
Abstract
Lately, evidence for early-Holocene emerging sedentism has been suggested among foragers in Northern Europe. The core of this suggested sedentism lies in the increasing dependency on large-scale fishing and mass consumption of fish and a territorial behaviour associated with access to the best fishing locations. This territoriality might also be associated with increasing numbers of people settling and living in Northern Europe at this time. In this article, we review the evidence for forager sedentism and territoriality and relate it to large-scale fishing, during a time of global warming, in early-Holocene Scandinavia. We explore the requisites of using the archaeological record to study the long-term effect of intense fishing on some of the best-preserved Stone Age sites in the area of study. We suggest that the archaeological record can enable a discussion of how aquatic life varies corresponding to human exploitation and climate change. In addition, we discuss how these changes might be traceable through temporal fluctuations in species composition, within species size reduction/increases, temporal fish age changes and within species dietary changes. In the end, we suggest that the archaeological record holds one of the keys to predict future impact on life below the surface, by offering a long-term perspective on aquatic exploitation in a period of climate change. At the same time as we acknowledge the potential hidden in the archaeological record, we also raise the dire warning that this record might be rapidly disappearing, because of an accelerated deterioration of archaeological organic remains in areas previously known for their good preservation.
Keywords
Introduction
Climatic and environmental changes affect our world, not least the aquatic ecosystems where rapid and sometimes unpredictable changes appear. The anthropogenic-induced changes are apparent, however difficult to pinpoint (Hoegh-Guldberg and Bruno, 2010). In addition, oceans are being polluted by human waste products (e.g. plastic) and by industrial waste. Aquatic environments are also being depleted by the ever-expanding human demand for aquatic resources. This is taking a heavy toll on aquatic life and we are now starting to see the consequences. The depletion of aquatic environments is noted in decreased fish catches over the years (Thurstan et al., 2010) accompanied by a need to change fishing focus to other species, as the originally sought species become rare because of overexploitation (Myers and Worm, 2003). This is evident by a general decline in global aquatic species population numbers with more than 50% in the last 40 years (Tanzer et al., 2015). This has also resulted in an exponential growth in the number of species threatened by extinction, for example, around 25% of all rays and sharks are now critically endangered and threatened by extinction because of overfishing (Dulvy et al., 2014). Further implications, such as technological adaptation, become key to catching new species and we have seen the increasing use of, for example, bottom trawling, and other mass capture methods of fishing, peaking in the mid-1980s (Watson et al., 2006). These technological advances have often compensated for a gradually depleting aquatic life, complicating historical comparison with more recent fish catches. However, when fish catches are corrected for advances in technology and calculations are done on the same premises, it becomes apparent that aquatic life has been diminishing, at least in well-exploited areas, compare, for example, the dramatic decline in fish landings per unit of fishing power outside the UK during the last 118 years (Thurstan et al., 2010). On the same line, humans are travelling further into previously uncharted territories to fish, as the more local fishing grounds become depleted, disturbing the balance in the water in remote areas as well as local.
The social, ecological and political consequences of a depleted aquatic economy are hard to foresee. Even though we are beginning to see an increasing awareness of how to delimit and preserve special restriction areas where no fishing or other human exploitation is allowed, in order for the aquatic systems to recover, the results vary depending on how strictly the fish ban is enforced and on the physical properties of the no-go zones (Edgar et al., 2014). Furthermore, and despite the positive effect on the fish population, in Sweden, these ‘no-fishing zones’ cover less than 1% of the available sea, divided into 11 different areas, four in the Kattegat, four in the Baltic and three in Lake Vättern (Bergström et al., 2016: Figure 1). In addition, these Swedish areas constitute two-thirds of all the ‘no-fishing zones’ in Europe (Bergström et al., 2016).
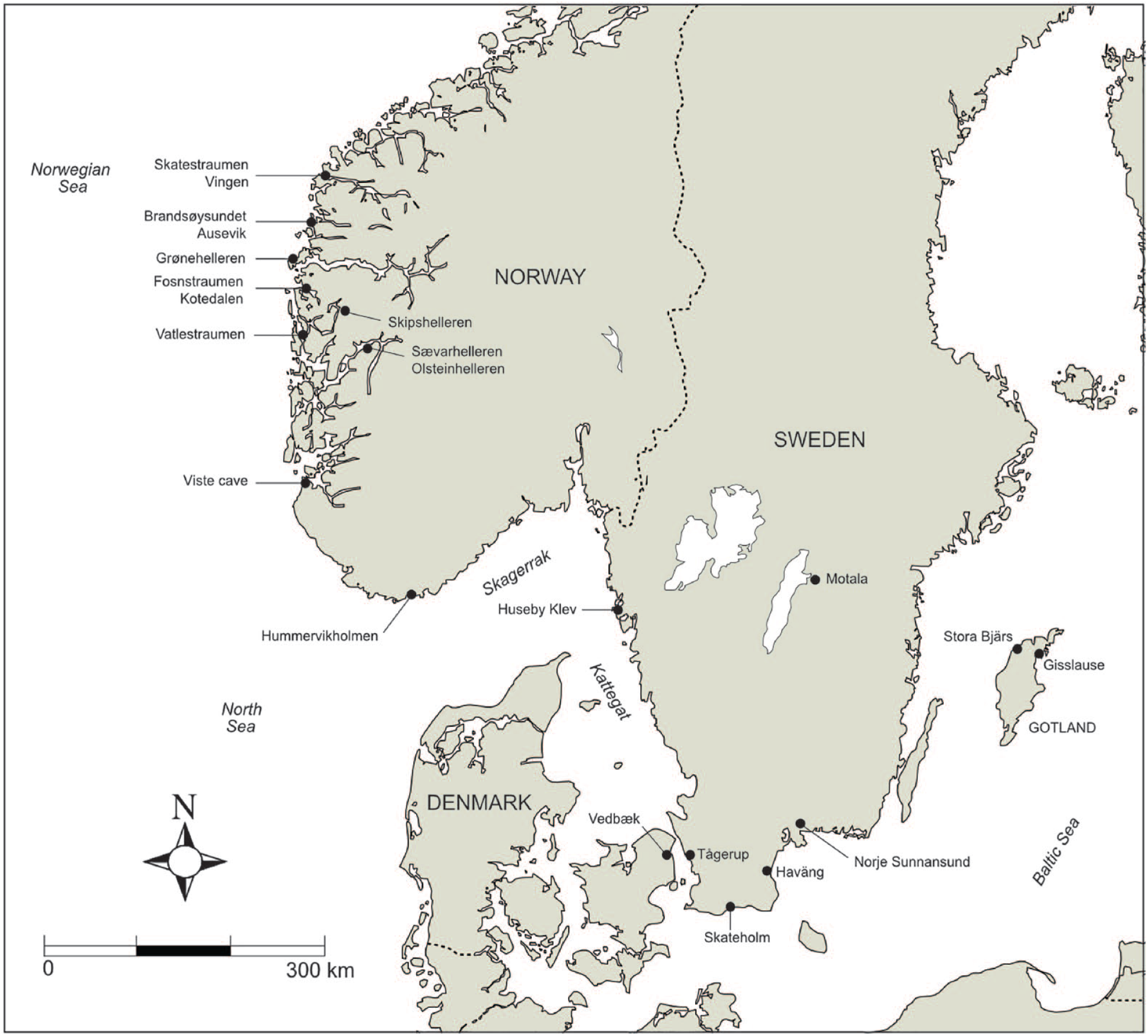
The area of study including the location of the sites referred to in this paper.
