
Abstract
Dinoflagellate cyst (dinocyst) records from the southern Nordic Seas were compiled in order to evaluate the evolution of upper ocean conditions, on a millennial timescale and supported by a highly resolved record from the Vøring Plateau. After the transitional phase from the last deglaciation, three main phases define the Holocene. The early Holocene (>7.5 ka BP) features important numbers of cool-temperate species dominated by Nematosphaeropsis labyrinthus and Impagidinium pallidum in the west. The assemblage composition changes drastically at the transition from the early to the mid-Holocene, from when on Operculodinium centrocarpum dominates. The changeover is dated between ~6.1 and 7.5 ka BP, perhaps earlier closer to the Iceland–Scotland Ridge, and appears to be linked to the onset of a modern type of surface circulation. ‘Warmest’ assemblages occur at the Vøring Plateau shortly after the transition, when Atlantic waters also appear to have spread farthest westward. The recurrence of colder elements can be linked to cooling from ~2.4 ka BP at the Vøring Plateau and presumably earlier in the west but is difficult to date there because of the low sedimentation rates. This is a general issue in many areas of the Nordic Seas and appears to have an important effect on cyst concentrations and assemblage composition, with the possible loss of oxygenation-sensitive cysts in the older parts of the cores. Comparing dinocyst-based quantitative reconstructions with those retrieved from other plankton reveals a significantly different trend between proxies, linked to a differing autecological response to seasonal changes at their respective depth habitats.
Introduction
The interchange of ocean surface currents and heat redistribution that occurs in the Nordic Seas, which include the Greenland, Iceland, and Norwegian seas, is a key element in the regulation of modern climate (Figure 1): in the west, the East Greenland Current (EGC) defines the ice-laden Polar Domain (PD) where cold and low-salinity surface waters from the Arctic Ocean are carried southward along the eastern Greenland margins into the North Atlantic Ocean. Opposed to this is a northward flow across the eastern Nordic Seas of relatively warm and saline Atlantic waters originating from the Gulf Stream and defining the Atlantic Domain (AtD; e.g. Blindheim and Østerhus, 2005; Mork and Blindheim, 2000). These warm and salty waters feed either into the Arctic Basin or divert westward to mix with the cold polar waters in the seasonally ice-covered area of the Arctic Domain (ArD), where their density increase through heat loss may trigger overturning and deep water formation (e.g. Marshall and Schott, 1999). Since the pioneer work of Kellogg (1977), who illustrated the particularity of the present-day oceanography by comparison with that of the last 450,000 years based on planktic foraminiferal assemblages, many microfossil tracers have been used to reconstruct the Holocene paleoceanography of the Nordic Seas. Thus, further findings from planktic foraminifera (e.g. Andersson et al., 2010; Bauch, 1997; Bauch and Erlenkeuser, 2008; Bauch and Weinelt, 1997; Bauch et al., 1999, 2001; Hald et al., 2007; Risebrobakken et al., 2003, 2011; Sarnthein et al., 1995, 2003) have been complemented with insights from benthic foraminifera (e.g. Ólafsdóttir et al., 2010; Ślubowska-Woldengen et al., 2008, and references therein), coccoliths (e.g. Andruleit and Baumann, 1998; Belanger, 1982; Giraudeau et al., 2010), radiolarians (e.g. Cortese et al., 2005), diatoms (e.g. Berner et al., 2011, and references therein; Justwan and Koç, 2008; Justwan et al., 2008; Koç et al., 1993; Ran et al., 2006), alkenones (e.g. Bendle and Rosell-Melé, 2007; Marchal et al., 2002), dinoflagellate cysts (Baumann and Matthiessen, 1992; Matthiessen and Baumann, 1997; Van Nieuwenhove et al., 2013), and more recently the sea ice biomarker IP25 (Berben et al., 2014; Massé et al., 2008; Müller et al., 2012).
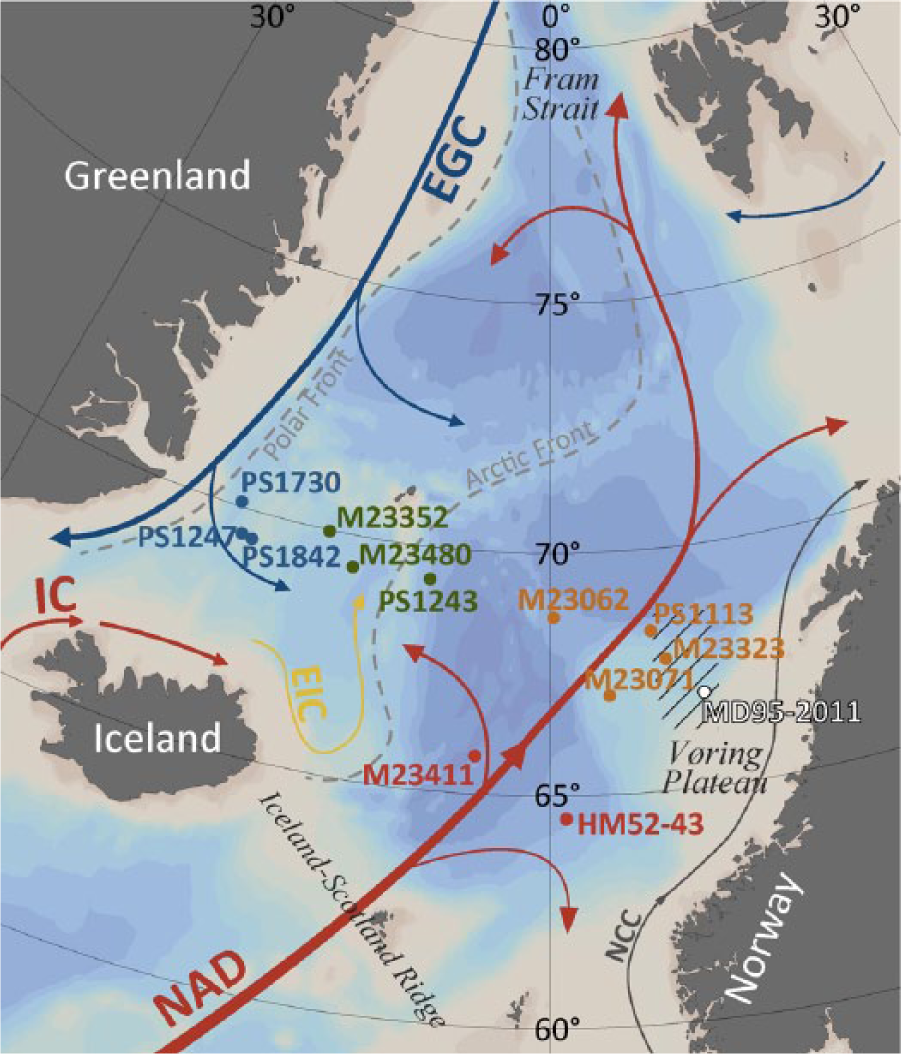
Map showing core locations, idealized modern surface current configuration, and the position of the Arctic and Polar fronts in the Nordic Seas. Red arrows represent warm currents, blue arrows cold currents. The core locations are color-coded according to the regional grouping applied in Figures 4 and 5. The hatched lines indicate the area delineated by the World Ocean Atlas 2001 data points used to calculate the average modern SST at the Vøring Plateau, as shown in Figure 6.
