
Abstract
Mixed millet and rice agriculture is a unique agricultural style of China, and is distributed in a broad band between the Yangtze and Yellow River basin. However, the development of this style during the Neolithic-Bronze Age has not been comprehensively clarified, owing to limited archaeobotanical data and imprecise chronology for most of the regions. Here, the Central Plain, a location where mixed agriculture may have first appeared, was selected as the area for research. Phytolith and macrofossil analyses from the Zhuzhai site, together with the accelerator mass spectrometer (AMS) radiocarbon dating of samples, reveal information about the temporal changes of mixed agriculture as well as the domestication and cultivation of crops in this region. The results indicate that mixed millet and rice agriculture formed in the Central Plain about 8000 years ago. Common millet was the principal crop in the Peiligang and Yangshao periods, with the domestication process beginning in the Peiligang period and continuing up to the Shang period, at which time it was replaced by foxtail millet. Foxtail millet may have gone through a significant degree of domestication by ca. 6000 cal. BC, but its domestication process was still unclear. Rice had appeared since the Peiligang period, but its proportions were always low. Rice assigned to the Peiligang and Yangshao Cultures was the domesticated japonica, and its cultivation was always performed in dry field systems through the Neolithic-Bronze time. Within the subsistence economy, mixed agriculture was a minor component during the Peiligang period, but has been dominant since the Yangshao period.
Introduction
It is generally accepted that China is an independent center where the domestication of millets and rice took place (Larson et al., 2014). This domestication is regarded as a protracted process that eventually led to the appearance of millet and rice agriculture in the Yellow River and Yangtze River valleys, respectively (Fuller et al., 2014; Gross and Zhao, 2014; Zhao, 2011a). The development and interplay of these two agricultural systems during the Neolithic-Bronze Age not only facilitated prehistoric cultural evolution and demographic expansion in China (Wang et al., 2014) but also was fundamental to the formation of ancient Chinese civilization (Liu and Chen, 2012). Hence, in view of their profound impacts on human societies, the timing, location, processes, dispersals, and spatial-temporal distribution patterns of millet and rice agriculture have been subjected to intensive research in China within the past two decades (Cohen, 2011; Jones and Liu, 2009; Lu, 1999; Qin, 2012; Zhao, 2014).
At present, archaeobotanical evidence has certainly suggested that the origin of Chinese agriculture can be traced back to the early Holocene, with the earliest domesticated millets and rice dating between 8000 and 6000 cal. BC (Deng et al., 2015; Lu et al., 2009a; Yang et al., 2012; Zheng et al., 2016; Zuo et al., 2017). Meanwhile, a north-south pattern for the development and distribution of millet and rice farming was confirmed (Crawford, 2006; Lu, 2006). However, this division was not distinct, as there was a large area of millet and rice mixed agriculture that existed between the Yellow and Yangtze Rivers. This was a result of the bidirectional spread between both kinds of agricultural systems (Qin, 2012; Wang and Xu, 2003). Furthermore, regional differences of mixed millet and rice farming have been recognized, with the relative importance of millets versus rice varying among the Hanjiang River valley (Deng et al., 2015; Weisskopf et al., 2015a), Huaihe River valley (Yang et al., 2016a), Weihe River valley (Zhang et al., 2010b), Shandong Highland (Crawford et al., 2016; Jin et al., 2016), Chengdu Plain (d’Alpoim Guedes et al., 2013), and Central Plain (Lee et al., 2007). Even within the same region, the agricultural preferences on millets or rice were likely to make a temporal change throughout different cultural periods. Previous studies have provided some sequences of agricultural development in different mixed farming regions (e.g. Deng et al., 2015; Lee et al., 2007; Yang et al., 2016a); however, excellent plant-remains data with accurate chronology for most regions is still limited, particularly with respect to the early stages of the establishment of mixed millet and rice agriculture (Liu et al., 2008). Thus, the diachronic development of mixed agriculture in China during prehistoric times cannot be comprehensively clarified at this point. More archaeobotanical evidence and radiocarbon dates from Neolithic-Bronze Age sites in the mixed farming regions, especially those key areas previously mentioned in this section, are needed to resolve this problem.
One of the ideal regions for addressing the above problem by means of archaeobotanical research is the Central Plain region. Given that early plant remains of common millet (Panicum miliaceum), foxtail millet (Setaria italica), and rice (Oryza sp.) were recovered from the Central Plain (Lee and Bestel, 2007; Pang and Gao, 2006; Wu, 1994; Zhang, 2011a; Zhang et al., 2012; Zhao, 2011b), this region is acknowledged as a home to the millet agriculture in North China and one of the major regions for the northward spread of rice agriculture from South China (Qin, 2012; Wang, 2013). The meeting of millet and rice in the Central Plain is not only significant to the origin and spread of agriculture, but it also coincided with the population migration and technical interchange between North and South China (Zhang and Hung, 2013; Zhang, 2011b). This facilitated great socioeconomic development, as well as the emergence of the City State, making the Central Plain a cradle of Chinese civilization (Liu, 2007; Yan, 1987).
Evidence of ancient agriculture in the Central Plain is mainly based on plant macroremains. Numerous assemblages of charred plant remains have been collected from the Yiluo valley (Lee et al., 2007), upper Ying valley (Fuller et al., 2007b; Zhang et al., 2010a), Luoyang basin (Zhang et al., 2014), and some single sites (Qin, 2009; Zhao, 2007, 2011b) of the Central Plain, which span from the Neolithic to Shang-Zhou Dynasties. Quantitative analysis of these plant macrofossils indicated that millets were consistently the principal crops, and while rice was introduced by the late Yangshao period (ca. 3500–3000 cal. BC), it was not always a stable component of agricultural economy (Lee et al., 2007; Qin, 2012; Zhao, 2011b). Nonetheless, data on plant macroremains for this region are limited to the early Neolithic, that is, Peiligang period (ca. 6500–5000 cal. BC), because of poor preservation conditions. Only few charred millet grains have been discovered from Peiligang Culture sites, and rice is absent (Lee et al., 2007; Yang et al., 2015; Zhang et al., 2014). Recently, studies of plant microremains have filled this gap. Residue analyses of stone tools and potteries from eight Peiligang Culture sites have found abundant millet starch grains (Liu et al., 2013; Yang et al., 2015; Zhang, 2011a), especially at the Tanghu site, where both millets and rice starch grains and phytoliths were excavated (Yang et al., 2015; Zhang et al., 2012). This implies that the Central Plain might be the place in which mixed millet-rice farming first appeared around 8000 years ago, parallel to that of the western Shandong area (Crawford et al., 2006, 2016; Jin et al., 2014). Since the time of origin is basically confirmed, the present study raises critical questions about the temporal changes of mixed millet and rice agriculture in Neolithic-Bronze Age Central Plain: How did the relative proportion of millets versus rice in crop assemblages and foxtail millet versus common millet in dry farming change over time? What was the specific domesticated status of millets in different periods and how did the process of millet domestication occur? How did the nature (wild or domesticated/japonica or indica), as well as the arable systems (wetland or dryland) of rice change through time?
In order to answer these questions, the Neolithic-Bronze Age site of Zhuzhai was investigated. The Zhuzhai site (113°30′19″E, 34°49′31″N) is located in Zhengzhou City, Henan Province, bordered by the Yellow River to the north and the Songshan Mountains to the south (Figure 1). The site is named after the nearby Zhuzhai village situated 0.5 km to the west, and occupies the tableland at the confluence of the Xushui River and its tributary with an altitude of about 105 m a.s.l. The site has various phases of occupations, with the earliest one being the Peiligang Culture, followed by another seven cultural periods: Yangshao Culture, Shang Dynasty, Western Zhou Dynasty, Warring States, Han Dynasty, Tang Dynasty, and Song Dynasty. In 2011 and 2012, the site was excavated by Zhengzhou Municipal Institute of Archaeology, and a total of 5000 m2 in settlement occupations were uncovered (Liu and Bao, 2012). During the excavations, 340 archaeological contexts were found, including ash pits, house foundations, burials, pottery kilns, ditches, and a large number of artifacts such as stone tools, bone implements, and ceramics. These remains mainly belong to the Peiligang Culture, Yangshao Culture, and Shang Dynasty, with few remains originating from other periods (Bao, 2017; Liu and Bao, 2012). The Zhuzhai site is considered as an ordinary settlement without central function during all phases of occupations, and only the Yangshao remains have clear function zoning; the eastern part of excavated area includes living, production, and burial areas, while high-hierarchy buildings, including a big house foundation (30 m2) and a wide drainage ditch, occurs in the western part (Liu and Bao, 2012).
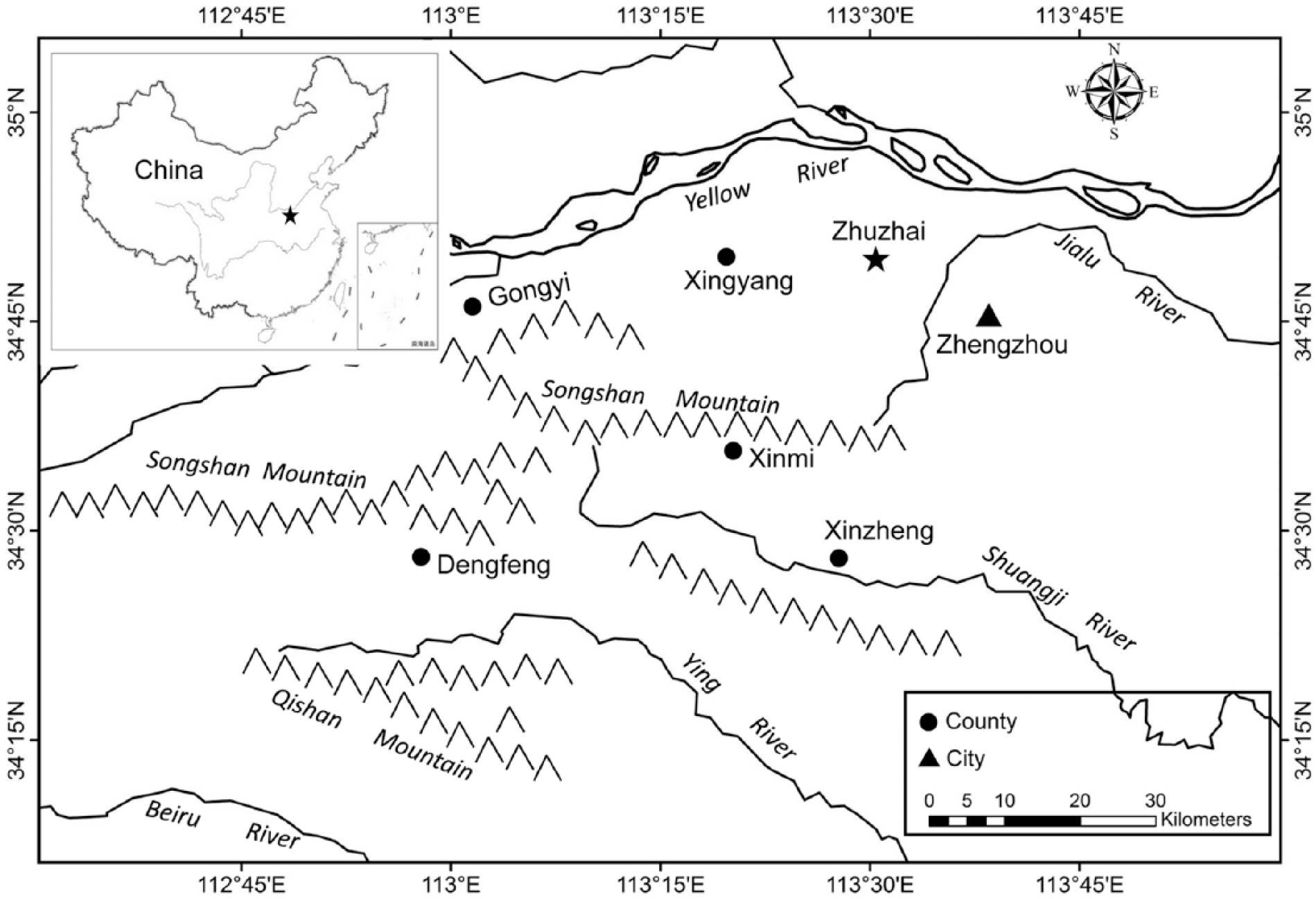
Location of the Zhuzhai site.