Considering the many anthropogenically induced problems in aquatic environments all over the world, it is evident that a successful aquatic future must rely upon scientifically trustworthy predictions. Today, aquatic ecologist and fisheries managers rely on historical catch records and ecological observations, spanning to around a century (often less), to create models for predictions of future aquatic life and abundances. These models are good for finding and using short-term fluctuations by naturally reoccurring phenomenon, for example, the fluctuation and correlation between El Niño and anchovy and sardine productivity in the eastern Pacific Ocean (Bakun and Broad, 2003; Chavez et al., 2003). However, the models are missing the long-term perspective in ecological variation, environmental changes and long-term effects of human aquatic exploitation.
This problem has been known for some decades and is often referred to as the shifting baseline syndrome (Pauly, 1995). Because of a lack of larger time depth, most modern ecological investigations fail to see the long-term perspective on both local and global aquatic life and are thereby fundamentally flawed. The failure to recognize the impact by indigenous people, prior to the industrial revolution, impairs the study of key species abundance and the long-term effect if they are heavily exploited for centuries (Jackson, 2001; Jackson et al., 2001). If global warming and its effect on aquatic life are added to the equation, we are in a predicament when modelling for future subsurface scenarios (if relying only on the historical record), since environmental changes are often highlighted as the most difficult aspect to account for when managing fisheries and establishing fishing quotas in different regions of today (Cury and Christensen, 2005; Hsieh et al., 2006; Pikitch et al., 2004).
If looking at today’s problem with depleting life in the oceans by using a longer time frame, it becomes apparent that the technological boom we have experienced in fishing methods over the last 100 years or so, for example, industrialized trawlers, sonar systems, steam and combustion engines, powered net drums, is a continuation of an ever-growing/changing adaptation to aquatic exploitation in general. Technological achievements in the past were likely driven by the same incitements as in modern times. Consequently, past humans were, just as today, trying to reduce the amount of invested labour while maximizing the yield and they were striving for stability by developing strategies to ensure that catches would not decline. Interestingly, this pattern has led to similar effects, as is observed in modern fisheries from all over the world, that is, a decline in one species (a high-trophic-level fish) is followed by an increased human focus of another (e.g. Baisre, 2000; Pauly and Palomares, 2005; Pinnegar et al., 2002; Stergiou and Koulouris, 2000; Valtýsson and Pauly, 2003). However, where the modern fisheries data stretch for decades, the archaeological record stretches for millennia, enabling the study of key species depletion and the long-term effect of their decline, as well as a deeper and more profound understanding of ecological bottlenecks and their effect on later species abundance and frequencies. In light of the time depth available from the archaeological record, it becomes clear that ecological modelling lacking this time depth becomes flawed because they fail to see the impact humans have had on the aquatic ecosystem for millennia prior to modern industrialized fishing (cf. Braje and Rick, 2011, and articles therein; Erlandson and Rick, 2010; Gifford-Gonzalez and Sunseri, 2009; Rick and Erlandson, 2008, and articles therein). Currently, these ecological models are thereby missing an important point, that is, they are based on false assumptions of an undisturbed baseline, aka a false ground zero. As a consequence, the ‘undisturbed and pristine’ environment believed to be ‘natural’ is, in fact, an aquatic ecosystem which has already experienced collapses, key species overexploitations and extinctions; thereby, there are no existing pristine ecosystems and most biotopes on earth have been under stress from humans for millennia.
The dangers of believing that a certain ecosystem is ‘natural’ when it, in fact, has been challenged for thousands of years is understandably hazardous as it becomes difficult to tell when and how close a system collapse is or when the allé-effect threshold (Allee et al., 1949; Dennis, 1989) is reached, that is, when further exploitation will push the species over the edge of extinction. However, this need not be the case and, unknowingly to most ecologists dealing with these types of dilemmas, the archaeological record is a good source of information to gain a long-term perspective on the fluctuations that have occurred during (in Scandinavia) the last 14,000 years or so. In this largely unused record lies the effect of climate change and intense human aquatic exploitation. In other words, aquatic life has changed and adapted over millennia of temperature changes since the last Ice Age, and subsurface ecosystems have undergone a continuous change through the effects of intense human aquatic exploitation.
What does the archaeological record have to offer?
Human impacts on aquatic environments are currently at an all-time high. We are exploiting both close and remote areas, changing the composition in all water bodies simultaneously, which makes repopulation of collapsed areas much more difficult now, compared with how it has been before industrialized fishing and modern fishing technology. The recovery time after a fish stock collapse is considerably longer than originally suspected (Hutchings, 2000), implying that if continuous exploitation occurs after a collapse or if the collapse is situated in the contexts of changing climate (e.g. through global warming), its effect might be altogether irreversible. The overexploitation of a fish population, under these circumstances, might cause severe changes in the aquatic ecological composition and cause local depletion of key species and disruptions in the ecosystem (Beverton, 1990). Furthermore, industrial pollution and plastic waste littering the oceans are affecting the aquatic ecology without any ancient or historical analogies to rely on for comparison. Considering this, is it worth the time and effort taken to secure a long-term record, given all the dissimilarities and unique prerequisites connected to modern fisheries, or should we resign and hope that the short-term record is enough to accurately model the future of different aquatic systems?
