
Abstract
This study presents the results of a multi-proxy analysis conducted to improve our understanding of the palaeoenvironmental conditions of the freshwater and brackish marshes of Cal Maurici (Barcelona, Spain) and the human impact on them during the mid-Holocene (6171–3891 cal. yr BP). The study integrates data from pollen, phytolith, diatom, charcoal, seeds and malacological analyses and helps to reconstruct the ecological conditions during the early establishment of farming communities in western Mediterranean facade. The results indicate a landscape dominated by Mediterranean vegetation with aquatic plants in the shallow marshes and well-developed forests in the nearby area, providing for the first time in the Holocene of NE Iberian Peninsula the palaeoecological conditions of deltaic areas. Ecofactual evidence indicates an initial landscape dominated by brackish marshes (6171–5773 cal. yr BP) in which Ruppia cf. maritima was predominant and human impact was low. Between 5026 and 4839 cal yr. BP, freshwater conditions expanded with an increase in Potamogeton sp. and the presence of Typha angustifolia and Spirogyra sp. algae with well-developed oak woodlands and deciduous trees in nearby areas. The expansion of evergreen forest occurred later (from 4960 to 4825 cal. yr BP until 3712 cal. yr BP), with the decline of deciduous woodland and the expansion of evergreen oaks, pinewoods, wild olive trees and box, coinciding with a period of increased human activity in the area. Additionally, the presence of marine resources at several archaeological excavation sites and domestic plants at Cal Maurici provides an opportunity to evaluate the interaction between earlier farmers and marine or deltaic ecosystems.
Introduction
Coastal ecosystems, in particular large deltas and wetlands, have attracted human populations in the past and are known to have played an important ecological and economic role. Archaeological excavations and sampling programmes carried out in these zones have led to the development of what has come to be known as coastal or wetland archaeology, the study of the interactions between ancient communities and coastal ecosystems (Louwe Kooijmans, 1993; Stanley et al., 1999; Van Neer et al., 2013). Indeed, the waterlogged conditions present in wetland areas are ideal for organic preservation, due to the decrease in the gas exchange rate between sediments and the atmosphere, and so they provide excellent opportunities for research (Brix, 1990).
Prehistoric communities were not totally unfamiliar with climatic and environmental changes in the landscape, where the natural dynamics, controlled by several geologic processes, could be rapid. This relationship between natural processes and human activities during the Holocene has long attracted the attention of archaeologists (Anthony et al., 2014; Berger and Guilaine, 2009). Holocene sea-level fluctuations (Zazo et al., 1994) altered the shores and hence emphasize the need to reconstruct past environments so that we might gain a fuller understanding of this relation between Prehistoric communities and littoral ecosystems. Today, the mid-Holocene and the Neolithic and Mesolithic landscapes of deltaic areas lie buried below current soils; however, they would have been important landscapes for the last foraging and early farming communities as they offered coastal, marine and terrestrial resources as well as rapid transit up and down the coastline and inland up the river.
The presence of marine elements, documented at coastal sites, provides direct evidence of littoral exploitation and procurement (Rainsford et al., 2014). At prehistoric sites, this exploitation of the littoral was an important factor in the economic organization of human groups. Stable isotope analysis has revealed that marine resources formed part of the human diet in the Iberian Peninsula and Mediterranean area during the Mesolithic (Mannino et al., 2011; Salazar-García et al., 2014) and perhaps, albeit in smaller quantities, during the Neolithic and Bronze Age too (Lightfoot et al., 2011; Salazar-García et al., 2013). There is also evidence that elsewhere in Europe salt marshes were exploited through husbandry practices (Britton et al., 2008). Additionally, marine elements can be traced far inland from the coast, signalling mobility, exchange or interaction between different groups over large territories (Chapman, 2008).
Evidence of human impact on the surrounding vegetation during the process of neolithization is frequently presented in environmental archaeology, being characterized primarily by a decrease in the number of arboreal plants and an increase in anthropic indicators (Badal et al., 1994; Innes et al., 2013; Kalis et al., 2003). Several studies have focused on north European wetland areas, estuarine and costal environments rich in archaeological sites from that period (Crombé et al., 2015; Out and Verhoeven, 2014; Paalvast and van der Velde, 2014). Over the last few decades, the number of Holocene palaeoenvironmental records in the Atlantic facade of the Iberian Peninsula, which are particularly rich in pollen sequences (Carrión et al., 2010; Fletcher et al., 2007; Jalut et al., 2000), has increased considerably.
Today’s littoral plains of Barcelona and the Llobregat delta (NE Iberian Peninsula) constituted a coastal wetland area with several marshes and lagoons (Gámez, 2007; Marquès, 1974, 1984; Riera, 1995) during the Holocene. Several Neolithic to Bronze Age sites located around this region have yielded a number of marine elements signalling a strong interaction between early farming communities and marine ecosystems. Despite the need to evaluate the ecological conditions of this human procurement, our knowledge of past littoral ecosystems is poor in this area with no multi-proxy analyses having been conducted to date. To rectify this, our archaeological study, conducted on several test pits at Cal Maurici, provides a unique opportunity to examine the environmental conditions during the Neolithic period and to identify foraging areas. Stratigraphic and ecofactual analyses have been carried out on site samples dated between the 6th and 4th millennia cal. yr BP in order to depict the complex palaeoenvironmental evolution undergone by the area of the Llobregat delta.
Cal Maurici
Cal Maurici (41°20′15.54″N, 2°3′24.26″E) lies in the trench of Module 2-Second Section (~1750 m) of the Madrid–Barcelona high-speed train network between location markers PK 100+560 and PK 102+300, in the municipality of Sant Boi de Llobregat (20 km W of Barcelona). In this area, railway tunnelling (Figure 1b and 1c) has exposed the uppermost units of the Llobregat delta complex along a stretch of the trench measuring 25 m in width and 11 m in depth.

Cal Maurici: (a) location of Cal Maurici and prehistoric sites yielded marine resources around the Llobregat delta; (b) the trench of Module 2-Second Section of the high-speed train network between Madrid and Barcelona in the municipality of Sant Boi; (c) location of Cal Maurici profiles.
The region occupies the meso- and thermo-Mediterranean environment, composed primarily of evergreen oak forest (Quercion ilicis) with the occurrence of small communities of deciduous oak occupying northern slopes and areas in shadow. To the south, the presence of calcareous rocks has favoured the development of thermo-Mediterranean communities (Oleo-Ceratonion) characterized by the presence of Quercus coccifera, Olea europaea, Pistacia lentiscus, Pinus halepensis and Chamaerops humilis.
Cal Maurici is located in the apical area of the Llobregat delta (Figure 1a), a plain with an ~23-km shoreline, occupying an area of ~97 km2. The source (les fonts) of the Llobregat lies in the Pyrenees under an equinoctial rainfall pattern. The delta, characteristic of micro-tidal seas, is formed mainly by detrital material from the Pliocene to the Holocene and consists of four delta sequences, three of which are currently submerged (Gámez et al., 2009).
The general architecture of the Holocene delta is widely described and consists of several lithological units (Gámez, 2007; Marquès, 1974, 1984), and the thickness of the most recent deposits ranges from ~20 m in the delta apex to ~60 m at the current coastline where the river feeds into the Mediterranean sea (Gámez, 2007; Lafuerza et al., 2005; Manzano, 1986; Marquès, 1974, 1984). Several earlier studies have attempted to reconstruct the millennia-scale shoreline changes of the Llobregat delta during the Holocene on the basis of historical and archaeological data (Gámez, 2007; Marquès, 1984). Other studies based on palynological analyses have shown some sporadic human clearance without qualitative and/or quantitative changes inside the natural woodland, except for slight oscillations in arboreal pollen (AP) values (Riera and Esteban, 1994). These clearances are synchronous with Neolithic and Bronze Age settlements.
The archaeological record of this area indicates that it was occupied continuously between the early Neolithic and late Bronze Age (ca. 7400–2900 cal. yr BP). The earliest settlement coincides with the rapid spread of the Cardial culture (ca. 7400–6500 cal. yr BP). Later, the area was occupied during several phases of the middle (ca. 6500–5200 cal. yr BP) and late Neolithic (ca. 5200–4200 cal. yr BP), as illustrated by the ongoing activity in the Can Tintorer mines (Gavà, Spain) (Borrell et al., 2015). The archaeological record from the nearby site of Cova de Can Sadurní (Begues, Spain) shows a continuous occupation between this former period to the Bronze Age (3800–2800 cal. yr BP) (Blasco et al., 2011).