Based on phytolith and macrofossil analyses, together with AMS radiocarbon dating, the development process of mixed millet-rice agriculture in the Central Plain during the Neolithic and Bronze Age was determined. Moreover, statistical analysis of morphotypes and morphological parameters was conducted on archaeological phytolith assemblages to shed light on the nature of crop remains and whether rice was cultivated in a wet or dry field at the site. The study provides new data for clarifying the formation and progress of mixed millet-rice farming in China from a regional perspective, and has implications for understanding the cultivation, domestication, and spread of the two crops in prehistoric periods.
Materials and methods
A total of 40 soil samples for phytolith analysis were collected from the Zhuzhai site, including 12 samples from the Peiligang period (ca. 6500–5000 cal. BC), 21 samples from the Yangshao period (ca. 4500–3000 cal. BC), four samples from the Shang Dynasty period (ca. 1600–1000 cal. BC), and three samples from the Warring States period (deposits from a pottery fu caldron in M36; 475–221 cal. BC) (Table 1). Systematic flotation work was not applied to the Zhuzhai site because of the fact that it was a salvage excavation project and was only given a short amount of time for investigation. However, archaeologists excavating the site still collected 49 L of soil from six flotation samples for plant macroremain analysis. These samples were taken from six ash pits, in which four pits (H158, H208, H218, H226) were assigned to the Peiligang Culture and two pits (H202, H225) were assigned to the Yangshao Culture.
Summary of samples collected for phytolith analysis from the Zhuzhai site.

The method used for phytolith extraction from the soil was wet oxidation, as described by Wang and Lu (1993) and Piperno (2006). The procedure consisted of weighing out 2 g of soil and adding 30% hydrogen peroxide (H2O2) and 10% cold hydrochloric acid (HCl) to each sample to remove organic matter and carbonates. Meanwhile, a Lycopodium Spore tablet (27,637 grains/tablet) was added to detect the concentrations of phytoliths for each sample, while separating with zinc bromide (ZnBr2, density 2.35 g/cm3) heavy liquid. The sample was ultimately mounted on a slide with neutral resin and fixed with a cover glass. Phytolith identification, counting, and photographing were conducted using a Leica DM750 light microscope at 400× magnification. More than 400 phytoliths were counted in each sample. Phytolith identification and classification was aided by the use of published references (Lu et al., 2006; Wang and Lu, 1993). Millet phytoliths were identified by reference to Lu et al. (2009b), and the identification of double-peaked, bulliform, and parallel-bilobe rice phytoliths was performed with reference to Fujiwara (1993) and Lu et al. (1997). Phytolith abundance was quantified using frequency expressed as a percentage of all phytoliths counted.
In order to determine the extent to which the millets were domesticated, the morphological parameters of millet husk phytoliths from the site by period were measured, as described in the work of Lu et al. (2009b), Zhang et al. (2011), and Ge et al. (2016). Then, these data were compared with that of modern millets obtained from a previous study (Lu et al., 2009b) in order to determine the morphological variations of phytoliths in millet domestication. For the determination of the nature of rice phytoliths, the number of scale-like decorations of rice bulliform phytolith and the morphometric parameters of both double-peaked and bulliform phytoliths were determined based on previous works (Gu, 2009; Gu et al., 2013; Huan et al., 2015; Lu et al., 2002; Wang and Lu, 2012; Wu et al., 2014b; Zhao et al., 1998; Zheng et al., 2003). The measurement of phytolith morphological parameters and data procurement was performed on images using ImageJ software. Then, a sensitive/fixed phytolith morphotype model defined by Weisskopf et al. (2015b) was applied to sediment samples from cultural contexts (i.e. ash pits) to track degrees of wetness in rice arable systems. Because the samples for this analysis should contain a substantial concentration of rice (6900 grains/g ~ 600 × 104 grains/g) (Weisskopf et al., 2015b), the subsequent number of selected samples is 2, 10, and 3, assigned to the Peiligang Culture, Yangshao Culture, and Shang Dynasty, respectively.
For the extraction of charred plant remains, the six samples were floated by bucket with a 0.2 mm mesh screen at the site. After drying, the light fraction samples were sent to the Paleontology Lab of the Institute of Geology and Geophysics CAS for sorting. These samples were first sifted in sample sieves (2, 1, 0.7, 0.5 mm), and the <0.5 mm fractions were excluded because no charred seeds or fruits were found in these tiny remains. The remaining samples were sorted under a Leica L2 binocular stereomicroscope. Charred seeds, fruits, and plant remains were moved from charcoal and stored in plastic tubes with the context number. These charred plant remains were then sent to the State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany CAS, and identified by Mr Changjiang Liu with reference to modern plant specimens and published identification keys (Liu et al., 2008).
A total of eight charcoal samples were selected for radiocarbon dating with an AMS at Beta Analytic Radiocarbon Dating Laboratory. All 14C dates were reported in years before present (BP, meaning from 1950 CE) and based on the Libby half year of 5568 with a 1σ standard deviation. Then, the 14C dates were calibrated using the OxCal 4.2.3 program (Ramsey, 2009) and IntCal13 curve (Reimer et al., 2013) with ranges expressed both at 1σ (68.2%) and 2σ (95.4%) confidence level. All calibrated dates reported were referred to as ‘cal. BC’.
Results
Chronology
The eight AMS dates obtained from the Zhuzhai site are shown in Table 2. Except for two abnormal dates from H218 and H214, the dates are in line with the cultural ages. The available dates can be grouped into two sets. The first set contains four dates covering the time interval 6050–5730 cal. BC (95.4% range) which falls within the age of the Peiligang Culture. The second set covers the period from 3348 to 3090 cal. BC (95.4% range), which corresponds to the late Yangshao Culture. The chronology of the samples from the Shang Dynasty and Warring States period is based on the archaeological ages inferred from archaeological stratigraphy and unearthed objects because of the absence of available materials for radiocarbon dating.
Results of AMS 14C dates from the Zhuzhai site.

Abnormal dates are shown in italics. AMS: accelerator mass spectrometer.
Phytoliths
Phytoliths from the Zhuzhai site were mostly well preserved with distinct features for correct identification (Figure 2). The phytolith contents of all samples were abundant, with a mean concentration of phytoliths reaching approximately 695 × 104 grains/g. Finally, a total of 24,174 phytoliths were counted in 40 samples and assigned to 29 types according to the classification system of Wang and Lu (1993) and Lu et al. (2006). Crop phytoliths were found in 30 samples, including double-peaked, bulliform, and parallel-bilobe types for rice (Figure 2d–f) and millet husks (η-type from common millet: Figure 2a and b; Ω-type from foxtail millet: Figure 2c). Other main phytolith types are short saddle, long saddle, bilobate, cross, reed bulliform, smooth-elongate, sinuate-elongate, rondel, trapeziform sinuate, square, and so forth.
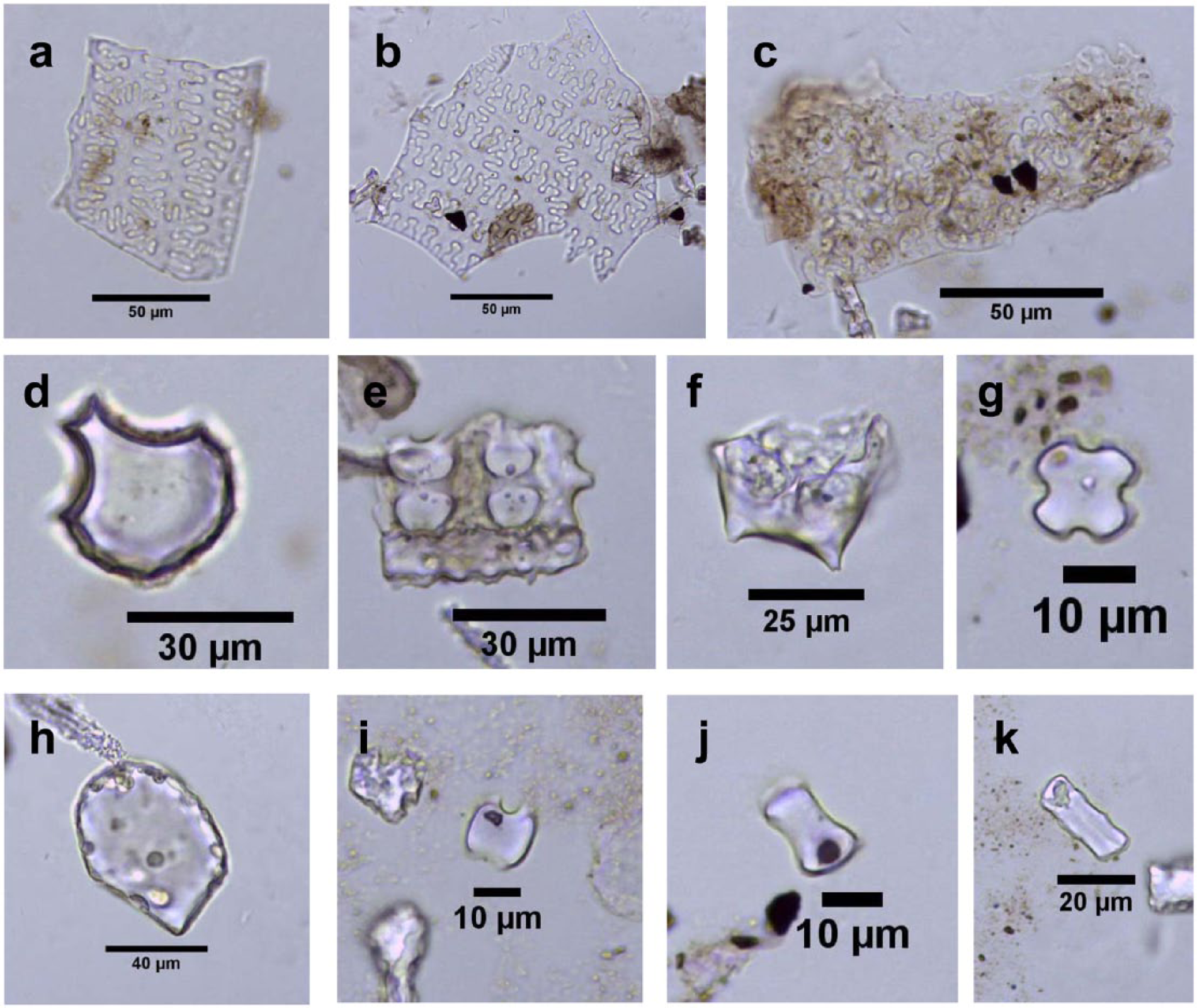
Main phytolith types from the Zhuzhai site. (a, b) η-type from husks of common millet, (c) Ω-type from husks of foxtail millet, (d) bulliform from rice leaf, (e) parallel-bilobe from rice leaf/stem, (f) double-peaked from rice husk, (g) cross, (h) scutiform-bulliform from reed, (i) short saddle, (j) long saddle, and (k) trapeziform sinuate.