To enable any form of comparable analogies for future long-term perspectives, it is important that the archaeological material is carefully selected. First, to simulate the global impact on fish stocks of today, the archaeological material has to represent areas that have been under intense human exploitation for centuries or millennia, without pauses that could have allowed the local aquatic systems to recover. Second, global warming and rapidly increasing water temperature also play a significant role in aquatic life abundancies. Thereby, to account for the ongoing global warming of today (Brooks et al., 2018; Goudie, 2018; McCarthy et al., 2001; Root et al., 2003), the studies on archaeological material have to derive from a period of similar temperature increase and the archaeological assemblages have to be large enough and cover enough time and space to be representative. Third, the effects of modern pollution have to be considered and its negative effect on different species and habitats has to be accounted for and the data have to be applied to the archaeological models before comparisons can be made. Fourth, the recovery of the archaeological material has to be (or have been) such that biases can be ruled out, or accounted for. Fifth, the organic preservation on the archaeological sites has to be very good to enable the preservation of small and fragile fish bones. Sixth, there should be a reasonable control with the shoreline displacement in the relevant study area, since there are often major differences in the use of aquatic resources between shore-bound and interior archaeological sites. Finally, investigations of archaeological material should aim to start as far back in time as possible, to mitigate the effects of earlier human exploitation disturbing the ‘baseline’.
While all the prerequisites discussed above cannot be met without further studies, the Scandinavian early-Holocene archaeological record fits most of these criteria and would, consequently, be perfect to use as a point of departure. We will exemplify and highlight this below, using southern Sweden/Denmark and western Norway as case studies (Figure 1). By discussing the prerequisites for heavy fish exploitation among Scandinavian early-Holocene societies, and how they developed into sedentary and territorial fishing communities, we wish to show that an understanding of ancient life and subsistence strategies is essential for discussing their impact on the environment. By extension, we will connect the material from some of the key sites and envision to what extent this material can be used to study the long-term effects of aquatic exploitation during a period of gradual global warming.
Early-Holocene fishing in Scandinavia
Fish have long been considered as a major dietary source in late Mesolithic (around 7500–6000 cal. BP) southern Scandinavian contexts (Rowley-Conwy, 1983), often because of the discovery of well-preserved shell middens and cultural layers with abundant fish bone material (Enghoff, 1987, 1989, 1991, 1994, 2011; Ritchie, 2010). Until recently, little data on fish exploitation have been available from the first millennia of the early Holocene, whereby fishing has been largely invisible in discussions of diet and subsistence (Boethius, 2018a). However, in the last few years, and from a Scandinavian perspective, assumptions regarding limited importance of fish in early-Holocene subsistence have been questioned (Boethius, 2018a; Hansson et al., 2019; Robson and Ritchie, 2019). Several new studies have shown an increasingly strong evidence for mass exploitation of fish during the early Holocene, not least through the discovery of the early-Holocene site Norje Sunnansund (dated to around 9600–8600 cal. BP), located on the southeast coast of Sweden, in which large amounts of fish bones were recovered (Boethius, 2016, 2017, 2018c; Kjällquist et al., 2016). The reanalysis of the bone material from the early to middle Mesolithic site Huseby Klev on the Swedish west coast (Boethius, 2018b) and the circumstantial evidence from the island of Gotland in the Baltic Sea (Boethius et al., 2017) provide further indications of the importance of aquatic resources. Furthermore, the discovery of late Maglemose large-scale stationary fish traps at the now submerged site Haväng on the southeast coast of Sweden (Groß et al., 2018; Hansson et al., 2018; Nilsson et al., 2018), and the even older Antrea Korpilahti fishing net from Finnish Karelia (Miettinen et al., 2008; Pälsi, 1920), highlights the importance of fish (Figure 2).

Photos from some of the Swedish sites mentioned in the text: (a) Two-step water sieving at Huseby Klev. Photo: Bengt Nordqvist. (b) The excavation of Norje Sunnansund. © Blekinge Museum. (c) Fish trap at Haväng. Photo: Arne Sjöström. (d) The fermentation pit at Norje Sunnansund. © Blekinge Museum. (e) and (f) Water sieving and excavating at the site Gisslause on Gotland. Photo: Jan Apel.
In western Norway, aquatic resources have long been assumed to have played an important role, not least because the vast majority of Mesolithic sites are located very close to the contemporary shorelines. Direct evidence for fishing and consumption of fish is generally lacking in this region, because of the bad preservation conditions for bone material. However, a few sites have excellent data from c. 9000 cal. BP onwards. Fishing equipment and large amounts of fish bones are, for example, present at the recently excavated cave/rockshelter site Sævarhelleren dated to c. 9000–8000 cal. BP (Bergsvik and David, 2015; Bergsvik et al., 2016; Ritchie et al., 2016). Data from the contemporary Viste cave also indicate fishing (Mikkelsen, 1979; Schulting et al., 2016). The open-air site Kotedalen also has fishhooks and significant amounts of (burnt) fish bones from c. 8500–8000 cal. BP (Bergsvik, 2001; Hufthammer, 1992; Olsen, 1992), and the later Mesolithic rockshelter sites Skipshelleren and Olsteinhelleren (c. 7600–6800 cal. BP) also have fishing equipment and abundant fish bone material (Bergsvik et al., 2016; Faltinsen, 2018; Ritchie et al., 2016; Figure 3). It appears that the main fishing methods were by fishhooks and barbed points. Except for large possible net-sinkers of stone and a few possible netting needles of bone (Bergsvik and Ritchie, in press), evidence for stationary fishing has yet to be uncovered, and structures for storage are also lacking. However, this may be because of the poor conditions for the preservation of organic materials at open-air sites in this region.

Photos from some of the sites in western Norway mentioned in the text: (a) The site Kotedalen during excavation in 1987. The tidal current channel Fosnstraumen is in front of the site. Photo: David N. Simpson. (b) The rockshelter Sævarhelleren during excavation in 2005. Photo: Knut A. Bergsvik. (c) Excavated floor at the Mesolithic site area at Kotedalen. Photo: Knut A. Bergsvik. (d) The rock-art site Vingen. Photo: Knut A. Bergsvik. (e) Mesolithic shells and bones during sieving at the site Sævarhelleren. Photo: Knut A. Bergsvik. (f) The rockshelter Skipshelleren during excavations in 1931. Photo: Johs. Bøe. (g) The tidal current channel Skatestraumen. Photo: Knut A. Bergsvik. (h) The rockshelter Olsteinhelleren during excavation in 2006. Photo: Knut A. Bergsvik.