While the use of such a variety of proxy records that each contains specific seasonal and habitat depth information extends our understanding of the whole-water column development, many of the biogenic proxies also suffer from a number of disadvantages. Calcareous microfossils such as foraminifers and coccoliths can be subject to dissolution, particularly in the colder regions of the Nordic Seas where low diversity furthermore prevents them from revealing subtle environmental changes or distinguishing the PD from the ArD (Matthiessen and Baumann, 1997; Matthiessen et al., 2001; Solignac et al., 2008; Van Nieuwenhove et al., 2013). Moreover, despite their abundance in the water column, diatoms might be poorly preserved in sediment as the result of under-saturation of silica in the water column (e.g. Birks and Koç, 2002) which also affects radiolaria. By contrast, dinoflagellate cysts, which are commonly referred to as dinocysts and composed of highly resistant organic matter, are generally less sensitive to dissolution and display relatively high species diversity even in the coldest areas of the Nordic Seas (Bonnet et al., 2010; Rochon et al., 1999). As a result, many dinocyst records have been produced over the last two decades. The goal of this paper is to compile these published and unpublished dinocyst records from the southern Nordic Seas (i.e. south of Jan Mayen; Figure 1) in evaluating the Holocene change in sea surface conditions as compared to the scenarios provided by other planktic microfossil records.
Material and methods
Table 1 shows the position, water depth, and data analysis information for the sediment cores discussed here. In the manuscript, we will refer to the different cores only by their station name. The Holocene sediments of all cores consist of brown to olive bioturbated silty clays with a coarser component mainly consisting of foraminiferal shells in the eastern Nordic Seas but increasingly larger amounts of small quartz grains and dropstones toward the west. Samples were taken as 1-cm-thick slabs and freeze-dried before further treatment.
Background information for the investigated cores.
Next to the newly produced dinocyst census data, a number of published records as well as records from the unpublished PhD thesis of Baumann (2007) are included in this overview of Holocene dinocyst records from the southern Nordic Seas (Table 1). Although a few additional records exist from the region (Baumann, 2007; Baumann and Matthiessen, 1992; Eynaud et al., 2002), these were not included here because of their extremely low temporal resolution and/or lack of age control.
Age model
Age control of the studied sections is based on conventional (M23062) and accelerator mass spectrometry (AMS; all other cores) radiocarbon dating of planktic foraminiferal shells (ES Table A, available online). All newly produced as well as existing 14C dates were converted into calendar years using Calib 7 (Reimer et al., 2013). Because of considerable differences between the standard applied 400 years and actual reservoir ages for those areas in the Nordic Seas under the influence of Arctic water masses (e.g. Eiríksson et al., 2004), all cores were given a site-specific reservoir correction (with delta R ranging from −12 to +594; see ES Table A, available online) derived from the Google Earth interactive map for the circulation model of Butzin et al. (2005; http://radiocarbon.ldeo.columbia.edu/research/resage2.htm). Thus, all ages are given as 103 calendar years before 1950 CE (ka BP). Ages were interpolated linearly in between dated levels. When the surface sediment had not been dated, core top levels were given an age based on upward extrapolation from the highest dated levels or as derived from available sediment accumulation rate estimates. As such, the age at the core top sediment was estimated to be 2.163 ka in core PS1113 (Seiter et al., 2005) and 0.5 ka in core PS1730 (Matthiessen and Baumann, 1997). The surface of core PS1842 was given an age of 0 ka because extrapolation resulted in a negative age. Also, note that one radiocarbon date in core M23071 and one in core PS1842 were left out because they were considerable outliers in the age–depth plots with respect to the average sedimentation rates, presumably due to important admixture of underlying sediment deposited under a differing (deglacial) sedimentation regime (ES Figure A and Table A, available online).
Dinocyst analysis
Slides for microscopy observation were prepared by means of standard palynological techniques. For the new data presented here, the following procedure was applied: freeze-dried sediment was subjected to repeated treatments with cold ~10% HCl and ~40% HF, to remove the carbonate and siliciclastic fractions, and sieving over a 106-µm (cores M23480 and PS1247) and 6- to 10-µm polymer mesh, to concentrate the palynomorph-containing fraction and increase the efficiency of the maceration. The final, longest HF maceration step generally lasted about 12 h and was always followed by another HCl treatment to remove any precipitated fluorosilicates. The residue was then recovered on a polymer mesh (6–10 µm) and mounted in glycerin gelatin on microscopy slides for observation under a transmitted light optical microscope. No oxidation or staining was done. Gentle ultrasonication, when applied, was done for less than 30 s. One or two tablets of Lycopodium clavatum were added to each sample at the beginning of chemical processing in order to allow calculating palynomorph concentrations per gram of dry sediment (Mertens et al., 2009, 2012; Stockmarr, 1971).
A minimum of 300 identified cysts per sample was targeted but proved practically not feasible for some samples (see supplementary data). Samples with counts lower than 150 were not included in our dataset, except one sample from core PS1842 (29 cm, 137 cysts) and HM52-43 (5.5 cm, 105 cysts), and the lowest three samples of core M23323 (~50 cysts). Cysts were identified to species level whenever possible and conform to the most recent paleontological nomenclature system (Fensome et al., 2008). However, note that, for reasons of consistency with previous paleontological studies, we chose to retain the paleontological name ‘Operculodinium centrocarpum’ (sensu Wall and Dale, 1966) when referring to cysts of Protoceratium reticulatum (Paez-Reyes and Head, 2013). Brigantedinium simplex and Brigantedinium cariacoense were grouped with other smooth round brown cysts under Brigantedinium spp., as unfavorable orientation or preservation often not allows these cysts to be identified at species level. Furthermore, Impagidinium spp. comprise all Impagidinium species other than the cold taxon Impagidinium pallidum. Percentage calculations are based on the total sum of cysts excluding unidentified and reworked specimens. A full list of recovered species is given in Table 2, and all raw data can be accessed through the electronic supplements to this manuscript.
List of recovered dinoflagellate cyst species.

Reconstructions from the modern analogue technique
Quantitative estimates of past winter and summer sea surface temperature (SST) and salinity (SSS) as well as seasonal sea ice cover (SIC) were done using the modern analogue technique (MAT; de Vernal et al., 2005, 2013; Guiot and de Vernal, 2007). The procedure we used involves a log-transformation of the percentage data to give more weight to (ecologically constrained) rare taxa, followed by calculation of the sea surface parameters based on the five closest analogues in the modern dataset from the Northern Hemisphere (n = 1492; de Vernal et al., 2013), with decreasing weight in the calculations for each analogue according to its decreasing similarity (i.e. increasing statistical distance) with the fossil assemblage. An analogue is rejected when its statistical distance is larger than the threshold value for reliable analogues. To estimate the reliability of the quantitative reconstructions, a code is attributed to each sample following de Vernal et al. (2013) based on the number of cysts counted, the similarity of the modern analogues to the fossil assemblage, and the number of analogues found below the similarity threshold. The reliability codes calculated in this way vary between A for most reliable estimates and D for reconstructions based on low cyst counts and/or poor analogues.