The presence of marine elements at these archaeological sites (Figures 1 and 2) indicates that the communities inhabiting these coastal areas made use of coastal resources (Bordas et al., 2013; Bosch et al., 1999; Lloveras et al., 2014). The practice of these activities is further supported by the recovery of bones belonging to fish from the Sparidae family in the mines of Can Tintorer (Bosch et al., 1999) and abundant shell remains in the open-air, prehistoric sites in Barcelona (Lloveras et al., 2014). Of particular note is the recovery of a conch (Charonia nodifera), interpreted as a shell trumpet (Villalba et al., 1986) and stone fishing sinkers (Bordas and Salazar, 2006; Carlús and González, 2008; Villalba et al., 1986). Marine elements have also been identified in the inland territories, as grave goods or functional elements, and are indicative of transport from coastal areas over short (~20 km), medium (~50 km) and long distances (~100 km). Ornaments made from a range of marine raw materials, including natural polished shells, gastropod pendants, and shell beads, are the most commonly occurring (Blasco et al., 2011; Oliva, 2012).

Prehistoric shell remains from Neolithic and Bronze Age sites around the Delta del Llobregat – (a) Cardium sp. and Glycymeris sp. from Cova de Can Sadurní (Photo: E Galardi; CEB-CIPAG, 2009); (b), (c), and (d) bivalve fragments, Columbella rustica and Dentalium sp., from Cova Bonica; (e): pectinid fragments of Reina Amàlia (AACC-MUHBA: Arxiu Arqueològic del Centre de Col·leccions del Museu d’Història de Barcelona); (f) gastropod fragments from Can Sadurní and Cova Bonica (Photo: E Galardi; CEB-CIPAG, 2009); (g) Dentalium sp. from Coveta del Marge del Moro (Photo: E Galardi; CEB-CIPAG, 2009); (h), (i), (j), and (k) Ostrea sp., Patella sp., Acanthocardia sp. and Glycymeris sp. of San Pau del Camp (AACC-MUHBA); (l) Glycymeris sp. of Conservatori del Liceu (AACC-MUHBA); (m) Charonia nodifera interpreted as a shell trumpet and stone fishing sinker from Can Tintorer (Museu de Gavà); (n), (o), and (p) stone fishing sinkers from Can Tintorer (Museu de Gavà), Carrer Riereta 37-37b (MUHBA), and Reina Amàlia (MUHBA). Scale: 1 cm.
Thus, the study of these marine elements should be able to provide us with considerable information about the interaction between past communities and the coastal landscape through the procurement of natural resources.
Materials and methods
Excavation methodology
Fieldwork was carried out during 2004 and 2005 following standard geological and archaeological methods. The site was excavated using a power shovel and eight profiles were established (Figure 1b and 1c) lying at an altitude between 0 and −4 m below mean sea level (b.m.s.l.) along a 54-m stretch of the west face of the railway trench. Profile-B represents the main stratigraphic sequence, from which most of the samples were collected. It is 3.9 m thick, 4 m wide and is divided into 11 layers. Profile-C lies at a distance of 33 m from profile-B and corresponds to the southernmost test pit, that is, the one closest to the present-day shoreline. A total of 30 layers have been identified in this profile, in a sequence that is 2.4 m thick and 2 m wide. At a distance of 7 m from profile-C lies profile-E, with a 1.3-m-thick and 0.6-m-wide exposed profile that has been divided into 18 layers. Profile-F is 5 m north of profile-E and is 2.1 m thick, 0.7 m wide and has 13 layers. Profile-G lies 12 m from profile-F and is 1.2 m thick and 0.9 m wide and has 17, mostly peat, layers. Profile-H lies between profile-G and profile-B and presents a thickness of 1 m, a width of 0.9 m and 9 layers. Profile-I is the northernmost sequence and is located 15.4 m from profile-B and 54 m from the southern edge. The profile is 2.4 m thick, 0.8 m wide and has been divided into 10, mainly sand, layers.
Henceforth, the layers, which are independent in each trench, are referred to using Roman numerals and are correlated across the different site profiles and grouped into five facies.
Sampling and laboratory methods
A total of 109 samples were collected during the excavation work as part of the multi-proxy analysis. These samples were recovered along the sequences of the two main vertical profiles, B and C, which correspond to the northernmost and southernmost columns. Other profiles (E, F, G and H) were not sampled as they are intermediate profiles between the main profiles and present the same geological record. Sampling was completed along profile-I, marking the edge of the wetland area.
Where possible, all the facies were sampled and if not an explanation is provided in Table 1. Even in those instances in which the sampling provenance corresponded to the same layer, facies or laboratory method, each sample was labelled independently (#1 to #109). Of these samples, 24 (#1 to #24) were collected for sediment analysis and stratigraphic description, 4 organic sediment samples and 1 bone remain were taken for 14C dating (#25 to #29), 11 (#30 to #40) for pollen analysis, 14 (#41 to #54) for phytolith and diatom analyses, 4 (#55 to #59) for charcoal analysis, 16 (#60 to #75) for seeds and fruits analysis and 44 (#76 to #109) for malacological identification.
Stratigraphic correlation summary and sampling.

Not material recovered or indeterminate wood (peat).
Scarcity of material and statistically insignificant.
″Sands with washed and transported ecofactual elements.
′Not dated because upper and lower facies are in the same range.
Stratigraphy and sedimentology
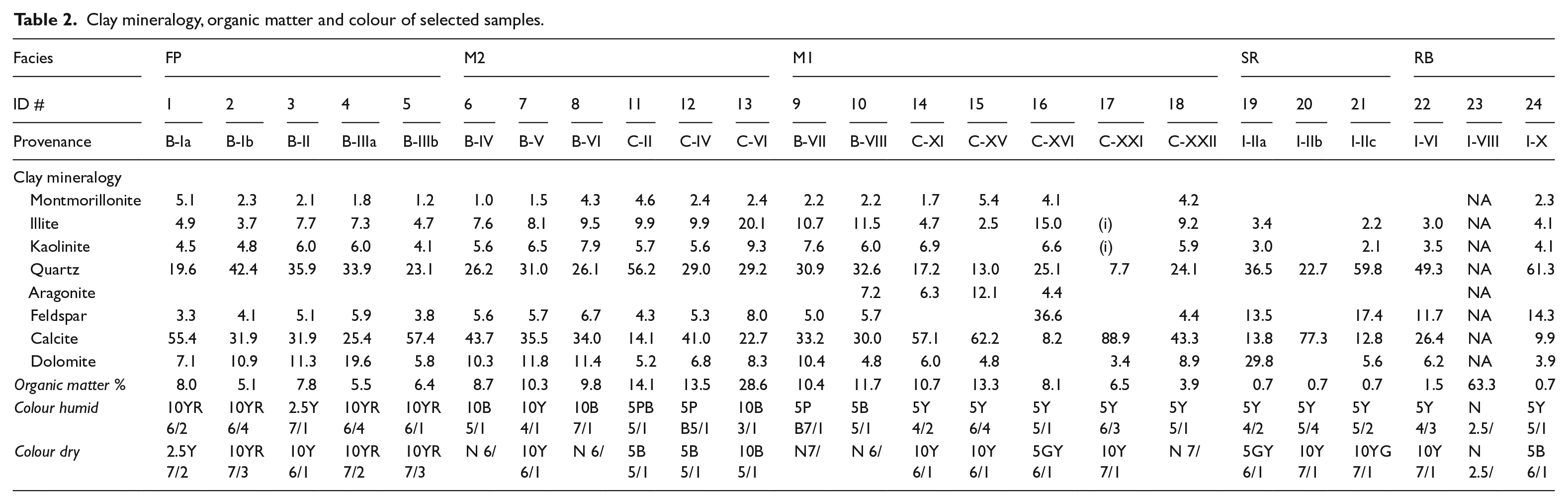
The stratigraphic description follows Schoeneberger et al.’s (2002) field book guidelines, while sample collection focused on the vertical profiles. After air-drying for 24 h, a total of 24 samples (#1 to #24) were colour-described (dry and humid) using the MUNSELL© soil colour chart and then subjected to grain size and mineralogical analyses. Particle size distributions were determined using a Beckman Coulter-LS™ analyser. Cumulative curves, frequency histograms and summary statistics, including statistical parameters such as mean, sorting, kurtosis and skewness, were calculated from these datasets following the geometric (graphical) method of Folk and Ward (1957) by means of the Gradistat ver. 8 software (Blott and Pye, 2001).
Mineralogy was determined with a Siemens© D-5000 x-ray diffractometer (XRD), using randomly oriented powder from the bulk samples. Samples were pre-treated with H2O2 to remove organic matter. Replicates were heated to 375 or 600°C for 1 h or treated with ethylene glycol at 60°C to differentiate between clay minerals. Selected samples were analysed by energy dispersive x-ray spectrometry (EDXS) (Bruker© X-Falsh Detector 4020) or observed by scanning electron microscopy (SEM) (Hitachi© S-3400N). Semi-quantitative mineral analyses were based on the peak areas obtained using EVA© ver. 7.0 software.