The above examples suggest that fish did make up a large part of the diet from at least the early Holocene in Scandinavia, and the Swedish evidence also shows that the fishing and storing techniques were highly effective, elaborate and stationary. It is somewhat more uncertain whether the fishing only with hook and line matches that level of efficiency (e.g. Kelly, 2013: 58). Nevertheless, when investigating the stable isotope values from the collagen of available human bone remains from early-Holocene southern Scandinavia (82 individuals), they indicate that fish and aquatic resources dominated the protein intake of these individuals (Boethius and Ahlström, 2018). A temporal decrease in human isotopic niche width is also detectable from the early to the middle Mesolithic period, which suggests a general homogenization of subsistence strategies, that is, a consecutive increasing fish dependency in southern Scandinavia (Boethius and Ahlström, 2018). In western Norway, only a few Mesolithic human bone remains have so far been dated and analysed. The stable isotope analyses from these skeletons equally indicate a dependency on aquatic resources (Schulting et al., 2016; Skar et al., 2016).
These new pieces of evidence, sprung from new methods of excavating, sampling and analysing the early-Holocene archaeological record, highlight the importance of aquatic resources, regardless of whether its exploitation is connected to marine, riverine or lacustrine ecosystems. Thus, aquatic life, from both fresh and saline waters, can be considered of major dietary importance for humans in Scandinavia from, at least, the early Holocene. A perceived importance of aquatic resources, in turn, affects our current view of these foragers and widens our understanding of these societies and the impact they did on their surroundings.
Sedentism among foraging aquatic economies
Human societies can, under certain circumstances, affect the local environment if they have a large enough demand for a certain resource. However, to do so, the humans must be numerous enough and stationary enough not to give the resource in question time to recover. When studying hunter–fisher–gatherer societies, they can be seen as either mobile or stationary. If applying optimal foraging theory to the ethnographic data on the mobility of a forager group, it indicates that when practising a high mobility rate, people are moving between different resource patches, and when it becomes more difficult to find resources, you move on to a better area (Kelly, 2013: 85–87). Thus, if discussing animal exploitation connected to high human mobility, the number of available animals in a specific area will not be diminished to unrecoverable levels. People will move from the area long before it is completely drained, that is, when the amount of labour invested in searching for, killing, butchering and bringing the animal back to camp is more ‘costly’ than moving to a new area where animals are easier to find.
In 1980, Binford divided hunter–fisher–gatherer societies into two different types, those practising logistical and those practising residential mobility (Binford, 1980). Logistical mobility was defined as humans who rarely moved their homes but moved logistically through the landscape to gather/hunt resources, which were brought back to the settlement. Residential mobility is practised when foragers move their entire homes when it becomes difficult to gather or hunt food in a particular area. Consequently, when people are practising logistical mobility, they have a more or less sedentary home base and are therefore able to exploit the local resources during extended periods.
Up until a few decades ago, foragers were largely thought to primarily rely on residential mobility strategies for their subsistence. However, in the last few decades, arguments have been put forward that supports the concept that sedentism can evolve in forager economies if the right requirements are met, for example, temporal and spatial resource abundance, stability, and variability. Sedentism may, in turn, imply population pressure, and sometimes it leads to developed storage systems, access to mass harvesting technologies, labour control and even tribal warfare (Ames, 1981; Burley, 1980; Fitzhugh, 2003; Kelly, 2013; Rowley-Conwy and Zvelebil, 1989; Schalk, 1981; Testart, 1982). These types of ideas have enabled the possibility to view foraging societies as a continuum between groups that are egalitarian and mobile on the one end, and hierarchal, complex and sedentary on the other (Murdock, 1968; Price and Brown, 1985; Rowley-Conwy, 1983). Foragers characterized by the last attributes are often connected to the exploitation of aquatic resources (Ames, 1994).
While sedentism among foragers is primarily known from ethnographic sources, indications of sedentism and limited residential mobility have recently been suggested from the early Mesolithic period and onwards. The dietary information obtained from stable carbon and nitrogen isotope values (δ13C and δ15N) from available southern Scandinavian human remains show, for example, no overlap in the early Mesolithic period and very limited overlap in the middle Mesolithic period (Figure 4), which indicates limited coast to inland mobility (Boethius and Ahlström, 2018). Limited residential mobility is also suggested at, for example, Norje Sunnansund, where the hunting strategies and animal exploitation patterns seem to have been adapted to a sedentary lifestyle (Boethius, 2017). Limited residential mobility can also be discussed in terms of increasing reliance on stationary fish traps (Nilsson et al., 2018) and on very large amounts of fish caught at Norje Sunnansund (Boethius, 2018c) in connection to evidence that people were also able to ferment fish as a means of conservation to facilitate long-term and large-scale storing (Boethius, 2016). Sedentism can also be seen at the coast of western Norway from at least during the late Mesolithic around 8200 cal. BP. This is indicated by the increased sizes of the sites and cultural layers, a broad-spectrum economy, faunal data showing several seasons of continuous occupation and evidence of reoccupation of the same sites (Bergsvik, 2001, 2002; Hufthammer, 1992; Olsen, 1992; Warren, 1994). Aside from arguments on population packing, sedentism in this area also generally makes sense, because residential mobility in the north–south direction along the coast would not provide access to any new resources (Warren, 1994: 86–88). Instead of residential mobility along the coast, then, the populations seem to have practised seasonal movements – within presumed territories – eastwards into the fjord systems (Bergsvik et al., 2016).
