Feral cats are widespread in the Bahamas, including Eleuthera (personal observation).
Sea level and land area
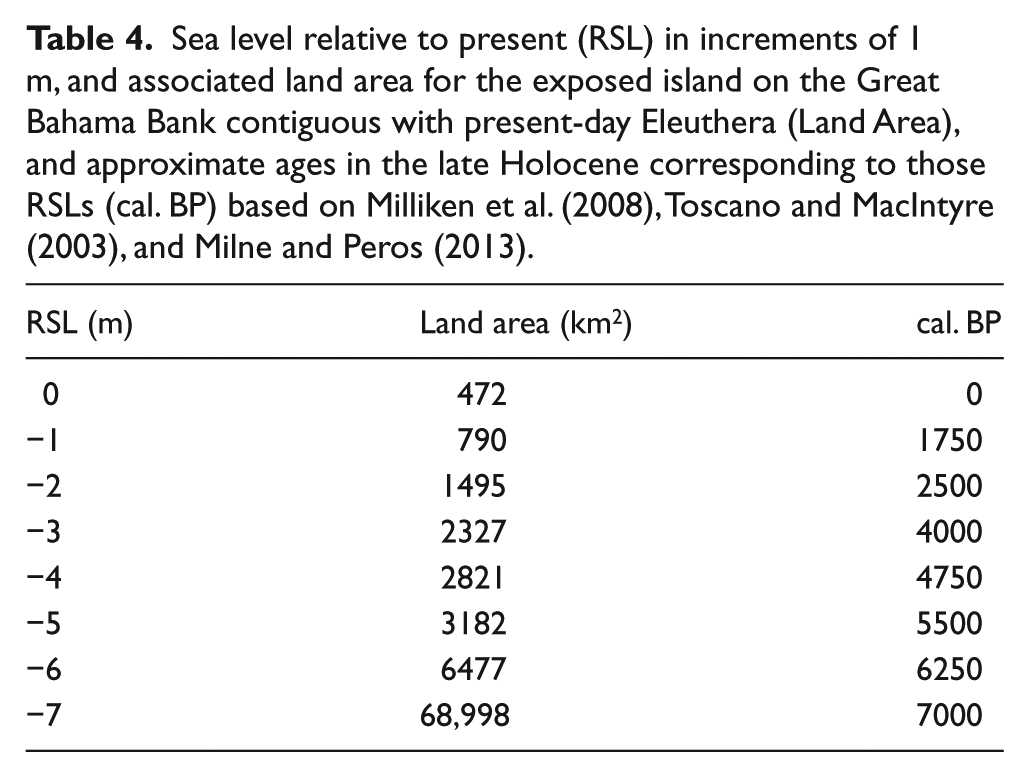
Looking at the RSL curve for the Bahamas during the Holocene (Milne and Peros, 2013: Figure 2), a lowering of sea level by only 1 m (−1 m) would increase the land area of Eleuthera from its current 471 to 789 km2. A decrease in just another meter (to −2 m) would connect Eleuthera to New Providence at ~2500 cal. BP and yield a land area >3× the current land area of Eleuthera (Table 4; Figure 3). An RSL of −4 m corresponds to a land area for Eleuthera of 2821 km2 at ~4750 cal. BP, which is roughly the age of the middle 20 cm of sediment and fossils from Garden Cave (Tables 2 and 4). When fossil deposition began at Garden Cave at ~4800 cal. BP, Eleuthera was ~6× larger than it is today.
Sea level relative to present (RSL) in increments of 1 m, and associated land area for the exposed island on the Great Bahama Bank contiguous with present-day Eleuthera (Land Area), and approximate ages in the late Holocene corresponding to those RSLs (cal. BP) based on Milliken et al. (2008), Toscano and MacIntyre (2003), and Milne and Peros (2013).