Phytoliths from the Peiligang Culture period
A total of 5845 phytoliths were counted from 12 samples belonging to the Peiligang Culture, of which nine samples contained crop phytoliths (Figure 3). Present in these samples were 448 pieces of common millet husks, 32 rice phytoliths (including 29 double-peaks and three rice bulliforms), and only four pieces of foxtail millet husks. The highest percentage of common millet was 40%, while the highest percentages of rice and foxtail millet were 5% and 0.6%, respectively (Figure 3). Using all 12 samples as basic analytical units, the ubiquity of both common millet and rice was 50%, while that of foxtail millet was 17%.
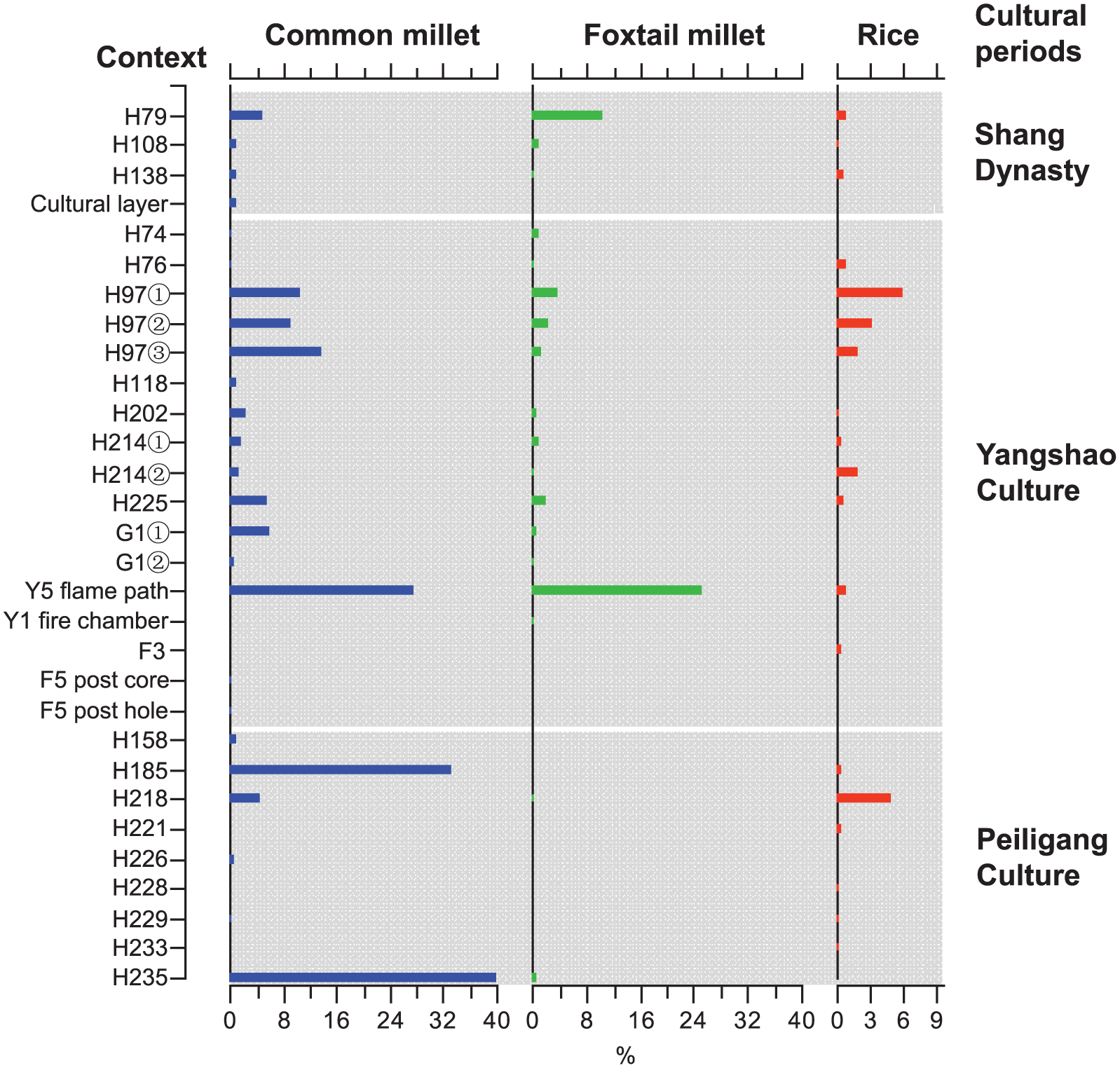
Variations in percentages of crop phytoliths among different periods at the Zhuzhai site.
Phytoliths from the Yangshao Culture period
A total of 13,874 phytoliths were encountered in 21 samples belonging to the Yangshao Culture, of which 17 samples contained crop phytoliths (Figure 3). In these samples, 592 phytoliths from common millet husks, 261 phytoliths from foxtail millet husks, and 133 rice phytoliths (including 115 double-peaks, 15 rice bulliforms, and three parallel-bilobes) were found. The common millet phytoliths were dominant in most samples with the highest percentage reaching 27.5%, while those of foxtail millet and rice were 25% and 6%, respectively (Figure 3). Using all 21 samples as basic analytical units, the ubiquity of common millet and foxtail millet was, respectively, 71.4% and 61.9%, while that of rice was 47.6%.
Phytoliths from the Shang Dynasty period
A total of 3109 phytoliths were identified from the four samples assigned to the Shang Dynasty period that contained crop phytoliths (Figure 3). Among these crop types, the most abundant was phytolith from the husks of foxtail millet (101 pieces), followed by common millet phytolith (66 pieces). Rice phytoliths (double-peaked and bulliform) were found in small quantities (15 pieces). The highest percentage of foxtail millet was 10.4%, while those of common millet and rice were 4.9% and 0.8%, respectively (Figure 3).
Phytoliths from the Warring States period
In total, 1346 phytoliths were counted in three samples from a burial (M36) belonging to the Warring States period. Unfortunately, no crop phytoliths were found in M36.
Figure 4 shows the temporal changes of relative proportions of phytolith from common millet, foxtail millet, and rice at the Zhuzhai site, indicating an evolution of mixed farming of millet and rice in the Central Plain. During the Peiligang and Yangshao Culture periods, common millet dominated all other crop types, with 92% and 60% proportions, respectively. Rice appeared in the Peiligang Culture period, but its proportions were always low (7–14%) during the three periods. Foxtail millet was present in the Peiligang Culture period in the fewest proportions (1%), but increased significantly to 26% in the Yangshao Culture period, exceeding that of rice. In the Shang Dynasty period, the proportions of foxtail millet further increased to 56%, replacing common millet (36%) as the dominant crop.
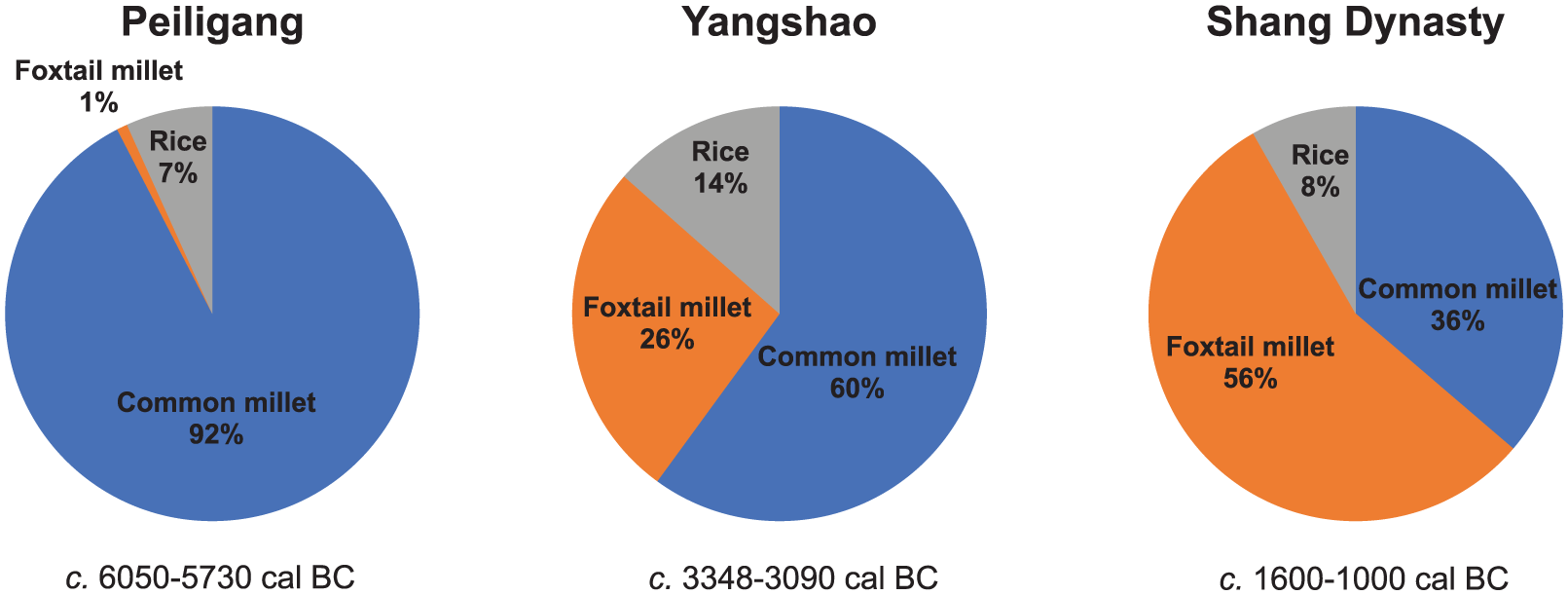
Proportions of phytolith from common millet, foxtail millet, and rice in different periods at the Zhuzhai site.