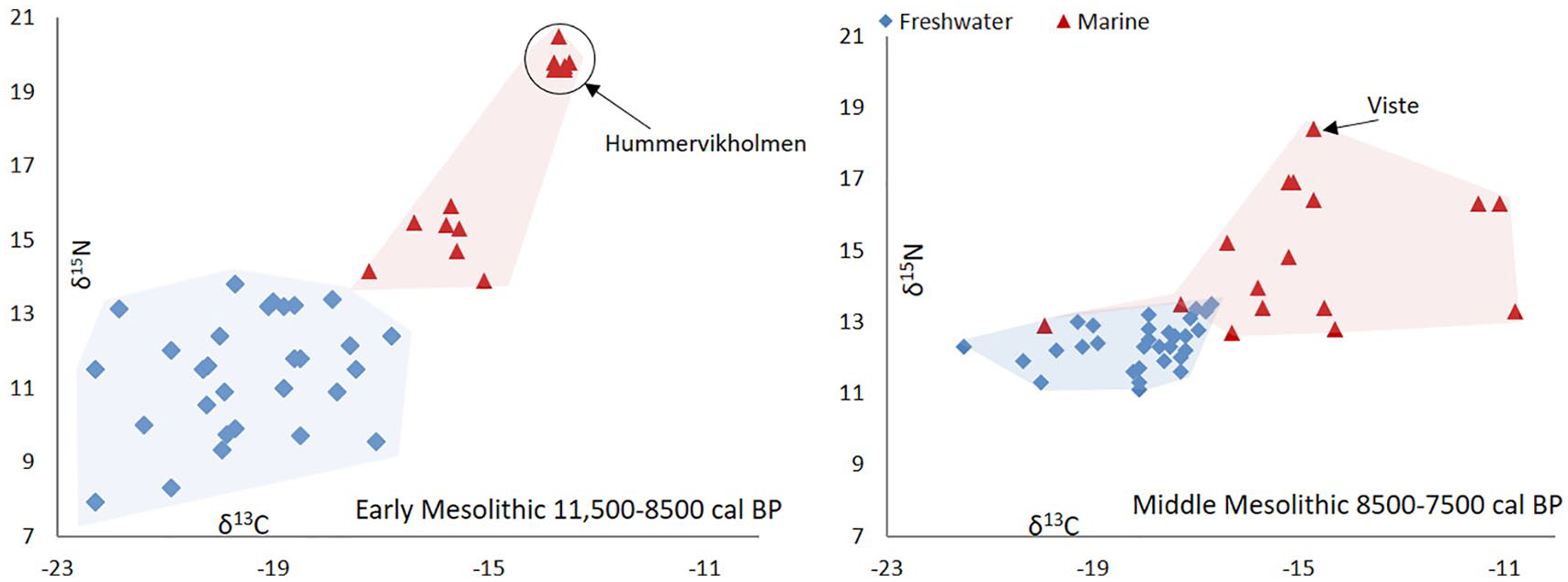
Bulk collagen stable isotope data from all available early and middle Mesolithic humans from Scandinavia showing no isotopic overlap during the early Mesolithic and little overlap during the middle Mesolithic, which suggests limited coast–inland mobility and an indication of a territorially divided landscape during the early Holocene. Data and original publication in Boethius and Ahlström (2018), here illustrated with the addition of the human isotope values from Hummervikholmen (Skar et al., 2016) and Viste (Schulting et al., 2016). Early and middle Mesolithic periods are here defined according to south Scandinavian chronology (Sørensen, 2012).
Territoriality among ancient foragers – A prerequisite for a long-term fishing record?
A general increase in fish dependency and expanding sedentism, as suggested above, can, under certain circumstances, be related to how territoriality emerges or expands among foragers (Boethius, 2018a; Groß et al., 2018) and it can even be self-generating and act to intensify fishing even further. This may happen if, for example, a foraging society with a high mobility rate live in an area where other groups of people start to settle and shift their economy more towards aquatic resources, as is basically a requirement for sedentary life among foragers on high latitudes when population pressure increases (Binford, 2001: 197, 366). Then, the mobile groups will be forced to avoid the more settled areas, which will be more densely populated by the sedentary fish reliant foragers (as fish dependency can also often be considered a density-dependent response; cf. Marlowe, 2005; or Binford, 2001), or risk physical confrontation with the more numerous settled groups. This, over time, will result in increasing mobility ‘costs’ as the most favourable spots in the landscape will be out of reach, resulting in larger detours through the landscape to reach other environmental ‘hot spots’ (Kelly, 2013). Thus, mobility becomes more expensive and under these circumstances, (1) where there is variation in resource accessibility (e.g. when important fishing sites, where it is possible to extract large amounts of fish, appear at specific but limited number of locations) and (2) high degree of predictability (e.g. when it is possible to predict when resources will be available at these important fishing sites). Territoriality can thereby be linked to decreasing residential mobility among all groups in an area and increasing fish dependency (Boethius, 2018a). Evidence of territoriality can also be suggested both among early-Holocene societies in southern Scandinavia and on the Norwegian west coast.
Territoriality and ‘hot spots’ along the Norwegian west coast
Because of the shallow waters in the skerry landscapes, and the strong water movements caused by the Atlantic current, a series of local areas along the Norwegian coast have very high marine productivity (Bjerck, 2007, 2008). These local ‘hot spots’ are mainly tidal current channels and are found in straits between islands at the protected parts of the coast. The shores along several such straits or channels have been test-pit surveyed and excavated, such as Fosnstraumen (Bergsvik, 2001; Hufthammer, 1992; Olsen, 1992), Vatlestraumen (Kristoffersen, 1995), Brandsøysundet (Olsen and Alsaker, 1984) and Skatestraumen (Bergsvik, 2002). Comparative studies show that there are marked concentrations of large sedentary Mesolithic sites in these areas compared with areas outside of the channels, which have smaller and fewer sites (Bergsvik, 2001). These ‘hot spots’ were likely nodes in a territorial organization. Another indicator of territorialism in western Norway is the distribution pattern of quarried lithic raw materials (basaltic rocks, siltstones and cataclasites). The distribution of these raw materials shows coincidental distribution falloffs, indicating restrictions on residential mobility (but not task group mobility) across territorial boundaries along the coast (Bergsvik and Olsen, 2003; Nyland, 2015; Olsen and Alsaker, 1984; Skjelstad, 2003). A further indication of territoriality in western Norway is the presence of large rock-art sites at Ausevik (c. 300 figures) and Vingen (c. 2200 figures), dated to between 7500 and 6000 cal. BP, which are situated relatively close to the ‘hot spots’ Skatestraumen and Brandsøysundet, respectively (Hjelle and Lødøen, 2017; Lødøen, 2014). These rock-art sites may have been important ritual arenas in which local territorial rights and obligations were established and maintained.
Territoriality through stationary technology
In southern Scandinavia, the earliest known evidence of fermentation is detected on the early-Holocene site Norje Sunnansund site in southeastern Sweden (Boethius, 2016). From a territorial perspective, evidence of fermentation is important as it is a type of delayed-return subsistence strategy, a food storage facility designed to store large amounts of food during extended periods. The effect and benefits of large storage systems are important as they are impractical and difficult to move over large areas (Rowley-Conwy and Zvelebil, 1989), meaning that if you store large amounts of food, you invest it in that precise area and thus need to make sure nobody else takes advantage of your food supply. Large food storages thereby act as a societal anchor, fettering a group to the food storage itself. Stationary fish weir and traps are similar to large storages as they are also an indication of a delayed-return subsistence strategy. By investing large amounts of both time and labour into creating these fish traps, you, much like with large storages, also anchor yourself to these contraptions. Thereby, because the traps are stationary and placed to optimally exploit a certain part of an aquatic system, they are also tying the group to the area. Interestingly, evidence of fish traps designed to capture large amounts of fish exists in Scandinavia from the early Holocene and onwards, both from the east and west coast of southern Scandinavia (Hadevik et al., 2008; Hansson et al., 2018; Karsten and Knarrström, 2003; Pedersen, 1995). Thus, both large food storages and stationary mass capture fishing devices can be seen as a form of early territorialisation (Nilsson et al., 2018).