Dating
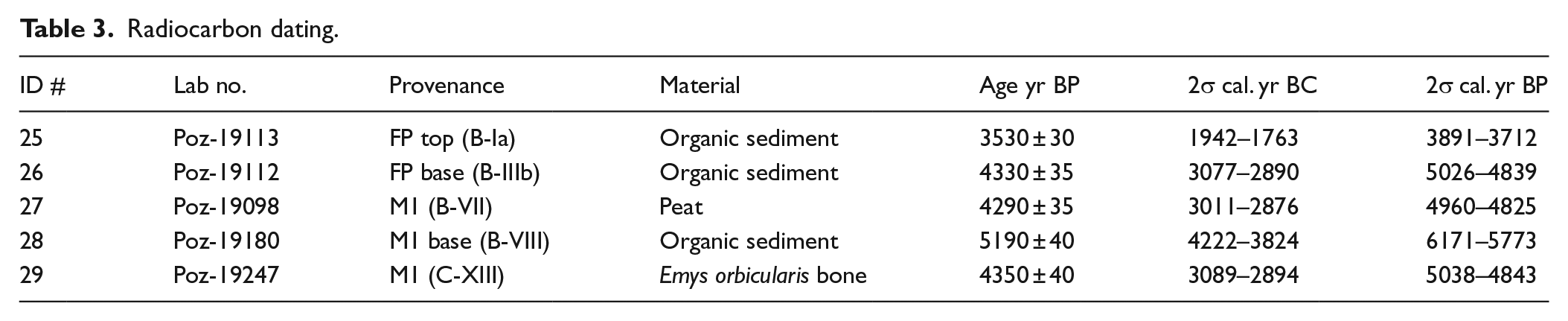
Five samples were selected for accelerator mass spectrometer (AMS) dating at Poznań Radiocarbon Laboratory and then calibrated using IntCal 13 curve (Reimer et al., 2013) and OxCal v4.2 software (Bronk Ramsey, 2009). Of these, three samples correspond to organic sediment, one to peat sediment from profile-B and one to turtle bone from profile-D.
Palaeobotanical remains
Palaeobotanical analyses were conducted on pollen, phytoliths and archaeobotanical macroremains (charcoal, seeds and fruits). Pollen analyses were conducted on profile-B and the sediments were treated following standard procedures (Faegri et al., 1989). The pollen sum used to calculate percentages excludes local pollen assemblage, which includes aquatic and hygrophilous taxa such as Cyperaceae, Typha angustifolia type and Myriophyllum.
Phytolith analyses were conducted on profile-A and profile-B following the rapid extraction procedure described in Katz et al. (2010). Phytoliths were counted and identified in random fields at 200× and 400× using an optical microscope (Olympus BX41). Morphological identification was based on our modern plant reference collection (www.phytcore.org) and standard literature (Mulholland and Rapp, 1992; Piperno, 1988, 2006; Twiss et al., 1969). The International Code for Phytolith Nomenclature (ICPN) was also followed whenever possible (Madella et al., 2005).
Seeds and charcoal were both obtained using the soil flotation technique, the former with 174 L of sediment from profile-B and profile-C and the latter with 72 L of sediment from profile-B and profile-I. For the flotation, we used 5-, 1- and 0.5-mm meshes and charcoal analyses were based on >5 mm fraction while the >2 mm fraction was dried, sieved and sorted from the smaller (1 mm) fraction.
The charcoal study was based on the analysis of 376 remains from profile-B and profile-I. Charcoal remains were identified using an optical microscope with reflected light (Olympus BX41) at 5×, 100×, 200× and 500×. Each piece was fragmented by hand in order to obtain fresh cuts of the three anatomical sections that characterize wood cell anatomy. The identification process was based on the observation of the wood anatomy and supported by Schweingruber (1990) wood anatomy atlas as well as a reference collection of Mediterranean woody species.
A total of 1746 seed remains were obtained from profile-B and profile-C. Fragments were not counted when the germ was not preserved. Determination focused on species and, alternatively, on genus when species identification was not possible and followed the criteria outlined in several anatomical atlases (Jacquat, 1988; Knörzer, 1970; Montégut, 1970; Schoch et al., 1988). Furthermore, the relative presence and abundance of plant species were determined by calculating ubiquity measures and/or standardizing specimen counts against soil volume (Popper, 1988). The methodological analysis was conducted following standard procedures of seeds analysis at prehistoric sites.
Malacology
Malacological sampling of molluscs, foraminifers, charophytes and ostracods was conducted on profile-B and profile-C. Additionally, several macro-molluscs were randomly collected from layers XIII and XVIII from profile-C, to complete previous sampling at the site. Thus, we recovered both macro- and micro-molluscs. Morphological identification was based on private malacological collections and on the standard literature (Barash and Zippora, 1992; Bech, 1990; Giannuzzi-Savelli et al., 1997, 2001; Graham, 1988; Haas, 1929; Montero Agüera, 1971; Parenzan, 1970; Poppe and Goto, 1991, 1993; Van Aartsen et al., 1984, and literature cited therein).
Results
Chronostratigraphy
The Cal Maurici excavation corresponds to the basement of Depositional Sequence I (DS I) (Gámez, 2007; Gámez et al., 2009) and comprises a basal fluvial, beach and marsh facies and a young deltaic succession. DS I and the underlying DS II are characteristic of Gilbert-type fan deltas and are attributed to the deltaic plain and offshore facies lying above the Pliocene basement (Figure 3b and 3c) according to a 50-m-deep core (M:58 Cal Salom) drilled in the surrounding area (Gámez, 2007; Marquès, 1974).

Profiles and stratigraphy of Cal Maurici: (a) location of Cal Maurici and Delta del Llobregat cross-section based on Gámez (2007); (b) M:58 Cal Salom core based on Gámez (2007); (c) Delta del Llobregat cross-section based on Gámez (2007); (d) stratigraphic logs of several profiles from Cal Maurici; (e) summary profile of the site.
Figure 3 shows the stratigraphic framework of the site divided into five facies (Table 1) on the basis of the layers’ characteristics, that is, grain size (Figure 4), mineral composition, organic matter, colour (Table 2), stratigraphic position and elevation. This succession corresponds to the onshore sedimentary facies of the Llobregat delta, including seasonally brackish to freshwater, shallow marshes influenced by two mechanisms of sedimentation, the Mediterranean sea and the Llobregat river.

Particle size of several selected samples: (a) FP, M2 and M1 facies; (b) SR and RB facies.
Clay mineralogy, organic matter and colour of selected samples.
The lowermost unit of this sequence is a fluvial facies (RB), composed of grey or bluish grey, very coarse to medium sands, granules and well-rounded gravels with clay ball intercalations in all its profiles. This facies has a sharp erosional surface, convex-up geometry, charcoal fragments and few continental and marine fauna. The beach facies (SR), present only in the N part of the section (profile-I), overlies this lowermost facies and is composed of well-sorted sands. The separation between the two is clearly visible, thanks to the grain size composition, while SR presents fewer charcoal remains than the RB deposits. On the basis of its beach deposits and their geometry and its cross-bedding stratification, SR is presumed to be a sand barrier or dune.
Two similar swamp and marsh facies can be identified overlying RB and SR. The lower of the two (M1) presents yellow, fine sands and grey silt, while the upper facies (M2) presents grey laminated silt to sandy silt, rich in organic matter (peat intercalations) and brackish to freshwater fauna. Layers XIII and XVII of the M1 facies in profile-C comprise several bivalve characteristics and shell deposits typical of marine environments. A reduction in the presence of these fauna and an increase in that of freshwater elements as we move up the sequence are indicative of delta progradation. The sequence ends in a floodplain (FP) facies, composed of brown sandy silt with a high presence of moulds of roots stems of large plants and land and water snails.
The results of radiocarbon dating are shown in Table 3 and enable us to build a chronometric framework of the Cal Maurici section. Although the SR beach facies could not be dated owing to the absence of organic components, the radiocarbon dating of the transition between M2 and M1 indicates that this unit is older than 2σ interval of 6171–5773 cal. yr BP. The formation of the swamp and brackish facies (M1 and M2) began at this point in time, extending to 4843 cal. yr BP, when the FP facies began to be deposited, a process that extended until the 2σ interval of 3891–3712 cal. yr BP.
Radiocarbon dating.
Pollen
Pollen sampling was conducted on profile-B (Figure 5) and on facies M1, M2 and FP. At the base of M2 (#31 and #33), we find a decrease in AP as a consequence primarily of the decline in the presence of deciduous Quercus sp. type as well as that of other deciduous trees such as Corylus sp., Alnus sp., Betula sp. and Ulmus sp. In contrast, we observe an increase in shrubby taxa (Olea sp., Ericaceae and Buxus sp.) and an expansion of herbs such as Poaceae, Artemisia sp., Chenopodiaceae and Apiaceae. The occurrence of apophytes, including Plantago sp., Brassicaceae and Rumex sp. (Behre, 1981), should also be noted.