Morphometric analysis of millet phytolith showing millet domestication at the Zhuzhai site
According to observations, the Ω-type husk phytoliths from the foxtail millet in the samples were too broken to perform morphometric analysis. Therefore, information cannot be provided about the domestication of foxtail millet. Instead, η-type (which can be subdivided into ηI, ηII, and ηIII) husk phytoliths from common millet were more intact than those in the foxtail millet at the Zhuzhai site. In order to characterize the morphological variations of the structures of the η-type undulated patterns, four parameters were used: (1) W = width of ending interdigitation of the η-type undulated patterns, (2) H = length of undulation amplitude of the η-type undulated patterns, (3) R = ratio of the width of ending interdigitation to undulation amplitude (R = W/((H1 + H2)/2)), and (4) W1 = length of undulation amplitude and ending interdigitation of the η-type undulated patterns (Figure S1, available online). The measured average values of the parameters of the η-type undulated patterns are shown in Table 3. Figure 5 shows the bivariate biplots of above W, H, and R values by period, which all overlap in the data distribution of modern common millet within the limit of error, indicating that there is no statistically significant difference in phytolith morphology between the modern domesticated common millet and the ancient species. Moreover, the values of H and W1 both show an increasing trend from the Peiligang Culture to the Shang Dynasty (Figure 6), indicating the average size of the η-type undulated patterns of common millet gradually gets larger over time at the Zhuzhai site.
Measured data of the parameters of the η-type undulated patterns of common millet from the Zhuzhai site.
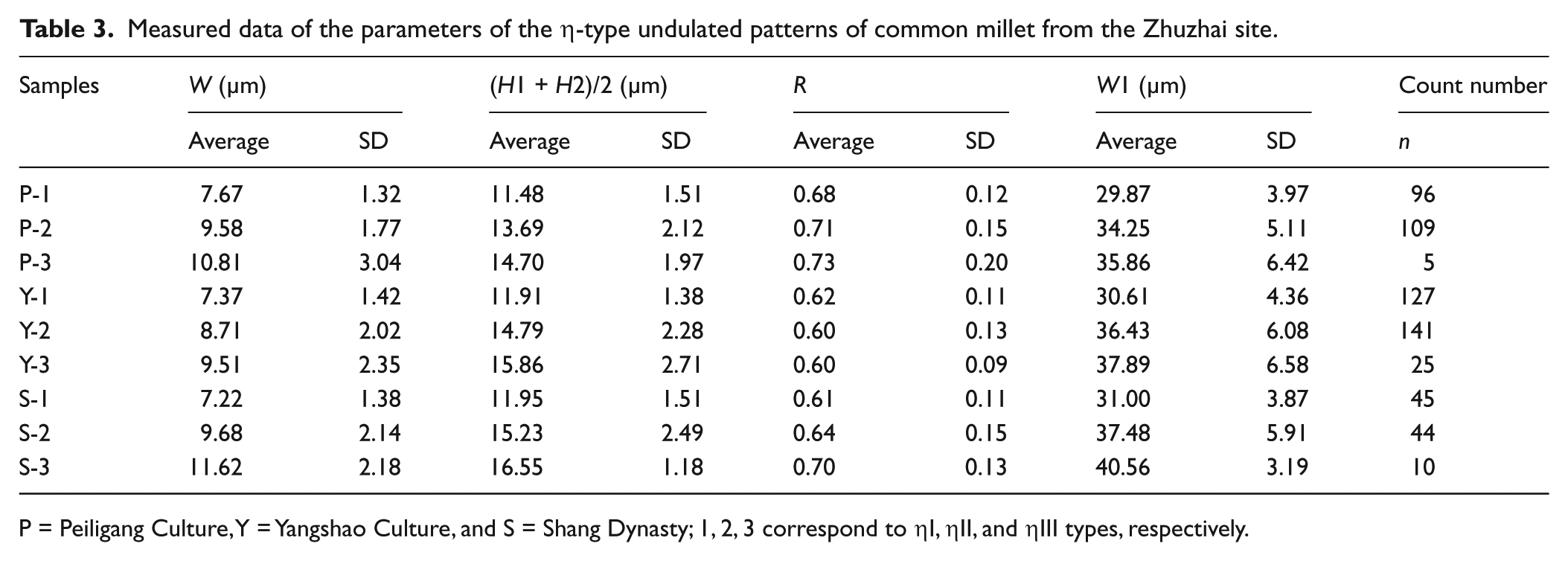
P = Peiligang Culture, Y = Yangshao Culture, and S = Shang Dynasty; 1, 2, 3 correspond to ηI, ηII, and ηIII types, respectively.
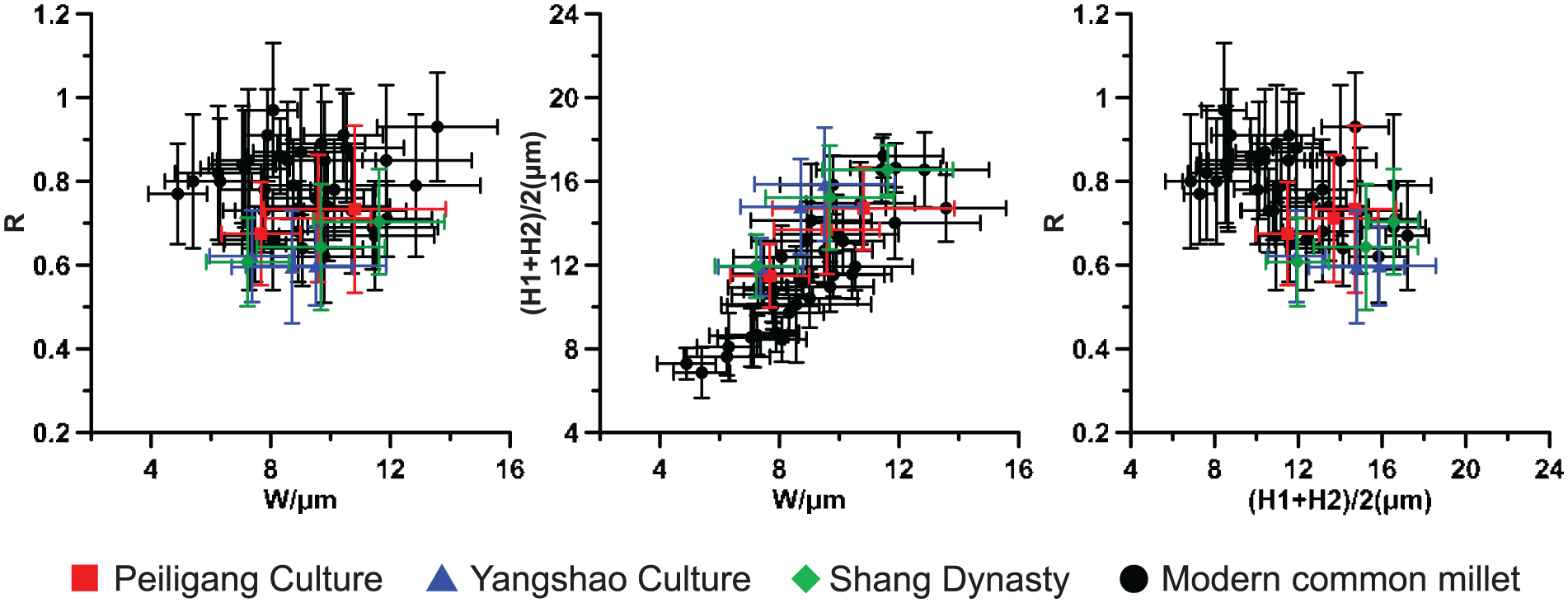
Comparison of η-type undulated patterns for common millet between three periods and modern time. Data of modern common millet (domesticated species = 12) are adapted from Lu et al. (2009b).

Box plot of (a) H and (b) W1 values of η-type undulated patterns in three periods at the Zhuzhai site. ηI, ηII, and ηIII represent three levels of the undulation amplitude.
Measurement of early rice phytoliths showing the nature of rice at the Zhuzhai site
The presence of rice phytoliths assigned to the Peiligang and Yangshao Culture periods is an important finding at the Zhuzhai site, though the contents of the samples are relatively low. Previous studies have suggested that the morphological characteristics of rice phytolith appear to be under genetic control, and therefore reflect taxonomical significance (Gu et al., 2013; Ma and Yamaji, 2006; Zheng et al., 2003), making their nature (wild/domesticated or japonica/indica) determination feasible. In the last 20 years, scholars have established several criteria for determining the nature of rice phytoliths. Such methods have played a vital role in the identification of rice remains from archaeological sites (Ball et al., 2016). In the present study, three methods were applied for this identification: (1) morphometrics of rice double-peaked phytolith (Gu, 2009; Zhao et al., 1998), (2) morphometrics of rice bulliform phytolith (Wang and Lu, 2012; Zheng et al., 2003), and (3) number of scale-like decorations of rice bulliform phytolith (Lu et al., 2002; Huan et al., 2015) (Figure S2, available online). Although there existed some doubt as to the effectiveness of these methods (Fuller and Castillo, 2014; Fuller et al., 2010), they were used as an alternative methodology aside from using rice macroremains.