The MAT method permitted to identify five good modern analogues below the threshold values for all samples, and all reconstructions but one (PS1842, 25 cm, 10.4 ka BP) could be attributed reliability code A or B (see supplementary data). Despite the robustness of the reconstructions as estimated above, we are aware that none of the approaches for quantitative reconstructions of climate parameters based on microfossil assemblages is unequivocal (e.g. Guiot and de Vernal, 2007), and some concern has been raised about the calculation of the error of prediction from MAT reconstructions (e.g. Guiot and de Vernal, 2011a, 2011b; Telford and Birks, 2011). To account for the uncertainties inherent to quantitative reconstructions, and also accommodate the age uncertainty on the low temporal resolution cores, the MAT results are used here to assess regional rather than sample- and core-specific fluctuations in SSS, SST, and SIC. For this purpose, the low-resolution cores were grouped according to region: the Atlantic Nordic Seas (M23411 and HM52-43); the Vøring Plateau (M23071, M23062, and PS1113); the Central Nordic Seas (M23352, M23480, and PS1243); and the Western Nordic Seas (PS1730, PS1247, and PS1842). The Holocene interval was then divided into 2-ka time slices for which the average values (most probable and possible extremes) for SST, SSS, and SIC were calculated per region (Figure 5). Core M23323 was not included in these regional averages but shown separately in Figure 5 to avoid bias due to its higher temporal resolution and age model difference (see below). Note that we remain cautious not to over-interpret conspicuous single events in the highly resolved record of core M23323 (Figure 6; ES Figure B, available online), and only the millennial-scale evolution of Holocene sea surface conditions will be addressed in the discussion.
Results
Only the unpublished census results will be outlined in detail here, with core M23323 taken as ‘benchmark’ because of its high temporal resolution (Table 1). We refer to the original publications for the census data of the published records.
Atlantic domain
Core M23323 yielded fairly rich samples in terms of species numbers and cyst concentrations, with peaks up to nearly 200,000 cysts g−1 and dropping under 10,000 cyst g−1 only in the few samples older than 12 ka (i.e. below 190 cm core depth). The section can broadly be divided into four zones (Figure 2a).
Before ~12 ka BP, Operculodinium centrocarpum is the most common species, accompanied by significant numbers of Spiniferites elongatus, Spiniferites ramosus, and Bitectatodinium tepikiense.
The second zone spans from ~12.1 to ~6.2 ka BP and is characterized by a dominance of Nematosphaeropsis labyrinthus (40–60%). Operculodinium centrocarpum constitutes about 25% of the assemblages, with Brigantedinium spp., Spiniferites species (i.e. Spiniferites elongatus, Spiniferites ramosus, unspecified Spiniferites spp.), and cysts of Pentapharsodinium dalei – all decreasing upward – making up most of the remaining assemblage share. Impagidinium pallidum occurs in low but notable numbers, while the previously abundant Bitectatodinium tepikiense is only observed in very low numbers within this interval. A noteworthy increase of Impagidinium spp. can be seen between ~8.4 and ~9.7 ka BP.
A clear dominance of Operculodinium centrocarpum (up to 95%) distinguishes the third interval, from ~6.1 to ~2.4 ka BP. All other species have their lowest relative abundances within this interval, with the exception of Spiniferites mirabilis.
Finally, Bitectatodinium tepikiense reappears after ~2.4 ka BP having been completely absent from the previous interval. Cysts of Pentapharsodinium dalei, Impagidinium pallidum, and Nematosphaeropsis labyrinthus are the other species that make up for the slight decrease of Operculodinium centrocarpum, which nonetheless remains clearly dominant.

Assemblage composition, cyst concentrations, and concentrations of reworked cysts per gram dry sediment (rew./g; when available) in (a) high-resolution core M23323, (b) the low resolution cores from the Vøring Plateau, and (c) the southernmost ‘Atlantic’ cores. The bars in core M23323 illustrate the subdivision of the Holocene section into four intervals based on the changes in the assemblage composition.
This assemblage pattern can also be seen in the other records from the AtD, albeit with some minor differences in the timing of the major assemblage changes (Figure 2b and c). The biggest difference in timing is observed in core M23062 off the western edge of the Vøring Plateau, where the assemblage changes seem to be shifted by about 2 ka, and the transition from zones 3 to 2 also appears to occur slightly earlier in the two southern cores (M23411 and HM52-43) with respect to the more northern low-resolution cores. Cyst concentrations are about one order of magnitude lower in the low-resolution cores than in core M23323. Assuming regionally comparable surface water primary productivity, the counterintuitive observation of highest concentrations in the core that featured highest sediment accumulation rates – and thus weaker dilution of original cyst fluxes – is a probable consequence of the fact that dinocysts behave like the fine silt fraction upon sedimentation. Therefore, cysts were efficiently advected together with fine sediment particles to sediment focusing site M23323 (Rumohr et al., 2001). The additional loss of peridinioid (‘heterotrophic’) cysts, which are more sensitive to oxidation than other dinocyst taxa (Zonneveld et al., 2008), could also be a consequence of the low sediment accumulation rates and will be addressed in the relevant sections further below.
Arctic–Polar domain
Cyst concentrations are (much) lower in the ArD and PD (which includes site PS1243 just east of the modern Arctic front) compared to the AtD, fluctuating between ~100 and 2000 cysts g−1 and only occasionally attaining more than 10,000 cysts g−1 (Figures 3 and 4). Similar to the Vøring Plateau cores, the Holocene interval can be divided into 3–4 different assemblage zones, depending on the temporal coverage of the respective core.
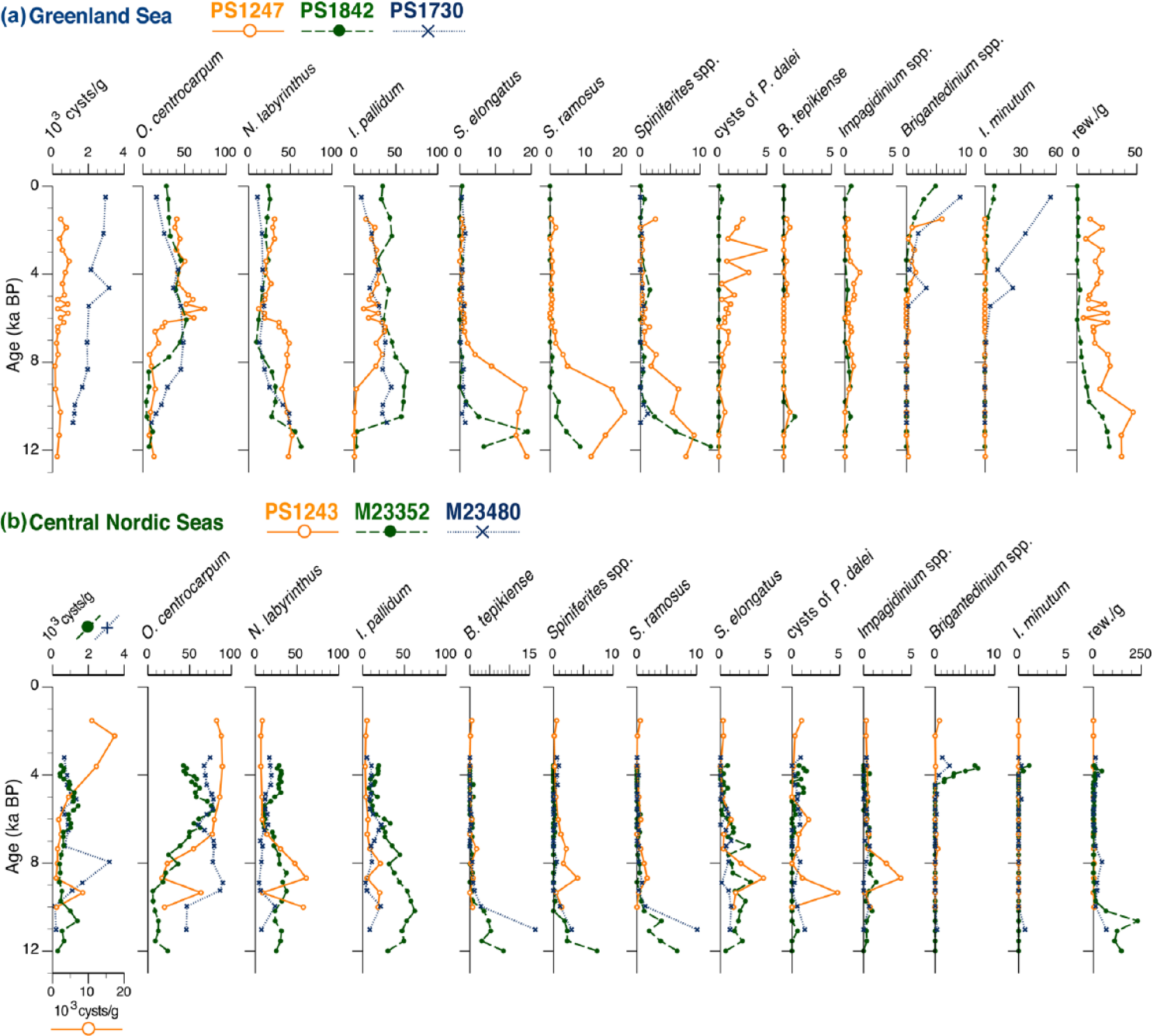
Assemblage composition, cyst concentrations, and concentrations of reworked cysts per gram dry sediment (rew./g; when available) in the cores from (a) the Greenland Sea, and (b) the Central Nordic Seas.