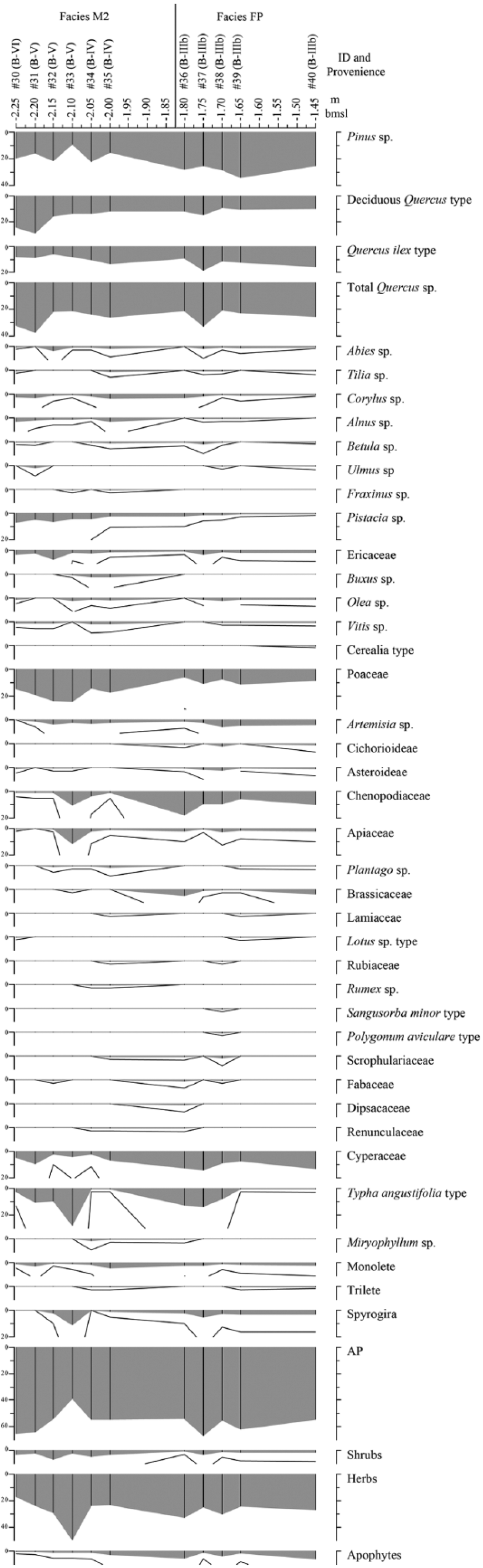
Pollen sequence from profile-B of facies M2 and FP and related to depth below sea level (m b.s.l.).
In the middle of the M2 facies (#34 and #35), AP values recover, due in the main to the increase in the occurrence of Quercus ilex type and of some deciduous trees, including Corylus sp., Alnus sp., Betula sp. and Tilia sp. This pollen change is coeval to a progressive decrease in the presence of shrubs (Pistacia sp. and Ericaceae) and herbs (Poaceae, Artemisia sp., Chenopodiaceae and Apiaceae). The occurrence of other herbs, including apophytes, such as Cichorioideae, Plantago sp. and Rubiaceae, increases. There is also a notable presence of Rumex sp., Scrophulariaceae, Fabaceae, Dipsacaceae and Ranunculaceae.
At the base of the FP facies (#36 to #39), a brief AP recovery occurs, mainly due to the expansion of Pinus sp. and Quercus ilex type, while Pistacia sp. and Buxus sp. decrease. At this juncture there is an expansion of Chenopodiaceae coeval to the decrease in Poaceae. Later, the upper samples of the sequence (#39 and #40) reveal a new decrease in the presence of Quercus ilex type and deciduous Quercus sp. type while Pinus sp. values increase. This upper part of the pollen record is also characterized by the decrease in other deciduous trees such Corylus sp., Alnus sp. and Betula sp. The decrease in Chenopodiaceae and the increase in Artemisia sp. are coeval with the increase and higher diversity of apophyte pollen taxa, such as Cichorioideae, Plantago sp., Brassicaceae, Lotus sp. type, Sanguisorba minor type and Polygonum aviculare type. The occurrence of Cerealia-type pollen is recorded in the upper sample.
Phytoliths and diatoms
Phytolith samples were taken from profiles B and C of the different facies (Table 4 and Figure 3). None of the samples analysed presented phytoliths in abundance, and in some samples none were identified (Table 4). Thus, our findings should be interpreted with caution. The small number of phytoliths recovered occurs independently of the type of sediment and facies. Associated with the phytoliths, diatoms were recovered in most profile-C samples. These were especially abundant in facies M1 (#52 and #55). Sponge spicules were noted in facies M2 and M1 (#49, #51 and #52).
List of phytolith samples.
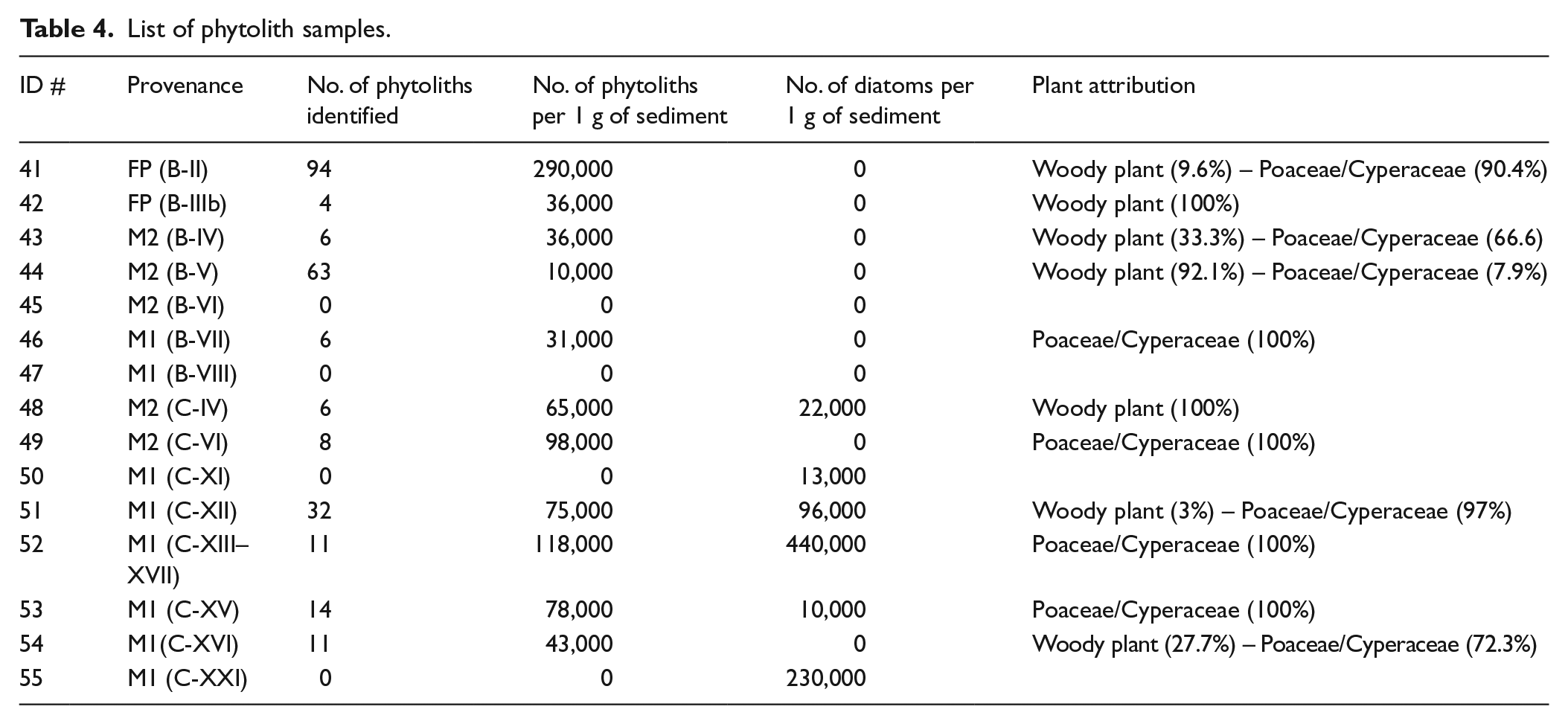
Despite the small number of phytoliths, the morphological types identified indicate a variation in plant composition depending on profiles and facies. Profile-B presents a mixed vegetation with Poaceae/Cyperaceae plants and woody/shrubby vegetation in most of the samples. Among these, the top of the FP facies (#41) presents the greatest phytolith variability. Most of the morphotypes correspond to elongated phytoliths with no decorated margins. These morphotypes have a low diagnostic value since they can be found both in Poaceae and Cyperaceae plants. Yet, the absence of any short cells, which are characteristic of grass, and the presence of cone-shaped phytoliths, which are characteristic of sedges, suggest that these elongates probably correspond to sedges rather than to grasses. Samples from profile-C differ from those in profile-B in that they present an almost exclusive presence of phytoliths formed by Poaceae/Cyperaceae plants, that is, elongated phytoliths with no decorated margins, associated with diatoms (Figure 6f) and sponge spicules (Figure 6e). In the case of the Poaceae family, the short cells identified are mainly of the rondel type, characteristic, together with trapezoids, of the pooideae C3 subfamily. One sample from the M1 facies (#55) presents, in addition to the rondel type, short cell bilobates, common both in C4 panicoid grasses and in reeds such as Arundo donax.