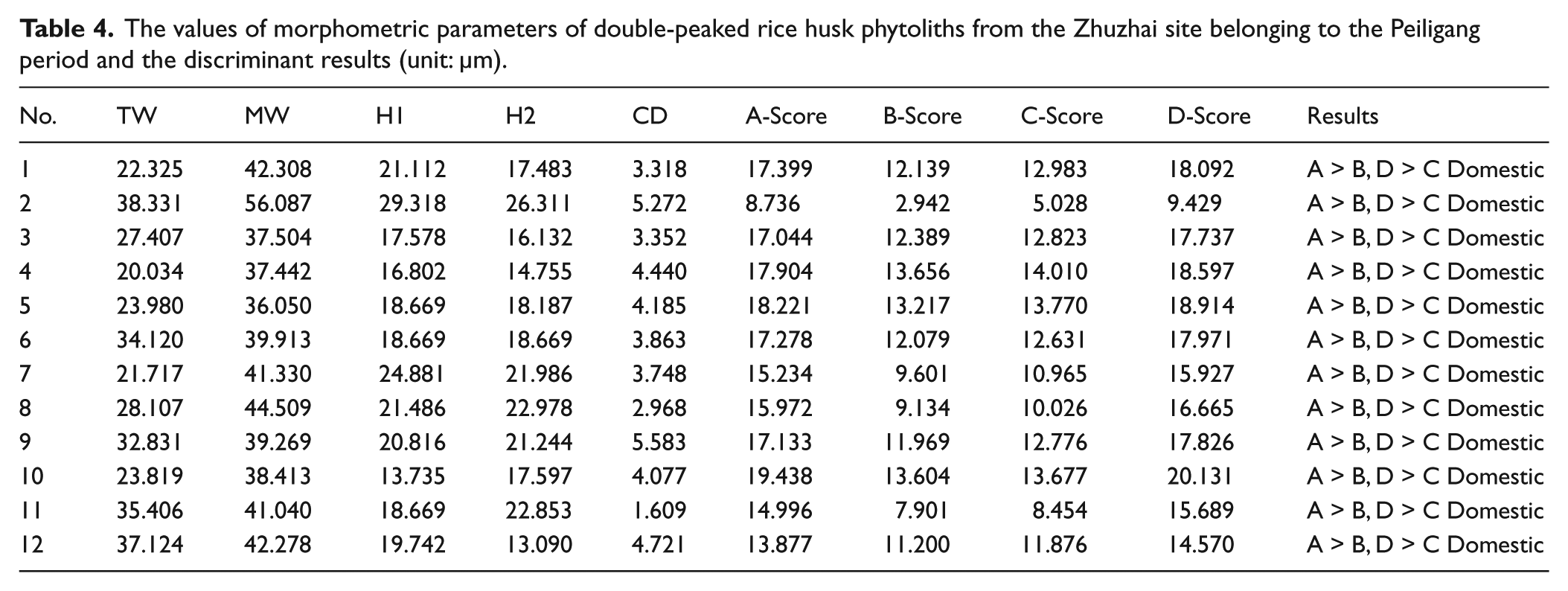
For the Peiligang Culture period, the double-peaked glume cell was a relatively abundant morphotype of rice phytolith, and thus was used as the main material for determination. A total of 12 double-peaked phytoliths were selected, and their shape measurements were incorporated into the formula used to distinguish wild from domesticated rice, derived from Zhao et al. (1998). The results indicate that the predicted values of the 12 double-peaked phytoliths for domesticated rice are greater than those for the wild rice (Table 4), thus deducing that these phytoliths are from domesticated rice. Additionally, although only three pieces of rice bulliform phytolith with clear scale-like decorations were found in the Peiligang Culture period, the number of scale-like decorations is 9, 10, and 12, respectively. This concurs with the diagnostic feature of the domesticated Oryza species observed by Lu et al. (2002) (Figure S2c, available online).
The values of morphometric parameters of double-peaked rice husk phytoliths from the Zhuzhai site belonging to the Peiligang period and the discriminant results (unit: µm).
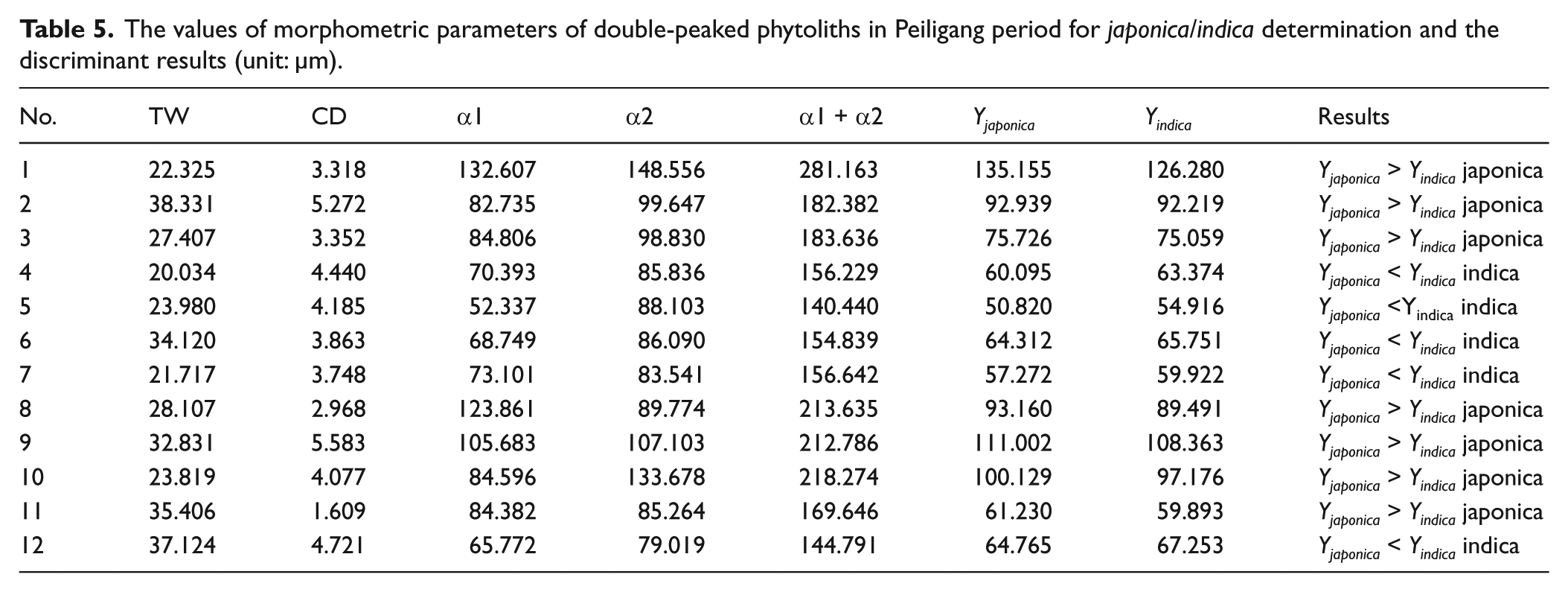
Given that the double-peaked phytoliths in the Peiligang period are determined to be a domestic type, further investigation into what subspecies of domesticated rice, Oryza sativa L. indica or Oryza sativa L. japonica, was cultivated can be performed. Four parameters (TW, CD, α1, and α2; Figure S2a, available online) of the above-mentioned 12 double-peaked phytoliths were measured. The observed data were inserted into a discrimination formula established by Gu (2009), and the scores (Y) of japonica and indica were obtained. If Yjaponica > Yindica, the phytolith was identified as japonica, and vice versa. The results show that seven pieces are determined to be japonica, accounting for 58.3% of the selected double-peaked phytoliths (Table 5). Therefore, the subspecies of Oryza sativa cultivated at the Zhuzhai site in the Peiligang Culture period is most likely japonica.
The values of morphometric parameters of double-peaked phytoliths in Peiligang period for japonica/indica determination and the discriminant results (unit: µm).
For the Yangshao Culture period, although double-peaked phytoliths were abundant, there were few morphotypes with distinguishable form available for morphometric analysis. Instead, rice bulliform phytoliths were used as the major material for the determination. A total of 50 bulliform phytoliths were measured. The number of scale-like decorations on them ranged from 5 to 14, and 27 individual phytoliths had nine or more scale-like decorations (Figure S3, available online), accounting for 54% of the selected phytoliths. This proportion of bulliform phytoliths with ≥9 decorations is similar to the proportion found in modern domesticated rice soil (63.70% ± 9.22%) (Huan et al., 2015), and therefore, these microfossils are designated as domesticated rice. Moreover, the morphological characteristics (VL and HL; Figure S2b, available online) of over 50 rice bulliform phytoliths (VL: 44.91 ± 7.82 µm, HL: 39.16 ± 8.28 µm) are not only larger in size than those from the Tanghu site (VL: 39.3 ± 5.0 µm, HL: 33.9 ± 4.4 µm) (5800 cal. BC) and the Majiabang Culture (VL: 36.11 µm, HL: 30.68 µm) (5000–4000 cal. BC), but also exceed those from the Songze (VL: 41.58 µm, HL: 35.34 µm) (4000–3300 cal. BC) and Liangzhu cultures (VL: 41.79 µm, HL: 34.28 µm) (3500–2200 cal. BC) (Zhang et al., 2012; Zheng et al., 1999). The large size of the bulliform phytoliths further confirms the domestic nature of the rice in the Yangshao Culture period.
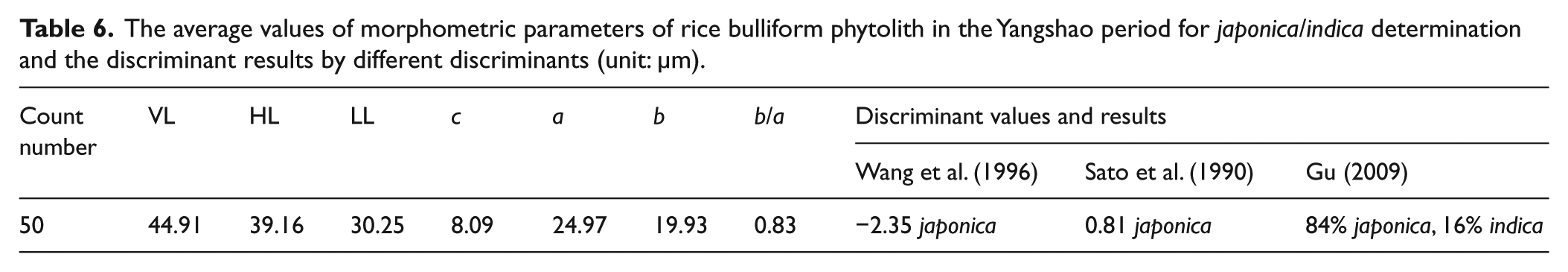
Two methods, including b/a ratio and discriminant analysis (Wang and Lu, 2012), were applied to the above-mentioned bulliform phytoliths to distinguish between japonica and indica through the measuring of six parameters (Figure S2b, available online). Generally, the mean b/a ratio of japonica-type bulliforms is <1, while that of indica-type bulliforms is >1. Three discriminants (see Gu (2009), Sato et al. (1990), and Wang et al. (1996) for detailed formula) can reliably identify japonica and indica, and when the average values of parameters are inserted into these formulas, the discriminant values and results are obtained. In our samples, the mean b/a ratio is 0.83 (Table 6), which indicates that the cultivated rice is japonica. This is supported by the results derived from three discriminants (Table 6). Thus, the subspecies of rice cultivated at the Zhuzhai site in the Yangshao Cultural period is still japonica.