Territoriality through human remains
Another interesting observation related to early-Holocene territoriality is the level of interpersonal violence among the foragers of southern Scandinavia. For example, among the foragers in Motala Kanaljorden in south-central Sweden, 34 human bones from a minimum of 10 individuals were recovered on the excavation. Seven of these individuals displayed antemortem injuries to the head, on some occasions single blunt force trauma and, on other occasions, multiple traumas of both healed and unhealed fractures (Gummesson et al., 2018). Violence among early-Holocene foragers from Scandinavia is not limited to Motala but can be seen on a number of occasions, for example, through the skull trauma and the slotted bone point suggested to have been shot into the pelvic area of the Stora Bjärs man on Gotland (Arwidsson, 1979; Larsson, 1982), and from Denmark several Mesolithic crania have trauma caused by blows of axes and blunt instruments (Bennike, 1997).
In addition to violence-related trauma among early northern foragers, there are also indications of graves having been used as territorial markers. From the early Mesolithic period, suggestions of plausible excarnations located on visible spots in the landscape have been brought forward (Boethius, 2018a). During the middle and late Mesolithic period, the burial customs include regular incarnation, that is, graves and cemeteries on visible locations, compare, for example, the location of the graves from middle Mesolithic Tågerup (Ahlström, 2001; Karsten and Knarrström, 2003; Kjällquist, 2001), possibly also the prominent Viste cave (Fürst, 1909) and the site Hummervikholmen (Skar et al., 2016), and late Mesolithic Skateholm (Larsson, 1988a, 1988b, 1993). The same may have been the case with the late Mesolithic Vedbæk (Albrethsen and Brinch Petersen, 1976; Brinch Petersen, 2015), Motala Strandvägen (Gummesson and Molin, 2016) and Grønehelleren (Jansen, 1972). To conclude territorial indication based on human skeletal remains, it is appropriate to return to the Motala Kanaljorden site where the human skulls found there not only displayed large amounts of trauma (Gummesson et al., 2018), but also showed that the human skulls had been impaled and put on display in the bog, with parts of the stakes still protruding from the foramen magnum on some of the skulls (Gummesson et al., 2018; Hallgren, 2011; Hallgren and Fornander, 2016). Suggestions have also been made that this was likely not a single occurrence and similar patterns of skull fragments in the proximity of a wooden stake have been noted in, for example, Norje Sunnansund (Boethius, 2018a). Furthermore, human scattered remains occur on around 50–70% of the known Scandinavian Mesolithic settlements, depending on how calculations are made and what type of settlements are included (Boethius, 2018c). Most of these scattered remains come from parts of human skulls, whereby suggestions of a Mesolithic skull cult have even been made (Schulting, 2015). Combined, this suggests that the Motala impaled skulls might not be a unique occurrence among Mesolithic societies; its uniqueness instead lays in the preservation and recovery of the skulls. Consequently, the scattered human remains on Mesolithic settlements create a territorial link to the sites, especially if it can be suggested that many of the remains represent impaled skulls put on display.
Additional territoriality signals
When the stable isotope values from C and N were analysed from the 82 early-Holocene individuals from southern Scandinavia mentioned above, and if also including the individuals from Hummervikholmen and the Viste cave, the results not only indicate that aquatic resources were the main protein source to diet, it also showed that resources from freshwater and marine carbon pools had been consumed in different areas and that their isotope values were separated from each other (Figure 4). This suggested a limited mobility between coast and inland. If there had been continuous mobility between coast and inland, the dietary signals would show overlaps as the humans would have consumed a non-local diet during different parts of the year, supporting an idea of territorial diet patterns (Boethius and Ahlström, 2018).
Indeed, this type of territorial dietary pattern is further supported by indications of selective hunting strategies aimed to optimize the long-term gain of secondary products from terrestrial mammals. For example, at Norje Sunnansund, it appears as if red deer (Cervus elaphus) were systematically hunted when they had reached full body size but before they were able to breed, thus juveniles and prime animals were spared presumably to ensure the continued presence in the area (Boethius, 2017). This type of hunting strategies only makes sense for sedentary and territorial hunter–fisher–gatherers (else they would move on when the population size of sought animals decreased) who relied on other food sources for sustenance (i.e. fish), thus needing a long-term supply of raw material, for example, bones, hides and sinews, but who primarily relied on other resources for food (Boethius, 2017, 2018a: 125).
Population size in Scandinavia
It is challenging to investigate and calculate population densities in archaeological contexts and the further back in time you go the more complicated it gets. Mesolithic demography and population densities are very likely among the most difficult to account for in Scandinavian archaeology, because of the problems with finding Mesolithic sites, especially with organic preservation as they are often inundated, located in inaccessible wetlands or on the bottom of the sea. Also, the Mesolithic remains on land can have potentially been disturbed by later societies living in the same area, making them harder to find and interpret. It is generally argued that during the early Mesolithic, the foraging societies in Scandinavia were more mobile and that the area was consequently less densely populated (Bjerck, 2008; Blankholm, 2008). Related to the debate on Mesolithic demography and with the suggestions of developing sedentism, it has been possible to argue that at least some areas in Scandinavia were rather densely populated during the later parts of the Mesolithic (Solheim and Persson, 2018; Tallavaara and Pesonen, 2018). But this has even been suggested on the early Mesolithic west coast of Sweden (central part of Bohuslän) which might have contained as much as 10,000 late Hensbacka sites (Schmitt et al., 2006). Based on these calculations, large population densities might be suggested during the transition from the late Pleistocene to early Holocene on the Swedish west coast. A large population density could also be plausible at Norje Sunnansund, where calculations of taphonomic loss indicate catches of fish large enough to support a large population over a long period (Boethius, 2018c). Furthermore, these types of large fish catches were not unique for Norje Sunnansund, for example, as illustrated by the contemporaneous fish traps found at Haväng (Nilsson et al., 2018), which were large and located in areas where they could provide large catches of fish. Because of the fishing capacity of the fishing weirs at Haväng, and its location about 60 km by boat from contemporaneous Norje Sunnansund, and because most coastal sites from the early Mesolithic period are now submerged through transgressions following the last Ice Age (whereby evidence of them is difficult to obtain), the results from Haväng and Norje Sunnansund could imply a general population intensification on the east coast of Sweden.