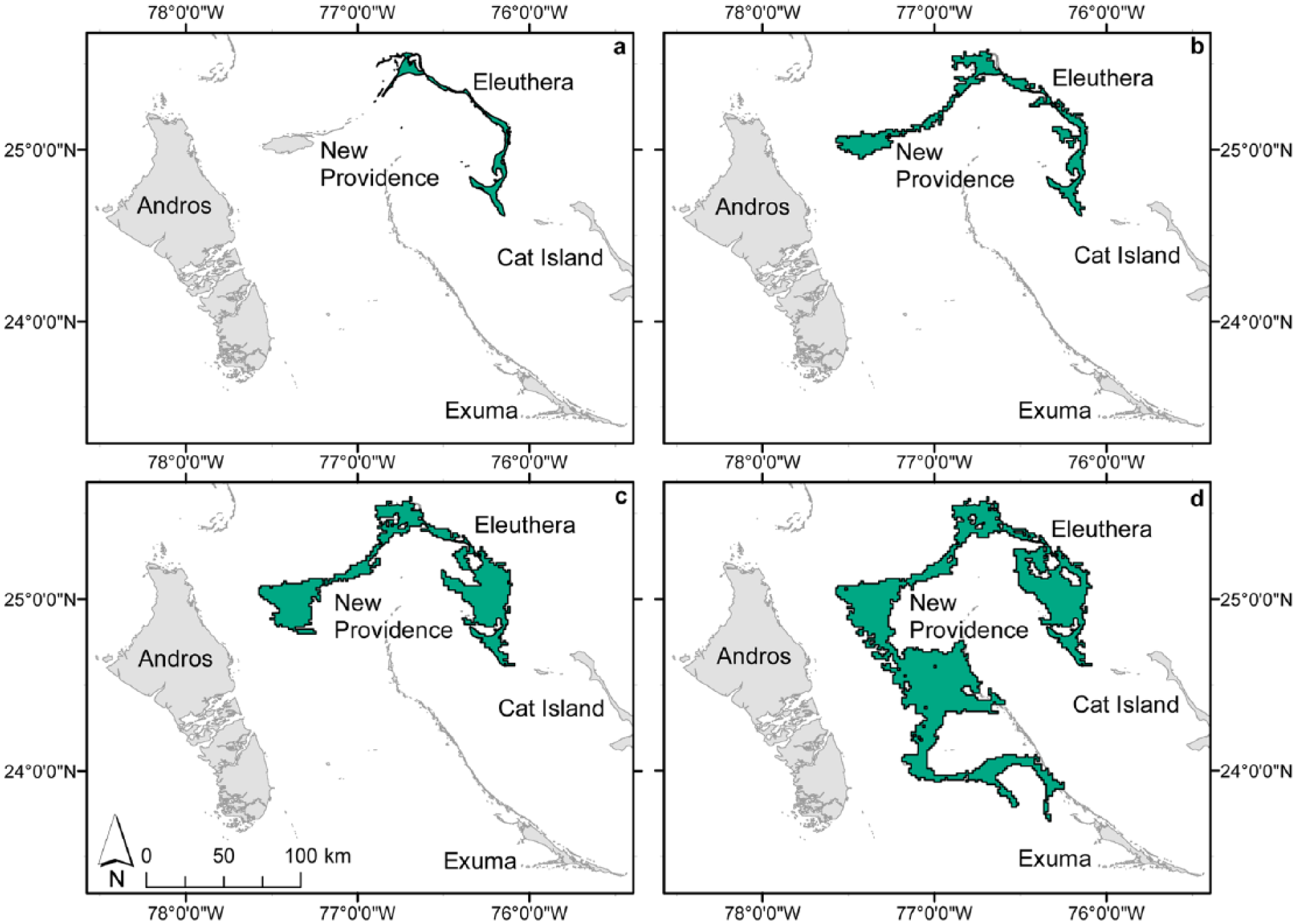
Estimated land extent of Eleuthera at (a) present, (b) −2 m relative sea level (RSL), (c) −4 m RSL, and (d) −6 m RSL (see Table 4 for chronology).
At a sea level of −6 m, corresponding to ~6250 cal. BP, Eleuthera’s land area was 6477 km2 (~14× current land area) and incorporated part of the Exuma Chain. At −7 m (~7000 cal. BP), the land area was increased greatly to nearly 70,000 km2 (~140× current land area) and incorporated not only the Exuma Chain but also the very large island of Andros (Figure 4). Given the late-Quaternary trend of slow subsidence of Bahamian islands (1–2 m per 100,000 years; Carew and Mylroie, 1995), our estimates of prehistoric land areas are minimum values.