The average values of morphometric parameters of rice bulliform phytolith in the Yangshao period for japonica/indica determination and the discriminant results by different discriminants (unit: µm).
The rice arable systems at the Zhuzhai site reflected by phytoliths
The presence of bulliform and parallel-bilobe phytoliths from rice leaf/stem suggests that the Zhuzhai rice was likely to be grown locally, making the assessment on the cultivation ecology of this rice feasible. The phytoliths from the Peiligang Culture show more than 50% are fixed morphotypes, and the sensitive to fixed ratio is 0.92 (Figure 7). In the later phases, such as the Yangshao and Shang periods, the phytoliths have many more fixed forms and a lower sensitive to fixed ratio than those of the Peiligang Culture (Figure 7). The overall sensitive to fixed ratios at the Zhuzhai site decreased through time (Figure 7), indicating a decrease in water abundance in each arable system.
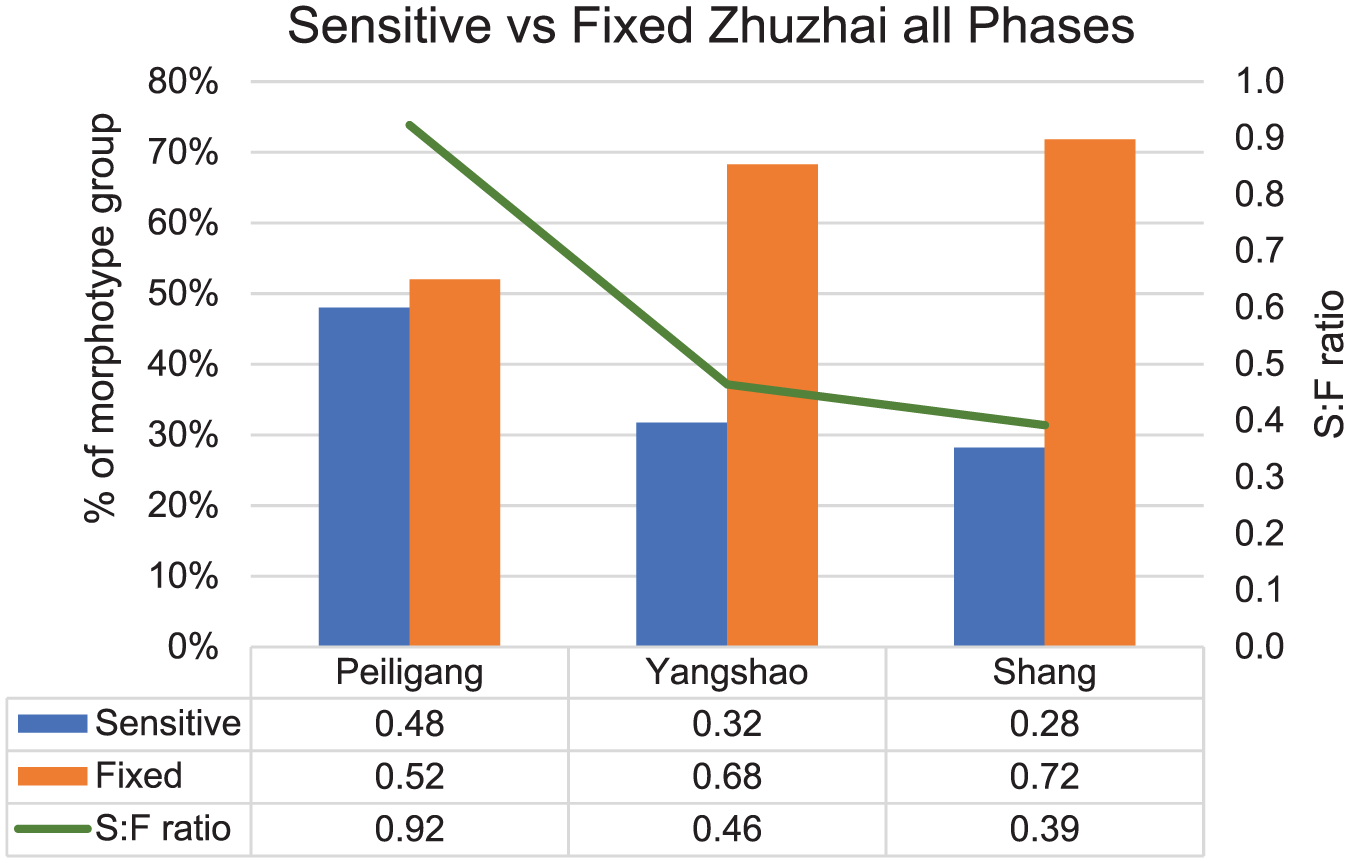
Percentage of sensitive versus fixed phytolith morphotypes in three periods at the Zhuzhai site; percentages exclude all phytolith types not within the sensitive/fixed classification as defined by Weisskopf et al. (2015b).
Charred plant remains
A total of 180 charred plant remains were found from six soil samples (49 L), of which 164 were seeds assigned to 23 plant taxa (Table 7). All of the 164 remains can be classified into three categories, comprising three species of crops, eight species of fruits or nuts, and 12 species of weeds (Table 7). The remaining plant remains, such as plant fragments and unknown seeds, were recorded as ‘others’, and could not be unambiguously identified because of poor preservation or lack of clear identification features. Representative taxa identified from the site are illustrated in Figure 8.
Charred plant remains recovered from the Zhuzhai site.


Representative charred plant remains from the Zhuzhai site: (a) Panicum miliaceum (H158), (b) Setaria italica (H202), (c) Oryza sativa (H158), (d) Setaria viridis (H225), (e) Eleusine indica (H208), (f) Setaria sp. (H218), (g) Chenopodium sp. (H225), (h) Vitis sp. (H208), (i) Physalis sp. (H225), (j) Zanthoxylum sp. (H226), (k) Melilotus sp. (H225), (l) Celtis koraiensis (H208), (m) Broussonetia papyrifera (H226), (n) Ziziphus jujuba (H225), (o) Juglans mandshurica (H158), (p) Quercus sp. (H226), (q) charred stalks (H158), and (r–t) unknown seeds (H218, H208, H202).
Charred plant remains from the Peiligang Culture period
A total of 64 charred plant remains were found in four pit samples (30 L). Among these, the most important finding was one charred rice grain (Figure 8c) from the H158 pit (5986–5786 cal. BC). This is the first macrobotanical evidence of rice belonging to the Peiligang Culture in the Central Plain. The rice grain is 3.56 mm in length, 1.57 mm in width, and 1.43 mm in thickness, and the length to width ratio is 2.17, which is similar to the rice from the Xihe site located in the western Shandong Province (Jin et al., 2014). The other crops were also present in low quantities, including five grains of common millet and six grains of foxtail millet. The crops made up a small proportion in all remains (18.75%), while fruits and nuts such as Celtis koraiensis (Figure 8l), Broussonetia papyrifera (Figure 8m), Vitis sp. (Figure 8h), Juglans mandshurica (Figure 8o), and Quercus sp. (Figure 8p) occupied 37.5% of the plant assemblages of the Peiligang Culture period. The assemblage of weeds included Eleusine indica (Figure 8e), Cuscuta sp., Setaria viridis, Galium sp., and Cyperaceae, accounting for 31.25% of all plant remains.
Charred plant remains from the Yangshao Culture period
A total of 116 charred plant remains were found in two pit samples (19 L), of which 71 were from foxtail millet and five were assigned to common millet. No rice grains were recovered, and this is possibly because of the limited sample size. The crops became the most abundant type (65.5%) in the plant assemblages of the Yangshao Culture period, while wild fruits became rare, not only in quantities (1.7%) but also in species (just two types: Physalis sp. and Ziziphus jujuba) (Figure 8i and n). Weeds, such as Fabaceae, Setaria sp., Chenopodium sp. (Figure 8g), Melilotus sp. (Figure 8k), and Carex sp., made up 25.9% of all of the remains.
Discussion
Millet farming at the Zhuzhai site
The results indicate that the millets were staple crops in the Zhuzhai site’s mixed farming during the Neolithic-Bronze Age, but the relative proportion of foxtail millet to common millet experienced a temporal change.
For the Peiligang Culture period, there were six foxtail millet seeds and five common millet seeds recovered from the Zhuzhai site (Table 7), but these charred remains were too few to reflect the relative proportion of the two millets. Phytolith analysis provided rich millet remains, and its results showed that the ratio of common millet phytoliths was much more than that of foxtail millet (Figures 3 and 4). This suggests that millet farming in the Central Plain was mainly based on the cultivation of common millet during the Peiligang Culture from 6050–5730 cal. BC. This archaeobotanical evidence, together with other studies from the Xinglonggou (Zhao, 2004), Yuezhuang (Crawford et al., 2006, 2013), Dadiwan (Liu et al., 2004), Cishan (Lu et al., 2009a), Tanghu (Zhang et al., 2012), Zhangmatun (Wu et al., 2014a), Bianbiandong (Sun et al., 2014), and other sites in the central Gansu Province (An et al., 2010), further demonstrates that the whole North China region was dominated by common millet in the early Neolithic stage, while foxtail millet was scarcely present.
Previous macrofossil studies revealed that foxtail millet finally replaced common millet as a staple food in the Yangshao Culture after around 4000 cal. BC in North China (Liu et al., 2008; Qin, 2012; Zhao, 2014), especially in the Central Plain (Fuller et al., 2007b; Lee et al., 2007; Qin, 2009; Zhang et al., 2014). However, the phytolith results from this research indicate that common millet was still more significant than foxtail millet in the late Yangshao period at the Zhuzhai site (3348–3090 cal. BC), though the ratio of foxtail millet did remarkably increase (Figure 4). Similarly, evidence from numerous sites in this stage such as Gongjiachuan, Wujiapo, and Yanggu in east Gansu (4500–3000 cal. BC) (Zhou et al., 2011); Lijiatai and Hulijia in Qinghai (4000–3000 cal. BC) (Jia, 2012); Gouwan in Henan (5000–3000 cal. BC) (Wang et al., 2011); Beiqian in Shandong (4200–3500 cal. BC) (Jin et al., 2016); and Quanhu, Wangjiazui, Yangguanzhai, Shuigou, Huxizhuang, and Anban in Shaanxi (4000–3000 cal. BC) (Zhang et al., 2010b) also revealed that the content of common millet was larger than that of foxtail millet, suggesting that the common millet-based dry farming not only continued but also widely existed in North China during the Yangshao period. This means that the transformation from a common millet-dominated pattern to a foxtail millet-dominated pattern of millet farming in the North (Liu et al., 2008; Qin, 2012) may not have been very common in the Yangshao period (4000–3000 cal. BC).