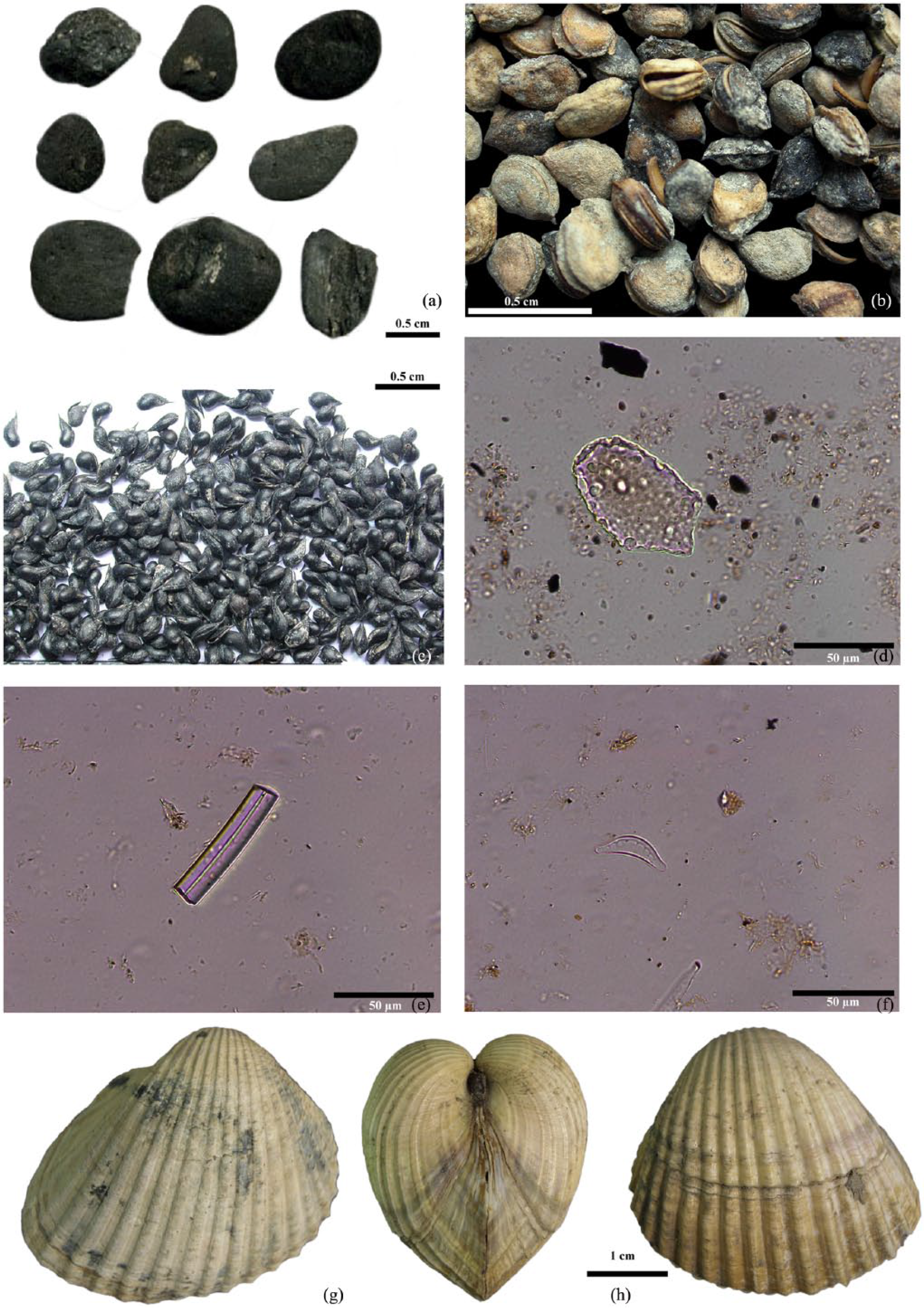
Cal Maurici palaeobotanical and malacological remains: (a) rounded charcoal remains from RB facies (#59); (b) Potamogeton sp. from M2 facies (#65); (c) Ruppia maritima from M1 facies (#75); (d) bulliform cell from RB facies (#41); (e) sponge cell from RB facies (#52); (f) diatom from RB facies (#55) characteristic of freshwater conditions; (g) Cerastoderma edule from M1 facies (#96 and #100); (h) Cerastoderma glaucum from M1 facies (#96 and #100).
Diatoms were, in general, highly fragmented and partially dissolved and did not permit a more detailed interpretation; nevertheless, some of the morphotypes identified are indicative of benthic and brackish environments and are commonly found in marine coasts and marshes. Few of the other morphotypes are indicative of freshwater conditions without further specification (Figure 6f) (Tomas et al., 1996).
Charcoal
The charcoal study focused primarily on profiles I and B. From these, the RB facies (#59) yielded most of the charcoal remains, whereas the rest of the samples presented very few remains (Table 5). The RB facies shows a high level of taxonomic diversity with 24 different taxa. Quercus sp. deciduous presents the highest values, representing 45% of the total. Corylus avellana, Taxus baccata and Buxus sempervirens present lower values, ranging between 4% and 9%, while the rest have a testimonial presence of less than 3% of the total of the fragments analysed (Table 5). Of the remains, 10% present a roundish shape, which could be a mechanical alteration resulting from their having been swept along in the river.
Results of the charcoal analysis.
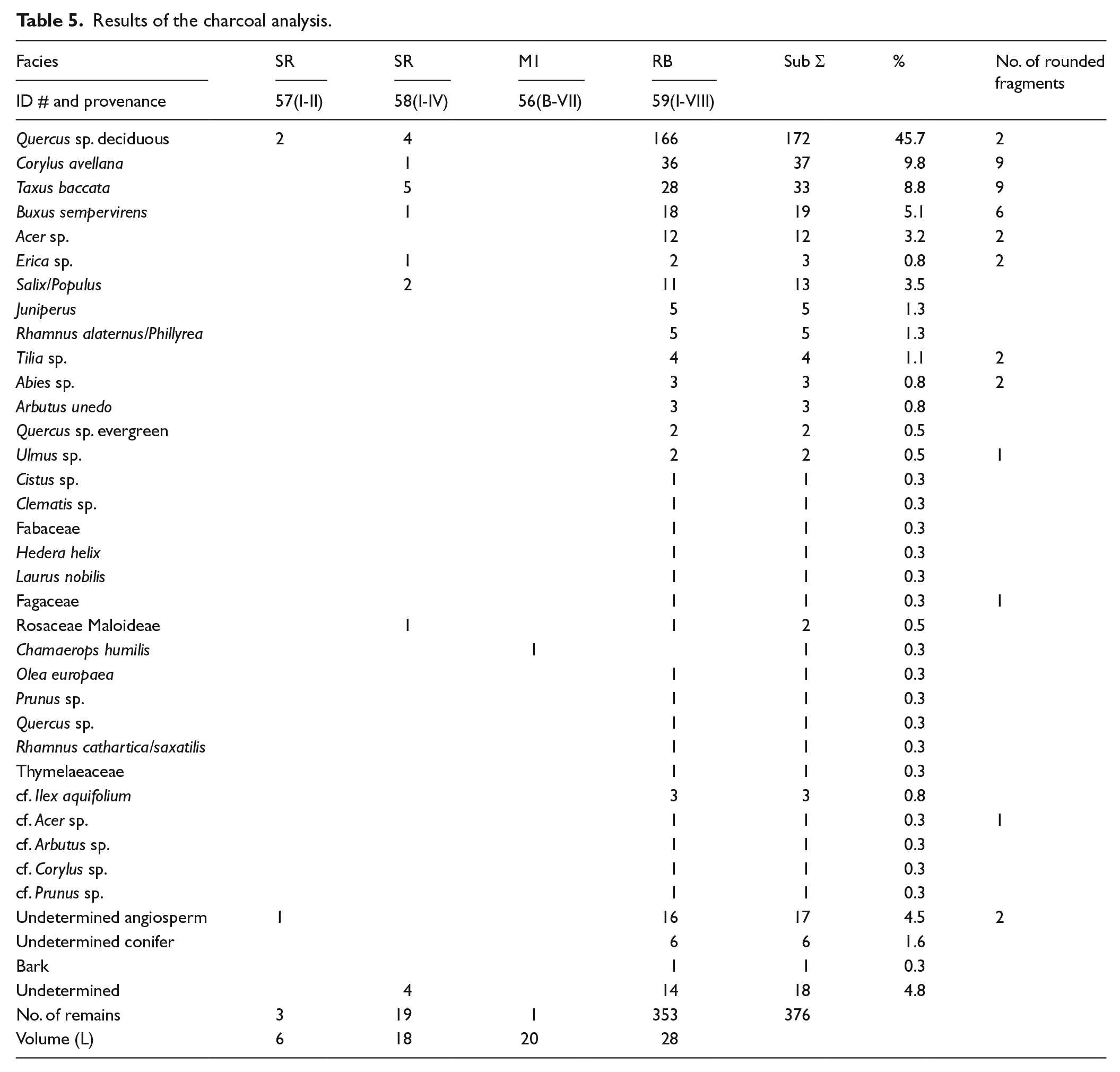
The assemblage presents species characteristics of different environments, representing a range of forest communities. Most of the taxa, including Quercus sp. deciduous, Acer sp., Tilia sp., Corylus avellana, Buxus sempervirens, Rhamnus cathartica/saxatilis, Rosaceae/Maloideae, Prunus sp. and Taxus baccata, correspond to woody vegetation growing in the deciduous forest communities of humid environments. They would have grown under such conditions from the pre-littoral mountain range to the Pyrenees. However, among these taxa, Tilia sp. and Corylus avellana are mainly present at higher altitudes of between 600 and 1000 m a.m.s.l. in the Pyrenees (Folch, 1986). Similarly, Abies sp. is located at higher altitudes of between 1200 and 1700 m a.m.s.l., corresponding also to humid environments (Folch, 1986). Fabaceae, Arbutus unedo, Erica sp. and Quercus sp. evergreen grow in evergreen forests or secondary plant communities that might have been distributed from inland areas along the coast to the pre-littoral mountain range. Olea europaea and Chamaerops humilis, the latter identified in the M1 facies (#56), are littoral species. Finally, Ulmus sp. and Salix sp./Populus sp. correspond to tree species characteristic of riverside forests.