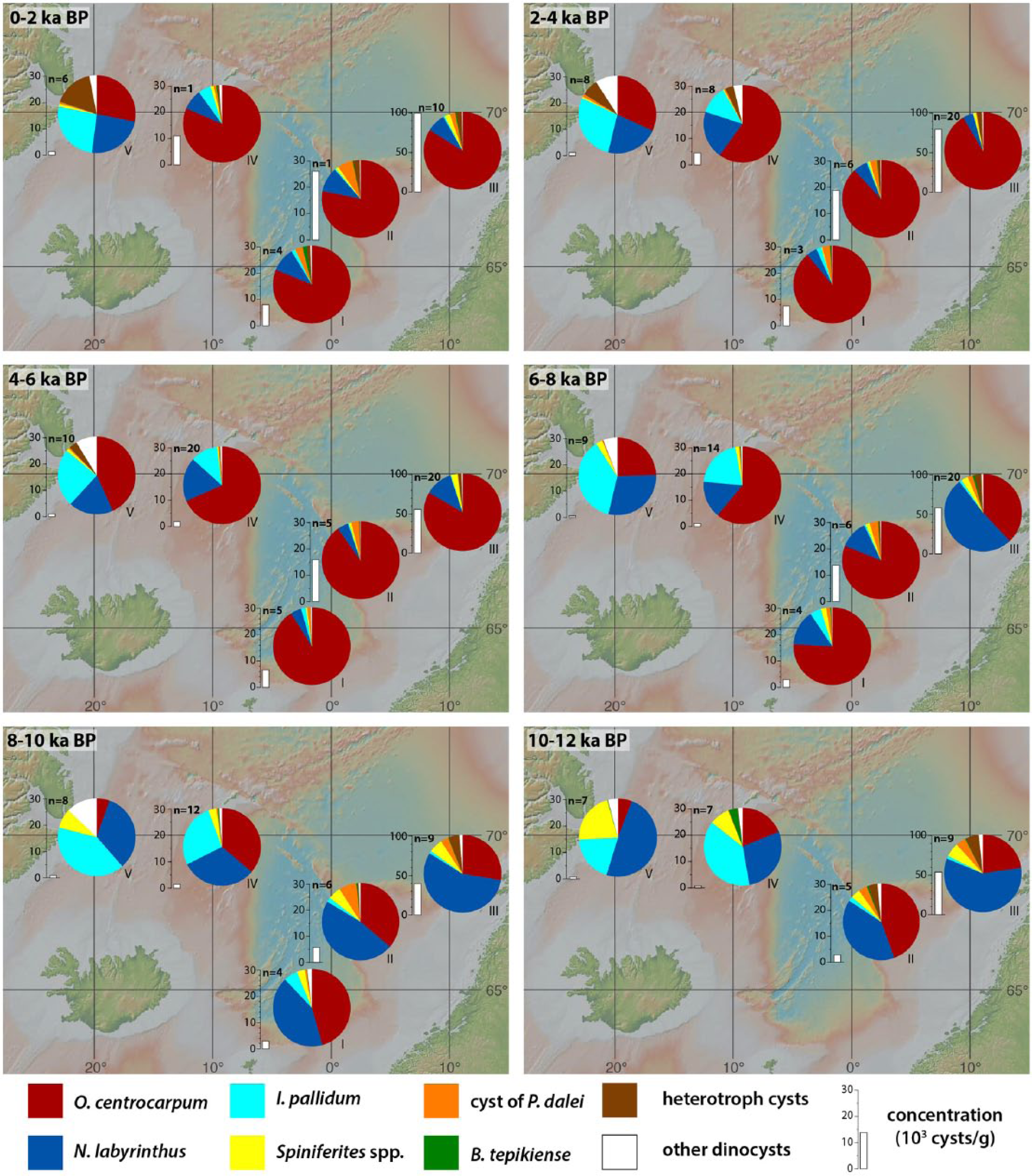
Regionally and time-sliced averaged (see main text) relative abundances of the main dinocyst taxa (with indication of the number of samples n used in calculating the average relative abundances), and absolute cyst concentrations (103 cysts g−1), for (I) the southernmost ‘Atlantic’ cores, (II) the low-resolution cores from the Vøring Plateau, (III) high-resolution core M23323, (IV) the Central Nordic Seas, and (V) the Greenland Sea.
A lowermost zone (not covered by core PS1243) is dominated by Nematosphaeropsis labyrinthus and/or Impagidinium pallidum, except for core M23480 where Operculodinium centrocarpum outnumbers both species. As with the lowermost interval at the Vøring Plateau, this interval shows significant occurrences of Spiniferites ramosus and Spiniferites elongatus, as well as Bitectatodinium tepikiense in the eastern ArD (Figure 3b) or Spiniferites spp. in the western ArD or PD (Figure 3a), which altogether appear to have prevailed longer (until ~9.5 ka BP) in these colder regions of the Nordic Seas. The dominance of Impagidinium pallidum and Nematosphaeropsis labyrinthus (Operculodinium centrocarpum for core M23480) continues until ~7 ka BP, but the abundance of most other species has fallen going into this younger age interval. A slight increase in Impagidinium spp. can be seen in some cores. Around ~7 ka BP, the assemblages shift toward a marked dominance of Operculodinium centrocarpum at the expense of overall species diversity, with Impagidinium pallidum and Nematosphaeropsis labyrinthus becoming equally important secondary species (Figures 3 and 4). Note that this marked abundance increase in Operculodinium centrocarpum appears to be shifted >2 ka downward in core M23480 with respect to the other cores. The final assemblage shift occurs around 2.4 ka BP at the Vøring Plateau site but is equivocal in the Greenland and Icelandic Sea because of the inaccurate chronostratigraphy of the core tops. Nonetheless, an overall abundance increase toward the core top can be seen in several species, most notably Brigantedinium spp. and Islandinium minutum (PS1730; Matthiessen and Baumann, 1997).