Large population densities in early-Holocene Scandinavia may also be implied by evidence of recent ancient DNA (aDNA) analyses, indicating a greater genetic diversity among Scandinavian Mesolithic foragers, compared with contemporaneous foragers from more southern regions of Europe (Günther et al., 2018). Scandinavian hunter–fisher–gatherer genetic diversity is seen as an indication of genetic mixing of two different populations migrating from separate directions, via a north-eastern coastal route (Eastern hunter–fisher–gatherers) and from the European continent via Denmark (Western hunter–fisher–gatherers), into Scandinavia to meet and mix (Günther et al., 2018; Kashuba et al., 2019; Mittnik et al., 2018).
Consider that close to optimal aquatic conditions existed in Scandinavia at the time, that is, an ecological bonanza in the ocean along the Scandinavian west coast during initial Holocene (Boethius, 2018b) and unlimited access to large productive freshwater lakes connected via rivers and outlets to larger water bodies (Boethius, 2018a; Boethius et al., 2017). Given these conditions, a greater human genetic diversity might not only be an indication of the genetic mixture of two hunter–fisher–gatherer populations (Günther et al., 2018), it might also indicate high population densities (Boethius, 2018a: 111) and the development of social territories (Olsen and Alsaker, 1984). This in turn could imply that the observed migration, seen by both genetics (Günther et al., 2018) and stone/bone tool technology (Bergsvik and David, 2015; Damlien, 2016; Sørensen et al., 2013), was driven by an aquatic abundance, which provided optimal conditions for staying here, resulting in higher population densities in Scandinavia if compared with more southern regions of Europe during the early Holocene.
Archaeology and aquatic ecology
Sedentary cultures with an increasing population are prone to affect the ecosystem in the area around the settlements. On land, this is hinted at by the lack of/temporal diminishing numbers of the largest ungulates with the slowest reproduction rates (aurochs and elk) in coastal areas (Magnell, 2017: Figures 7.2–7.3), that is, where the largest sedentary settlements were presumably located. A large dietary dependency on aquatic resources can also have affected the aquatic systems, because of the large amounts of resources being extracted. This type of plausible overexploitation is possibly seen on the Huseby Klev site on the west coast of Sweden. On Huseby Klev, three different settlements were located on the same area dating to 10,300–9600 cal. BP, 9600–8700 cal. BP and 8000–7700 cal. BP. Zooarchaeological analysis on the bone material from the three different settlements suggests a temporal diminishing of aquatic mammals (whales and seals), corresponding with a temporal increase in fish (Boethius, 2018b) (Figure 5). The shift in dietary focus coincide with landscape changes and even though the sites are today in the same area, the first occupation was located in a narrow strait where it would have been optimal to hunt small whales (Boethius, 2018b). This type of location is also characteristic for the earliest archaeological sites on the Swedish west coast (Kindgren, 1995). The sites from the two later phases on Huseby Klev are located in a more sheltered bay, which is also in accordance with the general temporal trend in the area (Boethius, 2018b; Kindgren, 1995).
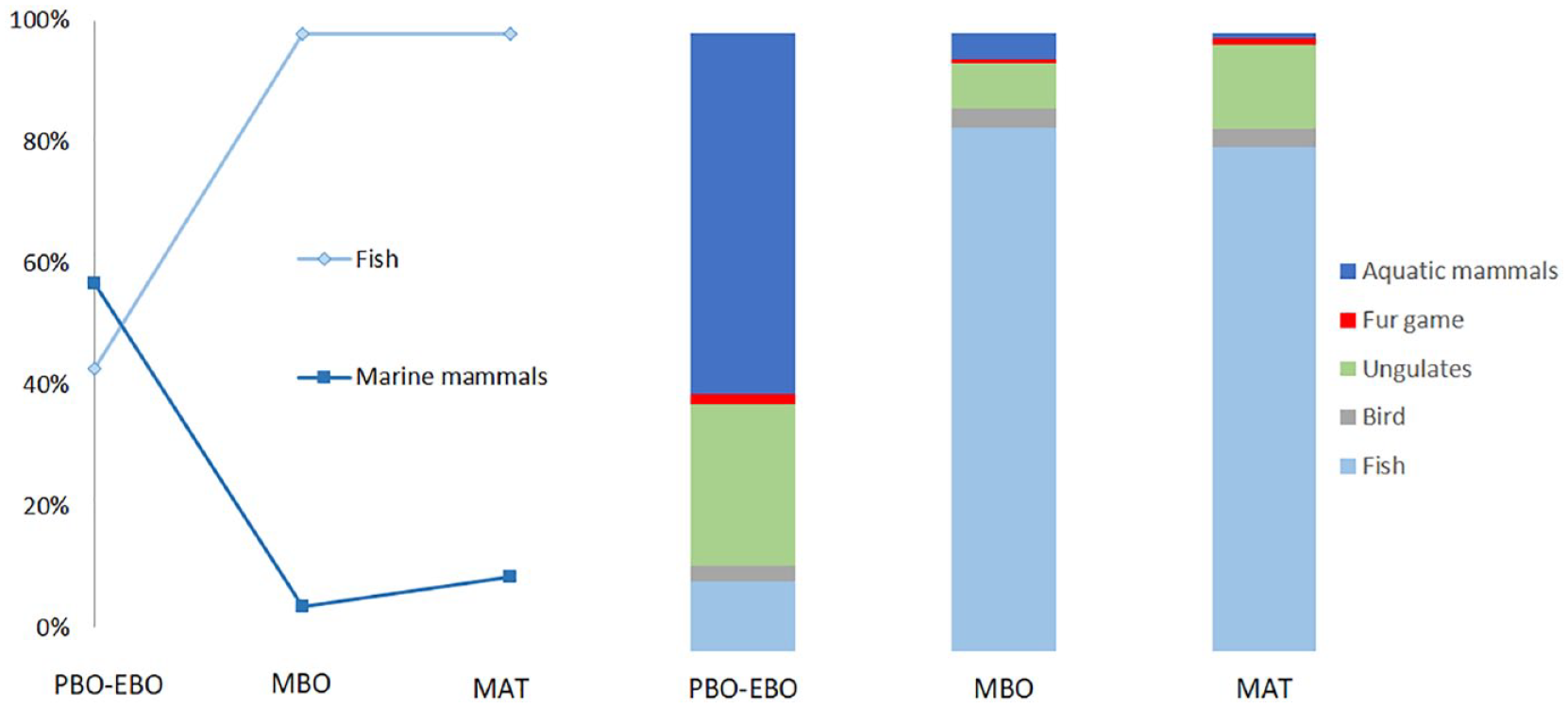
Frequency of fish and marine mammals between the different phases at Huseby Klev. Left: for marine mammals, based on a comparison of NISP (number of identified specimens) between marine mammal bones and the total amount of mammal bones. For fish, the comparison is based on ENISP (estimated number of identified specimens) in comparison with NISP from all mammals and birds. NISP: mammals: PBO–EBO (preboreal–early Boreal) = 364; MBO (mid-Boreal) = 149; MAT (mid-Atlantic) = 169. NISP birds: PBO–EBO = 77; MBO = 50; MAT = 15. ENISP fish: PBO–EBO = 327; MBO = 8782; MAT = 7939. See Boethius (2018a) for further details. Right: Animal-type distribution based on bone weight from the different phases at Huseby Klev. N: PBO–EBO = 4158.3 g; MBO = 3971.3; MAT = 1552.4 g.