Because of the lack of Longshan and Erlitou Culture sediments, the structure of millet cultivation at the Zhuzhai site during these periods was unclear. However, by the last phase of the Shang Dynasty period, foxtail millet had outnumbered common millet according to the phytolith record (Figure 4). This result was in accordance with plant macrofossil evidence from other Shang Dynasty sites in the Central Plain (Fuller et al., 2007b; Lee et al., 2007; Zhang et al., 2014; Zhao, 2011b), suggesting that foxtail millet had certainly become the dominant crop in millet farming during this stage.
In addition, it is necessary to note that in some instances, charred remain and phytolith analyses were both performed in the same sites or areas, focusing on the relative proportion of foxtail millet versus common millet, but their results were always inconsistent. Usually, the proportion of foxtail millet was much higher than that of common millet in charred plant assemblages, while phytolith records revealed opposite result. An example of this is found in the studies of the Lajia site (Wang et al., 2015; Zhao, 2003) and the Guanzhong Basin (Zhang et al., 2010b; Zhao, 2011a). The same situation also can be observed in samples H202 and H225 from the Yangshao pits at the Zhuzhai site (Figure 3 and Table 7). An experimental study has found that the same weight of common millet and foxtail millet yield almost the same amount of phytoliths within the same experimental approach (wet oxidation or dry ashing) (Zhang et al., 2010b). This means that the foxtail millet and common millet phytoliths have an equal chance of being preserved under the same circumstances, and the quantity of phytoliths reflects the relative production of the two millets, rather than the quantity of individual grains collected by flotation. However, in the Yangshao samples of the Zhuzhai site, even though the flotation results were adjusted, the relative production of foxtail millet (n = 31) was still much higher than that of common millet (n = 5), which remained different from the phytolith results. A similar case also occurred at the Lajia site (Wang et al., 2015). Thus, the difference between phytoliths and charred remains may be caused by other factors. For example, whether the two millets have different likelihoods of being charred in archaeological contexts has attracted the attention of scholars.
Charring is an ideal and widespread case for the preservation of seeds at archaeological sites (Liu et al., 2008). The charring of seeds requires exposure to heat, and is determined by the consistency and texture of the seeds themselves. Experiments on the charring of the seeds of the two types of millets indicated that the charring temperature range of common millet was lower than that of foxtail millet under any circumstances (Märkle and Rösch, 2008). Thus, the common millet seeds have a lower probability of being preserved than that of foxtail millet at sites during the charring process, and the amount of common millet would be underestimated against foxtail millet in charred plant assemblages. Given this taphonomic bias in charred millet remains, the plant macrofossil analyses may overestimate the ratio of foxtail millet in millet farming and their results may be problematic. Considering the quantity of phytoliths reflects the real relative production of the two millets, the plant macroremain results need to be tested by phytolith analysis when determining the millet farming patterns.
Millet domestication at the Zhuzhai site
Phytolith evidence from the Cishan site in the North China Plain indicated that possibly the earliest domestication of common millet dated back to 8000 cal. BC (Lu et al., 2009a). However, clues for common millet domestication in the start of the Holocene were still rare, especially in the plant macroremains (Qin, 2012). From 6500 to 4000 cal. BC, an essential stage for millet domestication (Fuller et al., 2014), the size of common millet grains from Xinglonggou (averaging 1.6 mm long, 1.2 mm wide) (Zhao, 2004), Dadiwan (averaging 1.3–2.1 mm long, 1.1–1.9 mm wide, and 0.8–1.9 mm thick) (Liu et al., 2004), Zhangmatun (averaging 1.7 mm long, 1.3 mm wide, and 1.2 mm thick) (Wu et al., 2014a), Yuezhuang (averaging 1.5 mm long, 1.0 mm wide, and 1.0 mm thick) (Crawford et al., 2013, 2016), and the Peiligang samples of the Zhuzhai site (averaging 1.6–1.9 mm long, 1.4–1.9 mm wide) partially overlapped with the modern domesticated specimens that produce a larger, more spherical grain shape (Zhao, 2014), implying that these remains indicate the early morphology of domesticated common millet. Our morphometric analysis of common millet phytoliths also suggested that common millet with domesticated traits have occurred at the Zhuzhai site since the Peiligang Culture period (Figure 5). Additionally, the two phytolith parameters (H and W1), as mentioned above, revealed that the average size of η-type undulated patterns of common millet at the Zhuzhai site became increasingly larger from the Peiligang Culture to the Shang Dynasty (Figure 6). This may have been caused by the increase in grain size, a remarkable result of plant domestication (Fuller, 2007; Fuller et al., 2014; Zhang et al., 2011). Hence, the phytolith data suggested that the process of common millet domestication at the Zhuzhai site began in the Peiligang period and continued up to the Shang period, and the degree of domestication gradually became higher over a long time. However, since the increases in η-type phytolith of ancient common millet fell within the morphological range of modern domesticated varieties (Figure 5), then the increasing trend of η-type undulated patterns of common millet shown in Figure 6 might also be the result of the cultivation of different domesticated varieties in each period.
The earliest plant remains of the domesticated foxtail millet were recovered from the Nanzhuangtou (Yang et al., 2012), Donghulin (Yang et al., 2012; Zhao, 2014), and Zhuannian (Yang et al., 2014) sites in the North China Plain from 9000–7500 cal. BC, suggesting that the beginning of foxtail millet domestication predated that of common millet (Larson et al., 2014). But their phytolith morphology in the early stage of the Cishan site (6700 cal. BC) was still close to the green foxtail (Zhang, 2010), indicating that foxtail millet was still undergoing domestication during the early Holocene. The size of the foxtail millet grain from the Yuezhuang site (averaging 1.2 mm long, 1.2 mm wide, and 0.9 mm thick; 6000–5700 cal. BC) (Crawford et al., 2013, 2016) and the Peiligang samples from the Zhuzhai site (averaging 1.5 mm long, 1.0 mm wide, and 1.0 mm thick; 6050–5730 cal. BC) fell within the range of variation of the later domesticated type (Crawford et al., 2016), while the phytolith and grain morphology of foxtail millet in the late stage of the Cishan site (5500 cal. BC) and the Xinglonggou site of the Xinglongwa Culture (6200–5400 cal. BC) was closer to the domesticated specimens (Zhang, 2010; Zhao, 2004). Both of these facts indicate that foxtail millet had already gone through a significant degree of domestication by 6000–5500 cal. BC. As mentioned above, substantial data could not be provided to illustrate the domestication process of foxtail millet at the Zhuzhai site in detail, but it is speculated that this process may be protracted according to the starch analysis of sites in the West Liaohe region, which have suggested that millet domestication proceeded gradually over a long period from approximately 6500 to 3000 cal. BC (Ma et al., 2016b).
Rice cultivation at the Zhuzhai site
The earliest presence of rice in North China was confirmed by the plant remains from the Jiahu (Zhao and Zhang, 2009) and Baligang sites (Deng et al., 2015) in the southern Henan Province and can be dated to as early as ca. 7000–6500 cal. BC. It subsequently spread into the lower Yellow River Houli Culture at ca. 6000 cal. BC, as suggested by the archaeobotanical studies of the Yuezhuang and Xihe sites in western Shandong (Crawford et al., 2006, 2013, 2016; Jin et al., 2014). Furthermore, the discovery of rice phytoliths and starch grains at the Tanghu site (ca. 5800 cal. BC) provided the sole evidence to date of rice cultivation in the Central Plain during the Peiligang Culture period (Yang et al., 2015; Zhang et al., 2012) The rice remains assigned to the Peiligang Culture (ca. 6050–5730 cal. BC) at the Zhuzhai site confirmed the findings of the Tanghu site. The rice was most likely grown there, rather than being a traded good because of the presence of bulliform and parallel-bilobe phytoliths from rice leaf/stem. This certainly indicated that rice had been cultivated in the Central Plain at ca. 6000 cal. BC, corresponding to the first northward expansion of rice resources in China (Qin, 2012).
There are two different opinions about the source of the 8000-year-old rice cultivated in the dry farming areas of Yellow River valley. The first idea considered that the rice cultivation developed from early regional wild rice collecting (Fuller et al., 2010; Qin, 2012; d’Alpoim Guedes et al., 2015), while the second idea regarded it as introduced by migrants from other areas such as the middle Yangtze and Huai River valley (Zhang, 2011b; Zhang et al., 2012; Zhang and Hung, 2013). Whether it came from the surrounding wild rice or was brought from different location, this early occurrence of rice cultivation in the Yellow River valley clearly benefited from the Holocene megathermals in China. During this time, the temperature was ca. 2–4°C higher than at present (Ge et al., 2007; Wang and Gong, 2000) and precipitation increased because of the expansion of monsoon circulation (Dykoski et al., 2005; Xiao et al., 2004). The rising temperatures and moisture levels offered favorable conditions for wild rice populations to spread inside the Yellow River valley, (d’Alpoim Guedes et al., 2015) and also provided great opportunities for the technical exchange of rice cultivation.
With regard to the cases at the Zhuzhai and Tanghu sites (Zhang et al., 2012), the selected rice phytoliths of the Peiligang Culture were determined to be a domesticated type, indicating that rice was already in the process of domestication during the Peiligang Culture period. At the same time, rice domestication in the Yangtze had not finished (Fuller et al., 2009). Therefore, the early rice in the Central Plain was unlikely to be brought from the Yangtze, but rather was a result of the exploitation of the surrounding wild rice, as was the case with the Houli Culture (d’Alpoim Guedes et al., 2015). However, it was speculated that this may have been influenced by the Baligang site to the south of the Central Plain, in which the cultural traditions varied between north and south. The domesticated rice spikelet bases dominated in the pre-Yangshao occupation at the Baligang site before 6300 cal. BC, indicating an earlier domestication than in the lower Yangtze (Deng et al., 2015). The utilization experience of rice at the Baligang site could have been introduced into the Central Plain through cultural communications, and such communications could have facilitated the practices of rice domestication in the Central Plain.