Discussion
Millennial-scale oceanographic changes
The dinocyst record of core M23323 confirms and refines insights into the upper ocean conditions at the Vøring Plateau during the Holocene as sketched by the previously published low-resolution dinocyst records (Baumann and Matthiessen, 1992). The relatively high sedimentation rates at site M23323 surely led to limited bioturbational smoothing of the original signal, which permits an accurate recording of regional pelagic fluxes. Overall, all dinocyst records seem to provide a coherent image of sea surface conditions in the Nordic Seas during the postglacial, with the distinction of four major climatic phases and three since 11 ka BP when considering the Holocene sensu stricto. These three Holocene phases appear to be a robust feature in various proxy records from the Nordic Seas and northern North Atlantic, including foraminifers, radiolaria, and dinocysts (e.g. Andersen et al., 2004; Cortese et al., 2005; Harland and Howe, 1995; Ólafsdóttir et al., 2010).
Younger Dryas – Earliest Holocene transition
The assemblages in the cores covering the transition from the cold Younger Dryas (YD; ~11.5–12.5 ka BP) into the Holocene reflect the deglacial conditions that existed in the upper ocean during that time: Bitectatodinium tepikiense and cysts of Pentapharsodinium dalei have been associated with enhanced stratification and seasonally strongly contrasting conditions (de Vernal et al., 2005, and references therein; Howe et al., 2010), which can typically be induced by ice(berg) melt freshwater input and offer a favorable time window for growth to a number of species. The occasional presence of drifting ice is further suggested by high amounts of reworked pre-Quaternary cysts and Brigantedinium spp. (Figures 2 and 3), whose cyst-producing heterotrophic dinoflagellates profit from the increased food abundance that can characterize drifting ice(bergs) (Smith et al., 2007). In modern sediments, Bitectatodinium tepikiense is furthermore a typical indicator for the subpolar-temperate boundary (Dale, 1996), and its time-transgressive abundance drop in the eastern and central Nordic Seas likely reflects the northwestward retreat of this frontal zone across the core sites during the YD-Holocene transition (cf. Eynaud et al., 2000; Grøsfjeld et al., 1999; Penaud et al., 2008; Van Nieuwenhove et al., 2011). Indeed, our and diatom data suggest that milder conditions gradually developed between 12 and 11 ka BP in the AtD (Figure 2; Berner et al., 2010; Justwan and Koç, 2008), while deglacial conditions persisted 1–2 ka longer in the cold domains of the Nordic Seas (Figure 3). Multiproxy evidence suggests that the Polar front retreated from the northern Iceland shelf from around 10.5 ka BP (Ólafsdóttir et al., 2010, and references therein).
Early Holocene
An early Holocene interval is defined here based on the second dinocyst assemblage zone and spans from the time after the YD-Holocene transition (~11.5 ka BP in the east, ~9 ka BP in the west) to roughly around 7 ka BP. The dominance of cold (Impagidinium pallidum) to cold-temperate species (cysts of Pentapharsodinium dalei, Nematosphaeropsis labyrinthus, and Spiniferites elongatus) during the early Holocene reveals the prevailing presence of fairly cool conditions in the surface waters, despite high boreal summer insolation at this time. Indeed, even at the Vøring Plateau, MAT reconstructions suggest the occasional presence of winter sea ice (Figure 5). Diatom-based transfer functions reveal that sea ice is gone at the Vøring Plateau at latest by May from 11 ka BP onward (Justwan and Koç, 2008), but the dinocyst signal of cool conditions nonetheless opposes various other plankton records that show highest temperatures in line with Northern Hemisphere summer insolation (Figure 6). This apparent proxy-specific signal suggests a different seasonal response to the environmental variables and is discussed in more detail in section ‘Comparison of micropaleontogical records of surface ocean conditions’. Although MAT shows that seasonal sea ice clearly was present in the western Nordic Seas (Figure 5; ES Figure B, available online), the assemblages there lack any heterotrophic cysts despite high prey abundance that typically can occur around drifting ice (see section ‘Younger Dryas – Earliest Holocene transition’; Smith et al., 2007). This discrepancy can probably be attributed to the low productivity of a short growing season combined with the loss of these sensitive cysts trough oxygen degradation as a result of the slow burial rates that characterize the Holocene at these sites (Zonneveld et al., 2008).
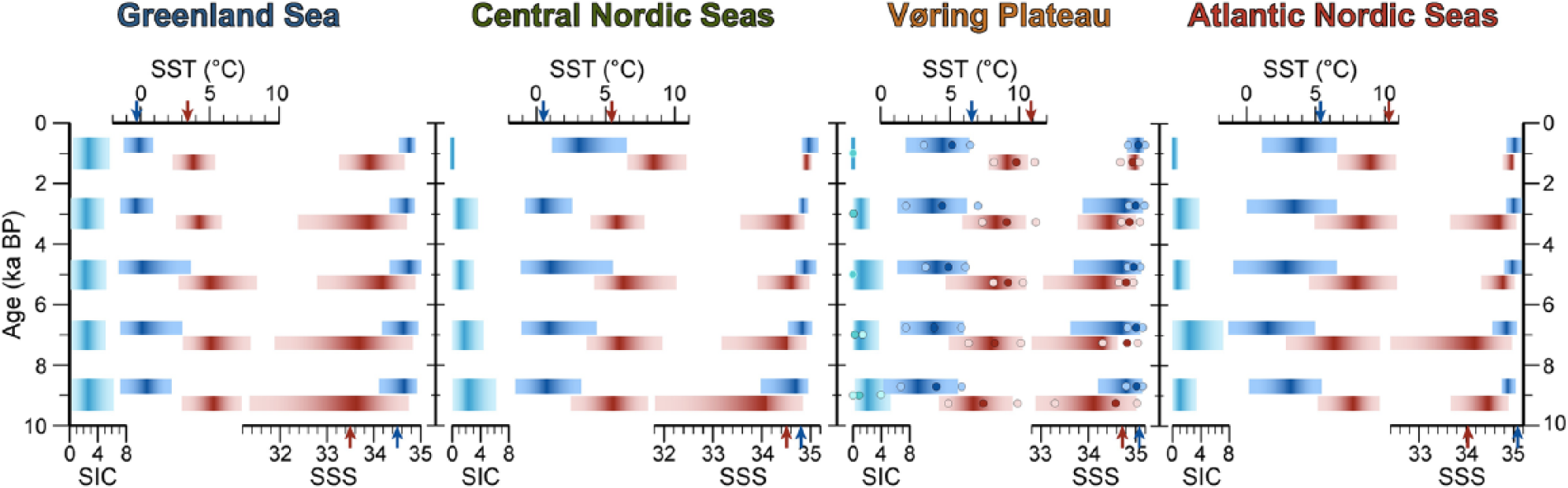
Regionally and time-sliced averaged (see main text) quantitative reconstructions from the dinocyst assemblages for sea surface temperature (SST), sea surface salinity (SSS), and sea ice cover (SIC, in months per year). Red bars indicate summer and blue bars winter SST and SSS. The dark band represents the ‘best’, most probable reconstructed value and fades out toward the minimum and maximum analogue value. The dots in the Vøring Plateau cartoon represent the values reconstructed from high-resolution core M23323. Modern values are indicated by arrows (from the World Ocean Atlas, 2001 (Conkright et al., 2002; Stephens et al., 2002); at site PS1247 for the Greenland Sea; at site M23480 for the Central Nordic Seas; averaged over the hatched area in Figure 1 for the Vøring Plateau; average of both core sites for the Atlantic Nordic Seas).