According to the Holocene data from the anthracological records of the NE of the Iberian Peninsula, deciduous taxa are best represented in the pre-littoral mountain range during the Atlantic period (early Neolithic). Data from the sites along the coast indicate the presence of deciduous oaks during this period (Fontanals et al., 2008), suggesting the spread of deciduous forests to the coastline. From the sub-boreal onwards (mid-Neolithic), deciduous taxa mixed within a dominant evergreen forest at the coastal sites and are only well distributed at higher altitudes or in northern areas of NE Iberia (Allué et al., 2009; Piqué, 2005; Ros, 1996). Tilia sp. and Abies sp. are only recorded in the pre-Pyrenees and the Pyrenees (Allué et al., 2009). Corylus avellana has a wider distribution, but still corresponds only to north-eastern areas (Allué et al., 2009; Piqué, 2005; Ros, 1996).
The high taxonomic and ecological variability presented by the assemblage and the roundness of most of the charcoal fragments from the RB facies suggest that these remains could have been transported some distance before being deposited (Figure 6a). This facies, in accordance with the sedimentology, corresponds to a fluvial deposit. The taxa with the most rounded fragments in the RB facies are found at higher altitudes in the NE of Iberian Peninsula, at some distance from the coast, and would have been transported further along the course of the Llobregat river. Bearing in mind that the Llobregat river has its source at 1300 m a.s.l. and its course runs through territories that stretch from the Pyrenees to the coast, the presence of such a diversity of taxa and vegetation formations may be the result of the dragging of charcoal fragments from points along the river course before being finally deposited in the riverbed layers.
Seeds and fruits
Seeds from profiles B and C and the M1 and M2 facies are mainly composed of wild species (88%), 11% are indeterminate and only 0.1% correspond to cultivated plants (Table 6). The assemblage is dominated by aquatic plants representing 97.7% of the sample, while weeds and ruderals represent 2.2% and cultivated plants are only represented by barley (Hordeum vulgare cf. nudum) (0.1%). Among the most frequently occurring taxa, the highest mean values were presented by Potamogeton sp. (Figure 6b), representing 43.5% of all samples, 52.2% of identified remains and 58.5% of the remains identified in M1. In this last unit, it was followed by Ruppia cf. maritima (37.9%), while all other aquatic plants were less frequent, for example, Nymphea cf. alba (0.6%), Claudium mariscus (0.4%) and Najas cf. marina (0.2%), with weeds and ruderals being even less significant. The distribution of aquatic plants is indicative of depositional environments; the top of the M1 sequence (#66 and #70) is dominated by the seeds and leaves of Potamogeton sp. (98.3%), a plant that grows in slow-running, fresh and brackish waters of marshes and riversides; the base is composed mainly of Ruppia cf. maritima (89.0%), a plant that requires brackish conditions. Furthermore, in Cala B, Potamogeton sp. represents 70.3% of the samples while Ruppia cf. maritima is absent, while in Cala C, Ruppia cf. maritima (Figure 6c) represents 61.9% of the sample signalling more brackish conditions.
Results of the seeds analysis.

Charred.
Evidence of human activity is recorded in the presence of two charred cereal grains of Hordeum vulgare cf. nudum and Hordeum/Triticum from the M1 facies (#75) dated to 5038–4843 cal. yr BP in the late Neolithic period. The ruderal plant, Galium sp., could also be considered as part of this group as it is common in cereal fields; however, the current ecology must differ from that of the past, especially when considering natural depositions, such as Cal Maurici, and the early phase of agricultural development. Other remains, such as Pistacia lentiscus and Rubus sp., which were gathered as food by prehistoric communities, could also be related to human activities.
The presence of seed remains in facies M2 and FP is markedly smaller. The vegetation in M2 is composed mainly of Claudium mariscus (50.6%), Potamogeton sp. (22.7%) and Scirpus sp. (19.9%), all wetland plants that tolerate salinity. Conversely, no plants adapted to brackish conditions (i.e. Ruppia cf. maritime) are recorded. The presence of another aquatic plant, Nymphea cf. alba (4.0%), is also noted. In the FP facies, only woody plants and leaves have been documented.
Malacology
All the faunal remains correspond to malacological associations, except a carapace of Emys orbicularis from profile-D. The associations come from profiles B and D and present a similar composition, comprising some 17 species (microgastropods and bivalves). The taxonomic list of the fauna identified is shown in Table 7. Among the macro-molluscs, one freshwater gastropod (Radix peregra) in the M1 and M2 facies and 7, very common, marine bivalve species (Glycymeris insubrica, Ostrea edulis, Spondylus gaederopus, Cerastoderma edule, C. glaucum, Mactra stultorum and Scrobicularia plana) in the M1 facies were identified. Bivalves were only documented in the M1 facies (#96). Micro-molluscs are represented by a diversity of gastropods, the most significant being Skeneopsis planorbis, Obtusella intersecta, Truncatella subcylindrica, Peringia ulvae, Pseudamnicola confusa, Physa fontinalis, Ancylus fluviatilis, Bithynia tentaculata and Helicella sp. All species inhabit fresh and/or brackish water with the exception of Truncatella subcylindrica, which is a semi-terrestrial species, living close to the high water mark, not in direct contact with the sea, under stones, wood or driftwood (Barash and Zippora, 1992; Giannuzzi-Savelli et al., 1997; Poppe and Goto, 1991).
Results of the malacology analysis.