Estimated land extent of the Great Bahama Bank paleo-island contiguous with modern Eleuthera at −7 m RSL (see Table 4 for chronology). Cat and Long islands are the only ones not connected at this time to other land on the Great Bahama Bank.
Discussion
Chronology
The very young AMS 14C date on a hutia bone from Room 2 of Garden Cave (450–290 cal. BP; Beta-330400) overlaps broadly with all four hutia-based AMS 14C dates from sites CR-5 (McKay’s Bluff Cave) and CR-25 (Crossbed Cave) on Crooked Island, each of which has a 2σ age estimate within the interval 510 to 305 cal. BP (Steadman et al., 2017). On both islands, therefore, hutias survived into historic times (= post-AD 1492), and we hypothesize that this trend will be found to be widespread on Bahamian islands (see below). From the excavation at Garden Cave, however, the four AMS 14C dates range from 4860 to 4830 cal. BP to as young as 1340 to 1280 cal. BP, a time interval that pre-dates the arrival of people (Lucayan Amerindians, who did not colonize the central and northern Bahamas until ~1000 cal. BP (Keegan, 1992, 1997; Steadman et al., 2007, 2014)). Thus, the bone assemblage from Garden Cave samples a mid- to late-Holocene time span of ~3500 years before the period of potential human impact.
Vertebrate fauna
We believe that the vertebrate fauna from Garden Cave is non-cultural in origin because (1) it lacks human artifacts made of shell, bone, or ceramics; (2) it lacks charcoal and burned bone; (3) all fish bones are small, as is typical of fish taken by non-human predators; and (4) the age range of the site pre-dates known human arrival in the Bahamas.
The presumably undescribed species of tortoise (Chelonoidis new sp.) from Eleuthera once was part of a major evolutionary radiation of Caribbean tortoises, no species of which survives (Franz and Franz, 2009). The material from Garden Cave consists of a single fragment of carapace, which is undiagnostic beyond representing a tortoise. For the undescribed species of tortoise (Chelonoidis sp.) on Crooked Island, the youngest AMS 14C date (925–785 cal. BP) resembles the more extensively studied situation on Abaco, where the youngest of five AMS 14C dates on the tortoise Chelonoidis alburyorum is 920–780 cal. BP (Franz and Franz, 2009; Hastings et al., 2014; Steadman et al., 2017, 2014). On Middle Caicos and Grand Turk, the extinction of an undescribed species of Chelonoidis may have taken place at a similar time (~900 to 800 cal. BP; (Carlson, 1999; Carlson and Keegan, 2004; Franz and Franz, 2009), although AMS 14C dates done directly on tortoise bone are lacking from Grand Turk. In each case, it seems likely that tortoises were lost within no more than several centuries after human arrival.
The rock iguana (Cyclura sp.) has not been recorded before (living or fossil) from Eleuthera or its offshore islands (Buckner et al., 2012). Remnant populations of several species of Cyclura occur locally (and never sympatrically) in the Bahamas and Turks and Caicos Islands today (Schwartz and Henderson, 1991: 388–402). Fossils from Abaco, New Providence, and Crooked Island show that rock iguanas once occurred essentially across the archipelago before human arrival (Hastings et al., 2014; Pregill, 1982; Steadman et al., 2017). All Cyclura fossils from Garden Cave are from adult individual(s). We note here as well that the Leiocephalus fossils are from large adults, which, by comparison with modern skeletons of known size, had snout–vent lengths of 106–114 mm.
The skink (Mabuya sp.) does not inhabit the Bahamian archipelago today except as Mabuya caicosae in the Turks and Caicos Islands (listed as Mabuya mabouya sloanei in Schwartz and Henderson (1991: 456–458) and as Spondylurus caicosae in Buckner et al. (2012); Hedges and Conn (2012)). Hedges and Conn (2012) split the genus Mabuya into 16 genera in the circum-Caribbean region; the possible osteological differences among these forms have not been explored, so we conservatively assign the two dentaries from Garden Cave to Mabuya sensu lato. Other species of Mabuya sensu lato occur in the Greater and Lesser Antilles. The Garden Cave specimens are the first Bahamian record for Mabuya sensu lato outside of the Turks and Caicos.