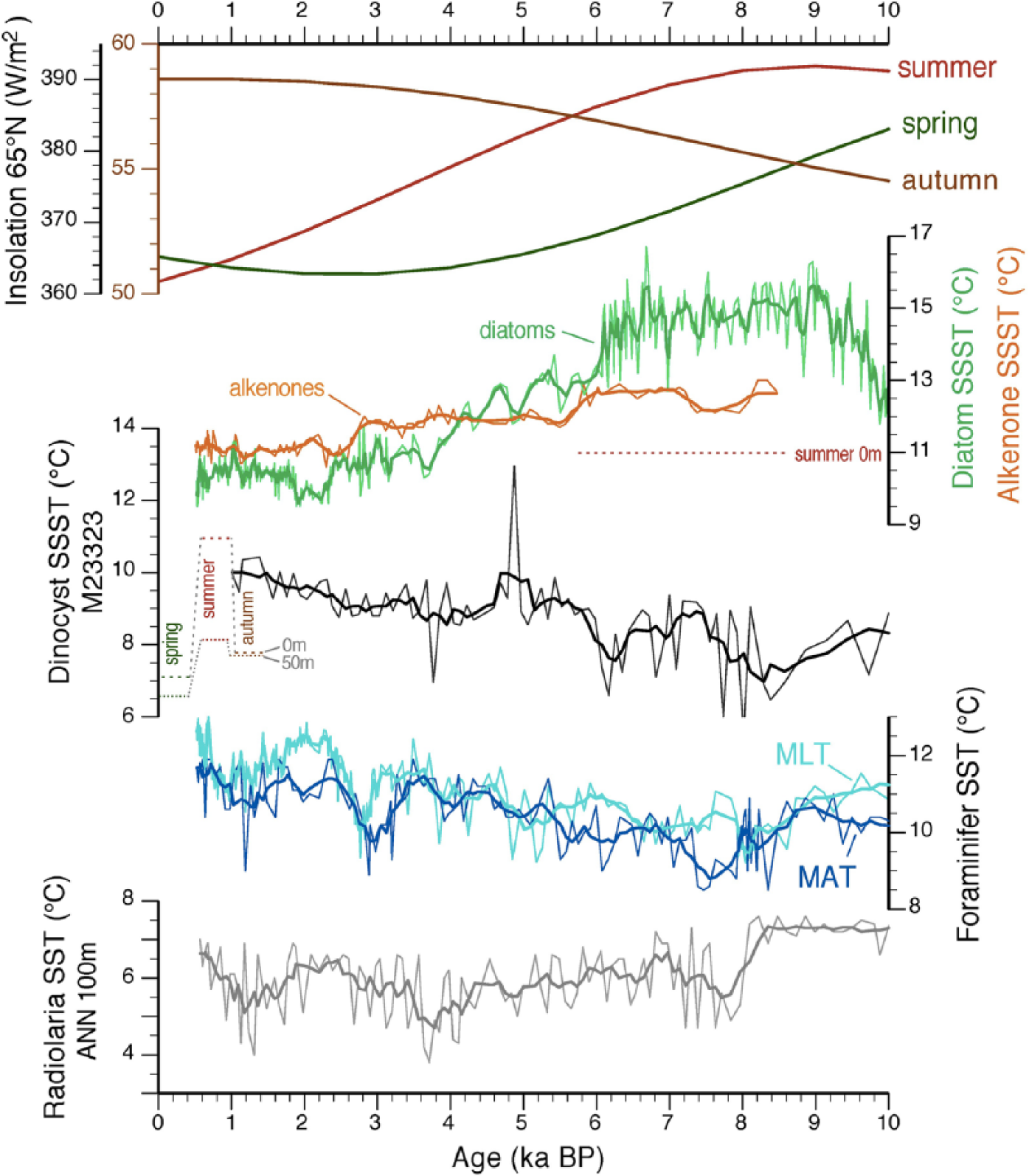
Comparison of reconstructed plankton-based (summer) temperatures at the Vøring Plateau, from dinoflagellate cyst assemblages in core M23323, and coccolithophores (alkenones; Calvo et al., 2002), diatoms (Berner et al., 2011; Birks and Koç, 2002), planktic foraminifera (Andersson et al., 2010; Risebrobakken et al., 2003), and radiolaria (at 100 m water depth; Cortese et al., 2005) in core MD95-2011, against spring, summer, and autumn Holocene insolation at 65°N (computed with ANALYSERIES 2.0 (Paillard et al., 1996) using the Laskar et al., 2004, data). Thicker lines represent a 5-point smooth of the original data. Also shown are the modern spring, summer, and autumn SST at 0 and 50 m water depths over the area of the two core sites (Conkright et al., 2002; Stephens et al., 2002). Note that the original age models of each respective record have been recalibrated to conform to the age model of core M23323 used here.
Mid-Holocene
The drastic change from assemblages dominated by Nematosphaeropsis labyrinthus, and also by Impagidinium pallidum in the ArD-PD, to assemblages dominated by Operculodinium centrocarpum that marks the onset of the mid-Holocene interval (Figures 2 and 3) suggests a major surface current regime shift across the entire Nordic Seas. The timing of this event, between ~6.1 and 7.5 ka BP, is consistent in all cores despite the low temporal resolution and sedimentation rates of most cores. In the low-resolution cores, the transition generally is more gradual, a likely consequence of the bioturbational smoothing of the event, both upward and downward. The high-resolution core M23323, therefore, can be assumed to capture the timing more adequately (Bard, 2001). It shows that the transition occurred around 6.1 ka BP and that the regime change was abrupt. Despite lower time resolution, core PS1247 from the southern Greenland Sea duplicates the shift and timing of high-resolution core M23323 (Figure 3a). The apparent offset of more than 2 ka at sites M23062 and M23480, where the Holocene section is less than 15 and 27 cm, respectively (ES Figure A, available online), is not necessarily significant as important size-selective bioturbation mixing (Bard, 2001; Shull and Yasuda, 2001) in addition to possible variations in sedimentation results in large uncertainties with regard to the chronostratigraphy. Indeed, it is hard to imagine a plausible oceanographic scenario that would explain a >2 ka offset of events for those two cores with respect to the consistent signal from the other sites across the Nordic Seas. On the contrary, the slightly earlier occurrence of the assemblage shift in the two southernmost cores M23411 and HM52-43 seems to represent a real indication for a time-transgressive northward latitudinal change.
The distribution pattern of Operculodinium centrocarpum in relation to secondary and subordinate species after the mid-Holocene shift is close to what is observed in the surface sediments (Rochon et al., 1999) and can thus be related to the onset of a modern type of surface circulation (Baumann and Matthiessen, 1992). Similarly, diatom assemblages contain a dominant signature of typical ‘modern’ water masses at the Vøring Plateau after ~6 ka BP (Berner et al., 2010). The interval corresponding to and shortly after the major dinocyst shift falls at the very end of the broad Holocene ‘hypsithermal’ period, which appears to be reflected by the subtle abundance maximum Spiniferites mirabilis at the Vøring Plateau. This species is the most thermophilic in the assemblages and the Vøring Plateau is at the northern limit of its modern distribution (e.g. Penaud et al., 2008). Considering the ecological preferences of Spiniferites mirabilis and the strong Atlantic signature of the eastern assemblages, the short interval from ~6 to 4.5 ka BP can, therefore, be regarded as the ‘Atlantic’, fully marine surface ocean optimum.