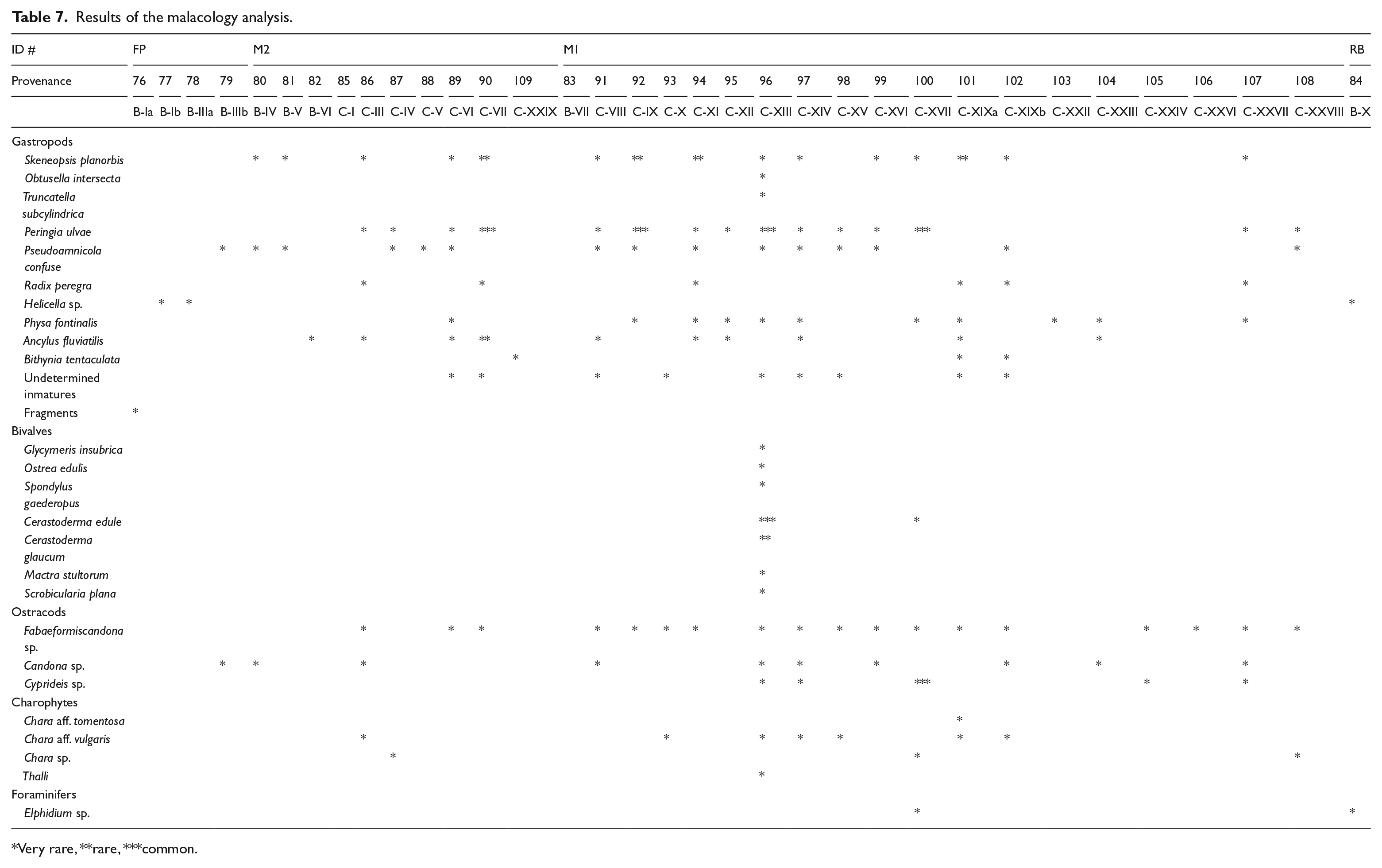
Very rare, **rare, ***common.
The highest level of malacological biodiversity corresponds to the samples from the M1 facies. The endofaunal bivalves, Cerastoderma edule and C. glaucum, dominate the assemblage in one of the samples (over 90% in terms of the number of individuals), indicating a high level of biodiversity but a low level of ecological diversity (Figure 6g). Both species usually inhabit sands and muddy sands within transitional environments and are adapted to salinity oscillations produced by freshwater inputs (Poppe and Goto, 1993). Their presence in the M1 and M2 facies corroborates the brackish/freshwater character of the environment. Epifaunal oysters and spondylus are scarce, being represented by big large valves (some 7–8 cm long), while findings of single right valves are common. These shells show signs of transportation and some bioerosion traces (Entobia, Caulostrepsis) are visible.
Peringia ulvae – also known as the mudsnail – is a very abundant hydrobiid microspecies at the site. It generally inhabits transitional environments (estuaries and salt marshes) and tolerates marine to brackish waters (Graham, 1988). Ancillus fluviatilis was collected only in the M1 and M2 facies. This is a delicate, rheophilic freshwater microlimpet that inhabits fast-running waters (Haas, 1929). The skeletons are unbroken, indicating the total absence of transport.
The endofaunal bivalves in the M1 facies are indicative of a very shallow environment within an estuary, with muddy bottoms and an abundance of vegetal remains. Some sporadic shells of the epifaunal bivalves (Ostrea edulis and Spondylus gaederopus) correspond to specimens reworked from neighbouring coastal areas. The shells of freshwater (Radix peregra) and semi-terrestrial (Truncatella subcylindrica) snails appear to have been carried by the river or to inhabit ponds close to the coast.
Site summary and discussion
Chronology
We have constructed a section by way of summary for the site based principally on profile-B (Figure 3e). The results show that the Cal Maurici deposits span a period from 6171 to 3712 cal. yr BP. This corresponds to layers dating to the mid-Holocene, that is, to the late middle Neolithic period and Bronze Age. Facies RB and SR were deposited before the late middle Neolithic and could correspond to the earlier Neolithic, facies M1 and M2 were deposited during the interval of 6171–4960 cal. yr BP, that is, from the late middle Neolithic to late Neolithic, while the age of the lower FP facies is statically the same as that of the M2 facies, spanning between 5026 and 3712 cal. yr BP, that is, late Neolithic to the Bronze Age.
Palaeoenvironmental reconstruction
Ascertaining connections between past climate and vegetation records from sediment traps can be complex, and requires careful consideration of sedimentary formation and post-depositional processes. At Cal Maurici, the apparent physical occurrence of several types of vegetation could be misleading, given potential differences in their preservation, transport and dispersion in water flow. For example, pollen may have been constrained by differences in pollen production and wind transport, some charcoal remains present a roundish shape in the RB facies signalling river transport and phytoliths are very scarce presumably because of dissolution processes. Despite these difficulties, the study conducted at Cal Maurici is the first multi-proxy analysis aimed at reconstructing the palaeoecological conditions of the Llobregat wetlands, in particular, and of the western Mediterranean facade, in general.
Seed and fruit remains provide the best insights into the vegetation record and appear to signal the presence of transitional environments from shallow brackish to freshwater marshes, especially in profile-C. Aquatic plants are homogeneous and composed primarily of Potamogeton sp., Cladium mariscus, Scirpus cf. maritimus and Nymphea cf. alba while Ruppia cf. maritime could have constituted mono-specific beds. Records of other aquatic plants are fewer in number, for example, Najas cf. marina, a species that generally thrives in calm environments on soft bottoms of freshwater or brackish environments. The identification of sedge phytoliths in profile-C is consistent with the above results. Associated with these aquatic plants, the occurrence of diatoms characteristic of brackish environments and marshes supports the interpretation of this area as a calm environment. Moreover, a gradient is found in the synchronic distribution, where the presence of plants adapted to brackish environments is more evident in a seaward direction. For example Ruppia cf. maritima is only recorded in profile-C (layers XIII and XVII) and limited onshore, while Potamogeton sp. is widely recorded in facies M1 in profile-B and more limitedly in profile-C.
The diachronic distribution of plants along the stratigraphic profile of Cal Maurici signals brackish environments composed primarily of Ruppia cf. maritima from the base of M1 (6171–5773 cal. yr BP) to the fresh and brackish conditions dominated by Potamogeton sp. at the top of M1 (5026–4839 cal. yr BP). The malacological analysis seems to confirm this, as marine bivalves are only recorded in the same profile and layers (C-XIII and XVII, i.e. M1 facies), while others signal the alteration of salt and brackish or freshwater conditions. The pollen analysis also supports this interpretation, as the peaty layers of facies M2 and FP are mainly rich in hygrophilous plants, including Typha angustifolia type and Cyperaceae, with the presence of Spirogyra sp. algae (Van Geel, 2001).
Pollen samples from facies M2 (before 4960–4825 cal. yr BP) point to a forested landscape dominated by deciduous trees, primarily oak woodlands. Along the riverbanks and in areas in shadow, other deciduous trees such as Corylus sp., Alnus sp. and Betula sp. might also have had a presence. The prevalence of deciduous oak woodlands in the Cal Maurici sequence is coherent with the charcoal evidence and corroborates former pollen data, which stress the dominance of such forests (Riera, 1995; Riera and Esteban, 1994). However, several differences occur in the area. Thus, in southern sequences, the presence of evergreen and thermophilous wooded taxa is higher than that in northern sequences during the Holocene, as shown by high percentages of Pistacia sp. at Cal Maurici (Riera, 1994, 1995). Despite the low number of phytoliths recovered, which impedes any further interpretation, the few morphotypes identified are consistent with the other results signalling the presence of mixed vegetation formed by Poaceae/Cyperaceae plants and varying degrees of wood/shrubby vegetation all along the sequence.