Both extirpated species of birds from Garden Cave once were widespread in the Bahamian archipelago but exist only very locally today. The rose-throated parrot (Amazona leucocephala) still exists in Cuba, the Cayman Islands, and, within the Bahamas, on Abaco and Great Inagua (White, 1998). Aside from Eleuthera, late-Quaternary fossils of Amazona leucocephala have been found on Abaco, New Providence, Middle Caicos, and Grand Turk, with historic (written) records from Acklins, Fortune, and Long Island (Carlson, 1999; Olson and Hilgartner, 1982; Steadman et al., 2015). The Cuban crow (Corvus nasicus) exists today on Cuba and the Caicos Islands but commonly occurs as a fossil on Abaco, Crooked Island, Little Exuma, New Providence, and Grand Turk (Olson and Hilgartner, 1982; Steadman et al., 2015, 2017).
The hutia (Geocapromys ingrahami) is a large rodent that once was widespread in the Bahamian archipelago (based on fossils), but now survives only on small, remote East Plana Cay. Four AMS 14C dates on individual hutia bones from sites on Crooked Island established its survival there into post-Columbian times (Steadman et al., 2017), similar to the situation on Eleuthera. Although bat fossils are scarce in Garden Cave, they do reveal that the southeastern myotis (Myotis austroriparius) formerly inhabited Eleuthera. To put the paucity of bats in Garden Cave into perspective, a very rich record of late-Holocene fossil bats on Abaco (hundreds of identifiable specimens) reveals extirpation of at least five species on that island within the last several millennium (Soto-Centeno and Steadman, 2015).
Holocene changes in the terrestrial landscape of Eleuthera
Because the Bahamian islands lie atop large but very shallow carbonate banks, fairly small changes in sea level can have major effects on land area. A considerable amount of attention has been paid to this phenomenon with regard to the major changes in sea level and land area (as well as in climate and habitat) that accompanied the Pleistocene–Holocene Transition (PHT) or the last glacial–interglacial transition from ~15 to 9 ka (Mylroie and Mylroie, 2013; Pregill and Olson, 1981; Steadman et al., 2015). Looking carefully at the smaller scale changes in Holocene sea levels (Milne and Peros, 2013), it becomes apparent that a number of Bahamian islands, including those on the Great Bahama Bank, continued to shrink in size even during the last half of the Holocene, the time interval represented by the fauna of Garden Cave (Figures 3 and 4).
Thus, for much of the time that Garden Cave was accumulating fossils, Eleuthera was roughly two to six times larger than it is today. The climate and habitat probably were similar to those of today, even though the island was substantially larger. We believe, nevertheless, that changes in island area of these magnitudes are typically, by themselves, not sufficient to cause extirpation of vertebrate populations (Franklin and Steadman, 2008). When accompanied by substantial changes in climate and habitat, as during the PHT, a number of species may be unable to withstand the multi-faceted changes (Soto-Centeno et al., 2015; Steadman and Franklin, 2017).
Knowing that many of the major islands of the Great Bahama Bank were connected as recently as the mid to late Holocene, we can establish the hypothesis that the species composition of bird and bat communities was essentially identical through most of the Holocene on the separate major islands, except for pine specialists. (New Providence and Andros are the only two islands on the Great Bahama Bank with pine woodlands today.) This hypothesis can be tested with both fossil and genetic data, providing rich opportunities for future research.
Footnotes
Acknowledgements
For permission to conduct the research and related courtesies, we thank the National Museum of The Bahamas/Antiquities, Monuments & Museum Corporation (Michael Pateman, Keith Tinker) as well as Michael Albury, Ruth Albury, Eric Carey, Ethan Freid, Olivia Patterson, and Kristin Williams. For help in excavating Garden Cave, we are grateful to Sarah Allen, Hannah Darcy, David Deren, Robbie Griffin, Jonathan Jordan, Andy Kilmer, Sean Moran, Jorge Pino, David Reed, Julie Ripplinger, Oona Takano, and Emily Woodruff. For assistance with sea-level analysis, we thank Riley Andrade. For comments that improved the manuscript, we thank Jessica Oswald and Gregory Pregill.
Funding
This study is supported by the National Science Foundation (BCS-1118340, BCS-1118369, and GSS-1461496), the UF Ornithology Endowment, and the Theodore Roosevelt and Gerstner Scholar Postdoctoral Fellowships at the American Museum of Natural History.