The mid-Holocene also seems to have been the time when Atlantic water was admixed farthest westward into the surface waters, as suggested by high percentages of Operculodinium centrocarpum, whose modern-day zone of absolute dominance (>60%) clearly delineates the main body of the propagating North Atlantic water mass (Grøsfjeld et al., 2009; Rochon et al., 1999). Although some cores from the ArD-PD display a slight temperature increase at this time (Figure 5; ES Figure B, available online), the dominance of Operculodinium centrocarpum should primarily be interpreted as a water mass fingerprint. Indeed, planktic foraminiferal assemblages would rather suggest cooling in the subsurface waters of the ArD-PD shortly after the mid-Holocene transition (Bauch et al., 1999), perhaps indicating (the initiation of) downward convection of strongly winter-cooled saline surface waters. In this respect, the timing of the mid-Holocene transition in the dinocyst assemblages coincides broadly with the final stages of the deglacial Laurentide and Greenland Ice Sheet volume loss (e.g. Carlson et al., 2008; Lecavallier et al., 2014; Occhietti et al., 2011) and the onset of deepwater formation in the Labrador Sea (de Vernal and Hillaire-Marcel, 2006; Hillaire-Marcel et al., 2001). In the absence of a manifest change in the reconstructed sea surface parameters around the transition (Figure 5; ES Figure B, available online), the complete dinocyst assemblage changeover is likely the reflection of the modification of the northern hemisphere surface water properties and circulation in response to the decrease in total Atlantic Northern Hemisphere freshwater input and various associated feedbacks (cf. Renssen et al., 2009). Consequently, the modern circulation was initiated, with ‘spillover’ of warm and saline North Atlantic Drift waters from south of the Iceland–Scotland Ridge – typified by high percentages of Operculodinium centrocarpum from early on in the Holocene (Harland and Howe, 1995) – feeding into the Nordic Seas (Rossby, 1996).
Late Holocene
The millennial-scale late Holocene cooling in the high-latitude North Atlantic (e.g. Wanner et al., 2008) resulted in a reduced westward intrusion of Atlantic waters and an eastward migration of the oceanic Polar and Arctic fronts. Slightly higher shares in the subordinate assemblages of Nematosphaeropsis labyrinthus and cysts of Pentapharsodinium dalei, indicative for the modern Arctic front and modified East Icelandic Current (EIC; Figure 1), respectively (Rochon et al., 1999), reveal an increased contribution of cold and fresher EGC-derived water masses in the ArD-PD (Figure 3). Although the MAT reconstructions do not show a significant increase in seasonal SIC (Figure 5), the occurrence of Islandinium minutum (de Vernal et al., 2013) suggests that the EGC brought drift ice to the area more frequently (cf. section YD – earliest Holocene). Alternatively, the increased abundances of heterotrophic taxa might also be related to the more recent deposition and, therefore, shorter oxygen exposure time of these cysts, which, by consequence, have not (yet) been affected by oxygen-induced degradation to the same degree as the ‘older’ assemblages (Zonneveld et al., 2008). While the (quasi-)exponential decrease of the heterotrophic taxa downward from the top of the low-sedimentation cores (Figures 2 and 3) indeed would seem to be a typical signature of diagenetic bias, this alternative explanation is hard to prove with the available data and would require the analysis of additional aerobic degradation indices (cf. Bogus et al., 2012).
The absolute timing of the late Holocene circulation shift again is difficult to evaluate due to the low sedimentation rates and uncertainties with the chronology. Planktic foraminiferal data from the central Nordic Seas reveal increased polar water masses there from 5 ka BP onward (Bauch and Erlenkeuser, 2008), and the eastward migration of the Arctic front has been estimated to occur between 5 and 3 ka BP based on diatoms (Koç et al., 1993). Evidence from the eastern Greenland shelf in Denmark Strait and off northern Iceland would further suggest an important eastward deflection of EGC waters (Solignac et al., 2006), thus giving a more polar character to the EIC as suggested by the dinocyst data.
The late Holocene assemblage change is observed around 2.4 ka BP in core M23323 from the Vøring Plateau (Figure 2a), a timing similar to the regional cooling recorded by diatom and alkenone records (Berner et al., 2011; Calvo et al., 2002; Marchal et al., 2002) and to the decrease of the Atlantic water indicator Operculodinium centrocarpum on the North Iceland shelf (Solignac et al., 2006). The transition at about 2.4 ka BP in the east appears to reflect a delayed reaction of the Atlantic-sourced water inflow to the gradual expansion of the ArD-PD. Overall, the recurrence of cool dinocyst taxa in the subordinate assemblages (notably Nematosphaeropsis labyrinthus, Impagidinium pallidum, and cysts of Pentapharsodinium dalei) illustrates the basin-wide late Holocene sea surface cooling. The surprising absence of a significant cooling reconstructed from MAT will be addressed in the discussion below.
Comparison of micropaleontogical records of surface ocean conditions
The dominance of cool species in the dinocyst assemblages prior to 6.5 ka BP in the AtD is at odds with the early to mid-Holocene Climatic Optimum associated with the Northern Hemisphere summer insolation maximum (e.g. Jansen et al., 2008). The contrast between the dinocyst and planktic foraminiferal assemblages, which show low abundances of the polar species Neogloboquadrina pachyderma (s) roughly between 10 and 6.5 ka BP and minima as early as 11 ka BP (e.g. Bauch and Erlenkeuser, 2008; Hald et al., 2007), can be related to the differing habitat depth of both plankton groups in combination with a decoupling of the surface and deeper layer seasonality and climate response. The differing autecology of zooplankton and phytoplankton, notably diatoms and coccolithophores, is also the likely explanation for contrasting trends in the respective SST reconstructions (Figure 6), as extensively discussed elsewhere (Andersson et al., 2010; Cortese et al., 2005; Hass et al., 2001; Leduc et al., 2010; Risebrobakken et al., 2011).
Given the light dependency of phototrophic dinoflagellates, which produce most of the dinocyst taxa fossilized in sediment, one could expect that they will show a trend similar to other phytoplankton tracers. However, such is not the case. The diatom paleotemperature records from the Vøring Plateau and westernmost Barents Sea shelf show a clear response to insolation with maximum SSTs between ~9 and 6 ka BP (Andersen et al., 2004; Berner et al., 2011; Birks and Koç, 2002; Calvo et al., 2002; Koç et al., 1993; Marchal et al., 2002), and the reconstructions from coccolithophore-derived alkenones show a slight but significant trend of decreasing temperature since about 8 ka BP. By contrast, the dinocyst assemblages and their MAT reconstructions show a very different signal (Figure 6; ES Figure B, available online). Inasmuch as the respective tracers were demonstrated to relate to temperature, the differences between the dinocyst SST reconstructions in core M23323 and the SST reconstructions from alkenones and diatoms in nearby core MD95-2011 have to involve the actual temperature-related parameter that is captured by each tracer. Possible factors steering the SST signal will now be explored.