Because of the lack of early Yangshao Culture sediments at the Zhuzhai site, it could not be determined whether rice was absent from 5000–4000 cal. BC, which was once considered as the southward retreat of rice cultivation to the Yangtze (Qin, 2012). According to the evidence at the Tanghu site, however, rice continued to be cultivated from 5800 to 2500 cal. BC in the Central Plain (Zhang et al., 2012). During the middle and late Yangshao period (4000–3000 cal. BC), the rice was widely grown in the Yellow River valley (Wu, 1994), while rice farming based on the fully domesticated species began to spread northward from the lower and middle Yangtze (Fuller et al., 2009; Qin, 2012). Considering the temporal-spatial route for the northward spread of rice cultivation as drawn by scholars (Qin, 2012; Zhang et al., 2010, 2012), the indigenous rice agriculture of the Central Plain may have been impacted by the rice-farming culture from the middle Yangtze. This assumption was also supported by the larger rice bulliform phytoliths at the Zhuzhai site in the late Yangshao period, which may reflect a higher degree of rice domestication than from the Songze and Liangzhu Cultures in the lower Yangtze. Its proportion of rice bulliform phytoliths with ≥9 scale-like decorations reached 54% during the late Yangshao period, while this proportion just became 59% till the Eastern Zhou Dynasty (770–256 BC) in the lower Yangtze (Huan et al., 2014). Meanwhile, more advanced rice farming in the middle Yangtze is probable in comparison with the lower Yangtze, as some scholars have suggested (Fuller, 2011; Nasu et al., 2012; Qin, 2012).
Although rice had been cultivated at the Zhuzhai site as early as 6000 cal. BC, it was always relatively rare in the sequence (Figure 4). The same situation also occurred in the Yiluo valley (Lee et al., 2007), upper Ying valley (Fuller et al., 2007b; Zhang et al., 2010a), Luoyang basin (Zhang et al., 2014), and the Tanghu site (Zhang et al., 2012), indicating that rice always constituted a negligible proportion of food production in the Central Plain at least until the Shang period. This means that the dominance of millet farming in the region did not change after the introduction of rice cultivation. Despite this, the occurrence of rice farming in the Central Plain certified that the earliest formation of mixed millet and rice agriculture extended to 8000 years ago, and its birthplace was likely located in North China. No millet has been found at the sites located in South China such as the Shunshanji site in the Huai River valley (Luo et al., 2016; Yang et al., 2016b) and the Xiaohuangshan site in the lower Yangtze (Yao et al., 2016), where rice has been cultivated. The earliest occurrence of mixed agriculture in South China may be 4500 cal. BC, when millets were introduced into the middle and lower Yangtze (Deng et al., 2015; Zhao and Jiang, 2016).
The implication of the nature and arable system of rice at the Zhuzhai site
Lee et al. (2007) have pointed out that the charred rice remains are too few in the Central Plain, and there is no way to determine the nature and arable systems of the rice grown there. In this study, phytolith analysis was used to address these issues. The results showed that (1) the early rice at the Zhuzhai site was domesticated japonica type, and (2) rice farming at the Zhuzhai site used consistent dryland systems.
The implication of domesticated japonica at the Zhuzhai site lies in two points. First, it provides a new reference for the debate about the origin time of domesticated rice. On one hand, the rice remains from the Shangshan, Kuahuqiao, Xiaohuangshan, Jiahu, and Baligang sites (7000–6000 cal. BC) were identified as domesticated (Deng et al., 2015; Liu et al., 2007; Zheng et al., 2007; Zuo et al., 2017; Zhao and Zhang, 2009); on the other hand, Fuller et al. (2007a, 2008) argued these remains were mostly wild rice, and even that of the Majiabang and Hemudu Cultures after 5000 cal. BC showed few signs of domestication. The 8000-year-old rice remains from the Yellow River region complicates this problem. In the case of the Yuezhuang and Xihe sites, the discoverer did not clearly define the nature of the rice remains (Crawford et al., 2006, 2013; Jin et al., 2014), and therefore whether they were wild or domesticated is controversial (Fuller et al., 2010; Qin, 2012; Zhang, 2011b) and it could not provide valid evidence in defining the origin time of domesticated rice. In contrast, it is certain that domesticated rice assigned to the Peiligang Culture was recovered from the Zhuzhai site, indicating that the Central Plain, a region far away from the Yangtze, had already used domesticated rice 8000 years ago. Hence, the earliest domesticated rice in the Yangtze River valley occurs at least 6000 cal. BC or earlier. Without doubt, the earliest appearance of domesticated rice is only the beginning of the long process of rice domestication, and the rice remains in the Yangshao period at the Zhuzhai site have shown a high degree of domestication, indicating that the rice domestication of the Central Plain may have been completed during 4000–3000 cal. BC. This timeframe is consistent with that of the Yangtze (Fuller et al., 2009; Ma et al., 2016a).
Second, it provides support for the hypothesis that China is the origin of japonica. According to new genetic studies, although Asian rice subspecies originated from multiple wild rice populations, the key genomes in domestication occurred only once in japonica and introgressive hybridization from early japonica to proto-indica led to domesticated indica rice (Choi et al., 2017). Some scholars further suggested that this early japonica originated from China, and that until 3000 cal. BC, indica only occurred in India (Fuller, 2011; Fuller and Sato, 2008; Huang et al., 2012; Gross and Zhao, 2014). Therefore, it is speculated that the early domesticated rice in China is only the japonica type. In addition, domesticated rice remains from early Neolithic Shangshan, Xiaohuangshan, Kuahuqiao, and Jiahu sites (Chen, 1997; Zheng et al., 2007); middle Neolithic Longqiuzhuang, Luojiajiao, and Caoxieshan sites (Udatsu et al., 1998; Zheng et al., 2000; Zou et al., 1998); late Neolithic Liangchengzhen site (Jin et al., 2004); and Zhuzhai site in Peiligang and Yangshao periods were all identified as japonica, thus supporting the above speculation. Therefore, as long as the rice from Neolithic China is determined to be domesticated, it is certainly japonica rather than another subspecies.
Previous studies have demonstrated that rice arable systems can be reflected by a ratio of sensitive versus fixed grass phytolith morphotypes (Weisskopf, 2016; Weisskopf et al., 2014, 2015a, 2015b). The fixed (dry) morphotypes are those which normally silicify and are considered genetically predisposed to form phytoliths. Sensitive types are formed only when much water is available, especially from later in the plant’s life cycle (Weisskopf et al., 2015a, 2015b) In the Zhuzhai site, sensitive types were always fewer than fixed types (Figure 7), suggesting the rice cultivation was carried out in a dry field system during the Neolithic-Bronze Age. It is also supported by no or few weed indicators of paddy fields such as Echinochloa or Cyperaceae in the samples (Table 7).
This dry field system mainly depends on having enough annual rainfall (>800 mm) (Fuller et al., 2011), so the paleorainfall reconstruction is crucial for understanding the rice arable system in the Zhuzhai site. Phytolith data from the Weinan section on the same latitude as Zhengzhou suggested the reconstructed mean annual precipitation at 10.5–2.8 ka was 745 ± 65 mm (maximum 870 mm) (Lu et al., 2007), which could sustain the dry-cropped rice in the Neolithic-Bronze Age. Moreover, the Zhuzhai site is located where two rivers meet, and its rice may be cultivated alongside the river in order to use the ground water or seasonal flood. Without irrigation, this arable system is less labor-intensive and subsequently gets lower grain yields, accounting for the small ratios of rice in the crop assemblages. It is likely a farming strategy to adapt to the available field where millets were grown, and fortunately, the water status could satisfy the demand, and therefore, it would not be necessary to make an effort to prepare a new wet field for rice. The Zhuzhai site is also an ordinary settlement without complex social organization, large population, and intensive land use practices, providing small opportunity for cultivating rice in paddy fields, which needs sufficient labor force and effective maintenance.
By contrast, the earliest paddy fields in the Central Plain have been found at the Huizui site, dating to the middle Yangshao Period (Rosen et al., 2017). This rice paddy farming facilitated population growth and social complexity, making the Huizui site a regional center in the Yiluo River Basin (Liu et al., 2005; Rosen et al., 2017). At Zhaojiazhuang in Shandong, another Neolithic paddy field in North China was found and dating to ca. 2000 BC (Jin et al., 2007). However, because of the limited research conducted on the determination of rice arable systems in North China, the diversity of ancient rice planting patterns could not be discussed. More studies are needed to complete the picture of rice farming during the Neolithic-Bronze Age.
The place of agriculture in subsistence economy at the Zhuzhai site
In the Central Plain, the Peiligang Culture has long been regarded as having achieved a significant level of agriculture (Li, 2003), but new records show that the gathering of wild plants dominated the subsistence economy and that agriculture was only a minor component during this stage (Liu et al., 2013, 2010; Zhang et al., 2011). At the Zhuzhai site, there were many kinds of wild fruits and nuts recovered from the Peiligang Culture contexts, and their proportion was larger than that of crops (Table 7), which supports the latter opinion. The crops finally dominated the plant assemblages of the Zhuzhai site during the Yangshao period, with wild edible plants becoming more sporadic and almost disappearing (Table 7), implying that agriculture had replaced gathering and became dominant in subsistence economy at ca. 3300–3000 cal. BC.
The developmental trajectory of agriculture in subsistence economy at the Zhuzhai site was similar to that of farming in the adjacent Yiluo valley (Lee et al., 2007). Taken together, the trend of agricultural development in the Central Plain was completely synchronous to the developmental paths of agriculture in North China (Wang et al., 2016).
Conclusion
The archaeobotanical evidence discussed from the Zhuzhai site verifies that mixed millet and rice agriculture has been established in the Central Plain about 8000 years ago. Common millet was used as the staple crop in the Peiligang and Yangshao periods, and was replaced by foxtail millet in the Shang period. Rice has occurred since the Peiligang Culture, but apparently was not an important crop throughout time. The process of common millet domestication in the Central Plain began at the latest in the Peiligang period and continued up to the Shang period. Foxtail millet may have gone through a significant degree of domestication in this region during ca. 6000–5500 cal. BC, but its domestication process was still unclear. Rice assigned to the Peiligang and Yangshao periods was the domesticated japonica, and its cultivation was always carried out in dry field systems during the Neolithic-Bronze Age. In the economic system of the Central Plain, mixed agriculture was a minor component during the Peiligang period, but became dominant beginning in the Yangshao Culture. The findings provide new references for clarifying the developmental process of mixed millet and rice farming in China, by contributing a diachronic and regional perspective to the Central Plain.
Footnotes
Acknowledgements
Funding
This work is supported by the National Basic Research Program of China (Grant No. 2015CB953803), the National Natural Science Foundation of China (Grant No. 41701233 and 41230104), the China Postdoctoral Science Foundation (Grant No. 2016M601124), the National Social Science Foundation of China (Grant No. 16CKG007), and the ‘Macroevolutionary Processes and Paleoenvironments of Major Historical Biota’ of the Chinese Academy of Sciences (Grant No. XDPB0503).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.