At some point during the formation of the FP facies (later than 4960–4825 cal. yr BP), the above forest configuration changed with a decline in deciduous trees and mastic communities and the expansion of evergreen oaks (Quercus ilex type), pinewoods, wild olive trees (Olea sp.) and box (Buxus sp.). Similar changes in wooded vegetation have been recorded across the Barcelona plain in earlier pollen studies, but with uncertain chronologies (Riera, 1994; Riera and Esteban, 1994). In the southern sector of the plain, the expansion of both box and pinewoods has also been recorded, albeit with an uncertain chronology, although later than 6192–5589 cal. yr BP (5100 ± 140 BP) (Riera, 1995, 2009). However, the greater presence of pines in littoral areas suggests that pinewoods were able to develop in littoral and sandy soils. The presence of pines in anthracological assemblages is also recorded from this period towards the interior (pre-Littoral mountain range) of this territory (Allué et al., 2009). Other anthracological records from coastal sites in NE of Iberian Peninsula suggest a gradient between the south, with thermophilous formations characterized by a predominance of Pistacia lentiscus and Olea europaea (Piqué, 2005; Ros, 1996), and the north, with a marked predominance of oaks. During the early Neolithic, deciduous oaks dominated the landscape, but from the middle Neolithic, evergreen oaks and scrubland (Ericaceae, Buxus sempervirens) formations emerged. At Cal Maurici, the presence of rounded deciduous charcoal remains does not allow an accurate diagnosis to be made as they might have been transported here from some distance, or, on the basis of the pollen analysis, they could be indicative of a vegetation type in the more immediate vicinity.
Human impact
Our results indicate that the vegetation on the Llobregat floodplain and in the surrounding forested areas changed along the sequence and so we can speculate as to human activity played any role in these changes. Evidence of human impact on the landscape occurs at several points in the sequence signalling a significant change in the vegetation. However, the palaeoenvironmental data available for the NE of the Iberian Peninsula suggest that climatic variations may also have been relevant in this period (Allué et al., 2009; Carrión et al., 2010; Revelles et al., 2014).
Two charred cereal grains of Hordeum vulgare cf. nudum and Hordeum/Triticum have been documented in facies M1 (#75), that is, middle–late Neolithic period. These remains are direct vestiges of human agriculture; however, the absence of archaeological evidence in the Cal Maurici wetland, as a consequence of rapid sedimentation dynamics area, does not allow us to speculate any further on the agricultural activity conducted in this area and as to whether Neolithic crops were well established in the Llobregat wetlands. However, the middle Neolithic mines of Can Tintorer (Borrell et al., 2015) are located at a distance of ~5 km from Cal Maurici and could serve to reinforce the idea of a well-developed settlement in this area. Likewise, the presence of the Cova de Can Sadurní, located at a distance of 12 km, is further evidence in favour of these hypotheses, given that thousands of cereal grains have been recovered there (Antolín and Buxó, 2011).
Although other plants (Galium sp.) from the M1 facies (same sample: #75) may be ruderal, and hence serve as indicators of crop practices when documented at archaeological sites, they are nevertheless not a clear signal in natural sediment traps. Other fruits identified, including Pistacia lentiscus, Rubus sp. and Vitis sp., were commonly gathered by prehistoric farmers as a plant product in the NE of the Iberian Peninsula (Antolín and Jacomet, 2015). At Cal Maurici, the Rubus sp. seed identified in the same facies (M1) and sample (#75) is charred. As such, it might be attributed to a chance event or result from the burning of cereal stubble, given that at other middle Neolithic sites such seeds are directly related with human consumption (Antolín and Jacomet, 2015). The Vitis seed could not be assigned to either a vine grape (Vitis vinifera ssp. vinifera) or a wild grape (Vitis vinifera ssp. sylvestris) since only one example was recovered and the results of the biometric analysis were not statistically significant. Wild grape has been broadly documented in the NE of the Iberian Peninsula throughout the Neolithic and is recorded at the nearby Can Tintorer mines and in several Bronze Age sites (Buxó, 1997). However, it was not until the Iron Age that this fruit became more prevalent in the archaeological record of the NE of the Iberian Peninsula (Buxó, 1997), while the vine grape is not documented until VII ac (Buxó, 2008; López Reyes, 2004; López Reyes et al., 2011).
Pollen analysis confirms the interaction between early farmers and the landscape. Vegetation changes occurred at the base of the M2 facies, shortly before the radiocarbon date 4960–4825 cal. yr BP, indicating human impact on landscape. This is evidenced by the decrease in AP values, the increase in shrubby communities (mainly Ericaceae) and the occurrence of apophytes such as Plantago sp., Brassicaceae and Rumex sp. (Behre, 1981; Riera, 1995). Similarly, evidence of human impact through the use of fire has also been observed in pollen sequences of the Llobregat deltaic plain (Riera, 2009), probably corresponding to the same phase of human activity reported at Cal Maurici. Charcoal records from the inner and northern areas also provide evidence of human impact with differences emerging according to biogeographical areas (Allué et al., 2009; Revelles et al., 2014). Activities related to forest clearance for crops or to forest management for the use of wood as fuel or fodder would probably have affected the arboreal cover. It should also be noted that significant aridity, due to general climatic variations, might have favoured the transformation of forests (Carrión et al., 2010; Fletcher et al., 2007).
Conclusion
The ~4-m-long core of the Cal Maurici sequence has been shown to be especially valuable for the palaeoenvironmental reconstruction of the mid-Holocene in the north-east of the Iberian Peninsula. The multi-proxy record obtained by combining data from different sources (namely, phytoliths, integrating uncarbonized and carbonized seeds and fruits, pollen, charcoal and malacology) provides a more accurate reconstruction of the biotic community in this coastal ecosystem than that achieved in previous studies (Riera, 1994, 1995, 2009). Furthermore, it has traditionally been difficult to reconstruct Holocene terrestrial archives in the Mediterranean area because of the absence of long stratigraphic sequences and so studies have often been based solely on the palynological analysis of lake deposits, including for example those undertaken for the Iberian Peninsula (Jalut et al., 2000; Reed et al., 2001), the Rhone delta (Muller et al., 2008) and Italy (Tinner et al., 2009). Here, we have demonstrated the advantages of undertaking multi-proxy analyses (macro- and micro-fossil studies) at Cal Maurici, the evidence from which is all in close agreement.
Our sampling programme highlights the ecological richness of these wetlands and appears to confirm the importance of the role played by coastal and deltaic environments during the expansion and settlement of pioneer farming communities, that is, the geographical distribution of the early Neolithic sites along the coast from Liguria to Valencia (Guilaine and Manen, 2007) emphasizes the importance of rich marine environments and maritime movement. The presence of domesticated plants (cereals) is indicative of the interactions between early farming populations and coastal environments during the earliest phase of agricultural expansion in the south-eastern corner of the Old World, and these anthropic markers are perhaps related to the presence of crop cultivation in these wetlands. Additionally, the large number of malacological species found at Cal Maurici points to the littoral richness for prehistoric human procurement, as has been evidenced at nearby archaeological sites. To date, evidence of human activity provided by seeds has only been recorded at archaeological sites (in storage pits, settlements and caves) and has been absent from natural traps. Here, however, our multi-proxy study serves to reinforce earlier palynological data and points to the direct human impact of early agriculture on coastal ecosystems. The palaeoenvironmental programme reported here identifies Cal Maurici as one of the few sites along the Mediterranean coast of the Iberian Peninsula where a human presence in brackish marshes dates back to the earliest phases of the Neolithic.
In short, Cal Maurici’s palaeoenvironmental record is in close agreement with the changes recorded in other areas of the Mediterranean, where the climatic variations that seem to have occurred during the mid-Holocene coincided with human activities that affected the vegetation and the landscape, placing both under increasing pressure (Branch and Marini, 2014; Branch et al., 2014; Mercuri et al., 2011; Revelles et al., 2014). Moreover, the results of this multi-proxy analysis are an important source of information for gaining a better understanding of coastal/wetland archaeology and, hence, of the close relationship between the last foraging and the first farming communities, on the one hand, and coastal areas, on the other, as well as of the impact of Neolithic practices on landscapes of this particular type.
Footnotes
Funding
This paper is the outcome of the archaeological research project El Plistocè Superior a la costa central catalana: paleoambients i ocupació dels neandertals sponsored by Servei d’Arqueologia i Paleontologia – Generalitat de Catalunya (2014/100639) and Grup de Recerca del Quaternari. Fieldworks were sponsored by ADIF-Alta Velocidad (Ministerio de Fomento). Sampling and laboratory analyses were funded by Catalan (2014SGR-00108) and Spanish (HAR2014-55131, HAR2013-42054-P and CGL2013-48441-P) MICINN projects. J Daura and M Sanz have been supported by a postdoctoral grant (SFRH/BPD/100507/2014 and SFRH/BPD/100828/2014) from the Portuguese Fundação para a Ciência e a Tecnologia using funding from the FSE/POPH and Ll. Picornell-Gelabert by the Conselleria d’Educació of the Government of the Balearic Islands-FSE.