The early Holocene, that is, prior to ~6 ka BP, features lowest Holocene dinocyst-based estimates of summer SSTs (Δ = −2°C relative to <3 ka BP) but a plateau of maximum temperatures in the alkenone (Δ = +1.5°C) and diatom estimates (Δ = +5°C; Figure 6). Part of this explanation could be that diatoms and coccolithophores live even closer to the surface, where the energy uptake from insolation is highest. In addition, the early-Holocene diatom-based estimates exceed alkenone-based SSTs by ~2°C and modern summer SST at the Vøring Plateau by over 4°C (Conkright et al., 2002; Stephens et al., 2002). As such, it can also be argued that the exceedingly high diatom temperature estimates are also a result of increased productivity of photosensitive species under higher luminosity, rather than a sole reflection of temperature (e.g. Lionard et al., 2005; Saito and Tsuda, 2003; Smayda, 1969). With decreasing summer insolation, diatom temperature estimates drop sharply around ~6 ka BP and compare with the range of alkenone-based temperatures from then on.
Another likely contributing factor is a difference in the seasonal growing behavior of each primary producer. Comparing the spring and summer insolation curves reveals that the diatom record from the Vøring Plateau most closely follows the declining trend of spring insolation, while alkenone-based temperatures rather follow the delayed declining trend of summer insolation in a stepwise manner (Figure 6). The early spring bloom in the subarctic is usually dominated by diatoms, while coccolithophores mainly grow during summer (Andruleit, 1997; Matthiessen et al., 2005; Zondervan, 2007, and references in the latter two). Dinoflagellates are never abundant during the spring bloom, and the presence of diatoms would notably favor the growth of heterotrophic dinoflagellates, which feed on diatoms (Gaines and Elbrächter, 1987; Taylor and Pollingher, 1987). The diatom spring bloom is usually followed by a (phototrophic) dinoflagellate summer and/or autumn bloom (Matthiessen et al., 2005, and references therein). Low dinocyst concentrations during the early Holocene suggest rather modest dinoflagellate productivity, possibly related to a nutrient-depleted water column at the end of the summer. Such seasonal succession in dinocyst species production could explain the sedimentary subordinate coexistence of food-dependent heterotrophic (Brigantedinium spp.) and more oligotrophic (-tolerant) oceanic taxa during the early Holocene (Impagidinium spp.; Marret and Zonneveld, 2003) and, to some degree, the low temperature estimates that rather vary around modern surface temperature for late spring and autumn (Figure 6). After ~6 ka BP, dinocyst-based temperature estimates are slightly higher, and the SST trend combined with decreased abundances of heterotrophic species would hint toward an increased weight of the summer or autumn bloom.
The reasoning above assumes that cores MD95-2011 and M23323 have registered the same climate due to their proximity (~150 km). However, the modern surface current configuration around the Vøring Plateau is highly complex, with a bathymetry-controlled decoupling of the main water mass pathways and variable westward admixture of the seasonally variable Norwegian Coastal Current (e.g. Saetre, 1999). Hence, conditions can be assumed to have been more unstable at site MD95-2011 relative to site M23323 located further offshore. The late Holocene increase of reconstructed temperatures based on dinocysts despite the recurrence of cool taxa in the subordinate assemblages can in part be attributed to the more open-oceanic position of M23323. Indeed, the poor, nearly monospecific assemblages from the mid-Holocene are typically found today far offshore in the cooler regions of the Norwegian Sea, while the modern analogues to the more diverse late Holocene assemblages are mostly from the Vøring Plateau.
A seasonal bias between phytoplankton proxy records is generally less marked in the PD, where unfavorable conditions can delay the growth initiation until as late as July or even August (Matthiessen et al., 2005). This results in a more harmonious signal between the different biogenic tracers, possibly reflecting the conditions over a shorter growing season, also marked by lower amplitude of temperature. Both alkenones (Marchal et al., 2002) and the MAT reconstructions from dinocysts indicate generally (slightly) higher temperatures in the PD from 9 to 6 ka than during the ensuing period (Figure 5), reflecting a primary response to elevated summer insolation. While pelagic, benthic, and sedimentological records indicate the presence of Atlantic water on the Iceland shelf (Andrews and Giraudeau, 2003; Eiríksson et al., 2000), the early Holocene dinocyst assemblages as well as coccoliths and radiolaria (Andruleit, 1995; Schröder-Ritzrau, 1994) in the ArD-PD to the north still contain a dominant polar water mass component, revealing that any inflow of Atlantic water through the Irminger Current remained confined to the northern Icelandic shelf. The important share of typical cold EGC elements in diatom assemblages from the outer north Icelandic shelf seems to confirm the prominent continuing early Holocene contribution of these cold water masses in the ArD-PD (Justwan et al., 2008).
Conclusion
The compilation of Holocene dinocyst records along an E-W transect across the southern Nordic Seas illustrates the utility of census and MAT dinocyst analysis for paleoceanographic reconstruction at high latitudes, offering complementary information to other biogenic tracers. As for all proxy records, a number of caveats have to be taken into account when working in the Nordic Seas, which typically result from the low sediment accumulation rates. Next to the increased uncertainty of the absolute timing of events due to higher bioturbational smoothing, the low sedimentation rates may also affect the assemblage composition because of longer exposure of cysts to more-oxygenized conditions, especially when productivity was low. Nonetheless, a consistent picture of Holocene surface water evolution emerges from the compilation of low-resolution cores and reflects the major trends revealed by a high-resolution core from the Vøring Plateau. Four intervals can roughly be distinguished in the studied core sections:
A transitional phase from the YD into the earliest Holocene, characterized by significant meltwater input and likely strong seasonal contrast in sea surface conditions. This transitional phase occurs around 12–11 ka BP in the AtD and appears to have persisted 1–2 ka longer in the colder western domains;
An early Holocene interval featuring a dominance of cold-temperate dinocyst species despite high boreal summer insolation at that time. This suggests a response of dinoflagellates to season-dependent environmental changes across the growing season that is different from that of other (phyto)plankton, which generally show a SST maximum at that time. The registered differences between proxies tends to be more pronounced as possible favorable conditions occur more spread-out through the year;
A marked basin-wide shift in assemblage composition, from the dominance of Nematosphaeropsis labyrinthus, and Impagidinium pallidum in some western cores, to the dominance of Operculodinium centrocarpum, between 6.1 and 7.5 ka BP marks the transition to the mid-Holocene. It is interpreted as the onset of a modern-type of surface circulation with strong northward spreading of Atlantic water masses. The signal appears to be a dominant water mass fingerprint, as no drastic changes are seen across the event in the quantitative reconstructions. Despite some uncertainty with the dating, the assemblage transition might have occurred slightly earlier closer to the Iceland-Scotland Ridge, suggesting this event to be latitudinally time-transgressive. The ‘warmest’ Holocene assemblages occur at the Vøring Plateau shortly after the transition, when the Atlantic waters also seem to have spread farthest westward;
Finally, the uppermost core sections are characterized by the return of colder elements in the assemblages, in line with the late Holocene cooling and/or enhanced contribution of EGC waters. This change occurs from ~2.4 ka BP at the Vøring Plateau and probably earlier in the west.
Footnotes
Acknowledgements
We thank the teams from the Leibniz Laboratory for Radiometric Dating and Stable Isotope Research (Kiel University) and the Stable Isotope Laboratory at GEOMAR (Kiel) for their support in the radiocarbon dating and isotope analyses. We are grateful to Maryse Henry (Geotop-UQAM) for assistance with the palynological preparation. Helpful reviews were provided by Kari Grøsfjeld and one anonymous reviewer.
Funding
This work was supported by the German Research Foundation (Grant NI1248); the European Union’s Seventh Framework program (Grant Agreement 243908 ‘Past4Future’: climate change – learning from the past climate); the Ministère du Développement Économique de l’innovation et de l’exportation du Québec; and ArcTrain-Canada.