Considerations must be taken because of variation in preservation conditions, dissimilar deposition patterns, minor variations in recovery methods (though all three phases were excavated within the same excavation) and possible disturbances to the cultural layers between the different occupations at the site. Nevertheless, the observed pattern might be one of the oldest indications of humanly (in combination with increasing aquatic temperatures) induced changes to life in aquatic environments. Huseby Klev is thereby one of the sites where it would be interesting to further analyse the fishbone assemblage to see whether changes in both size and fish species composition occurred over time, especially since the three phases likely stem from different settlement types, which, if considering optimal foraging theories, might be said to reflect the availability of different resources. The initial occupation phase could thereby represent a seasonal hunting site where the inhabitants gathered during limited periods of the year to mainly hunt marine mammals, while later occupation phases likely represent more sedentary communities relying on fish as their main diet source. The shift in diet could, in this aspect, be stemming from a decline in the originally hunted small whales and seals and would, in this case, be an early indication of the ‘fishing down the food chain’ syndrome.
Similar changes may also have taken place in western Norway, albeit possibly with a slight temporal delay suggesting that the dietary changes occurred somewhat later further to the north. Stable isotope values (δ13C and δ15N) from the human skeletons (C14-dated to between c. 10,000 and 9000 cal. BP) from the site Hummervikholmen show that more than 80% of the dietary protein was from marine resources, and from a very high trophic level. The results are interpreted to indicate that these individuals were specialized seal (Phocidae) hunters (Skar et al., 2016). However, since no animal bones related to the human finds have been recovered, the very high human δ15N-values at Hummerviksholmen (ranging from 19.6% to 20.5% (Skar et al., 2016)), may also imply, similar to the oldest phase from Huseby Klev (Boethius, 2018b), a heavy exploitation of small whales (Delphinidae) along with the exploitation of seals. The skeleton from the Viste cave is dated to c. 8200–8000 cal. BP, and the isotopic analyses show values that indicate a high trophic marine diet. However, with around 1–1.5% lower δ15N values than the individuals from Hummervikholmen, Schulting et al. (2016) argue that large cod (Gadus morhua) may also have been consumed by this individual. When looking at the osteological material from phase 2 and 3 at the sites Kotedalen (c. 8600–8000 cal. BP) and Sævarhelleren (c. 9000–8000 cal. BP), marine mammals, particularly seals, are well represented in the faunal data, but these assemblages are nevertheless dominated by fish bones – first of all gadids (Gadidae) (Bergsvik et al., 2016; Hufthammer, 1992). This may indicate that there was a development from a focus on marine mammals until around 9000 cal. BP, and then a change took place, where fishing of cod gained importance at the expense of seal hunting. One possible cause of this change may have been overexploitation, that is, a reduction in the stocks of seals along the outer coasts during these earliest millennia of human occupation in western Norway, which increasingly led people into other niches of food procurement.
There are also changes during the period c. 9000–6800 cal. BP, particularly considering fish species caught within the gadid family. This development is observed at the rockshelters Sævarhelleren and Olsteinhelleren, which are situated close together by the Hardanger fjord. In the faunal assemblages at the oldest site Sævarhelleren, there is a dominance of cod (Gadus morhua), whereas other gadids have lesser portions. In addition, wrasses (Labridae) make up a reasonable share. In the later Olsteinhelleren, a marked change has occurred, where saithe (Pollachius virens) has taken over the dominating position, whereas the consumption of cod is significantly reduced (Figures 6 and 7). The fishing of wrasses is also reduced. At both sites, 2- to 3-year-old specimens dominate for all gadids (Ritchie et al., 2016). It is somewhat uncertain what caused this change. The conditions for preservation were similar at the two sites, and they have been fishing in the same waters. One possible factor might be changing environmental conditions, leading to changes in species composition in the sea. During the relevant period, the sea level in front of the rockshelters decrease from c. 27 to 17 m. However, despite these changes, it is unreasonable that they had any impact, considering that the Hardanger fjord is 860 m deep near the rockshelters. Changes in the salinity of the waters might have played a role. However, these were also marginal, and it is also uncertain whether there were differences in sensitivity to salinity between different types of gadids. Another possible factor is human behavioural change. However, both sites seem to have been mainly used during the summer as field camps during logistical forays from the coastal settlements – although Olsteinhelleren for longer periods at the time. The groups using Olsteinhelleren may have fished at deeper waters, where saithe are more numerous during the summer months (Ritchie et al., 2016). Still, one might expect that such a change was reflected in the fishing technology between the two sites. This is hard to see, as both sites are characterized by fishhooks of similar sizes (Bergsvik et al., 2016). The changes between the two sites may only be of local significance, considering that cod continue to dominate at other fjord sites contemporary to Olsteinhelleren (Faltinsen, 2018). However, since it is difficult to find reasonable environmental and cultural explanations for these changes in fish composition at Olsteinhelleren, we cannot rule out a decline of resources caused by overfishing of cod in the Hardanger fjord as a factor.
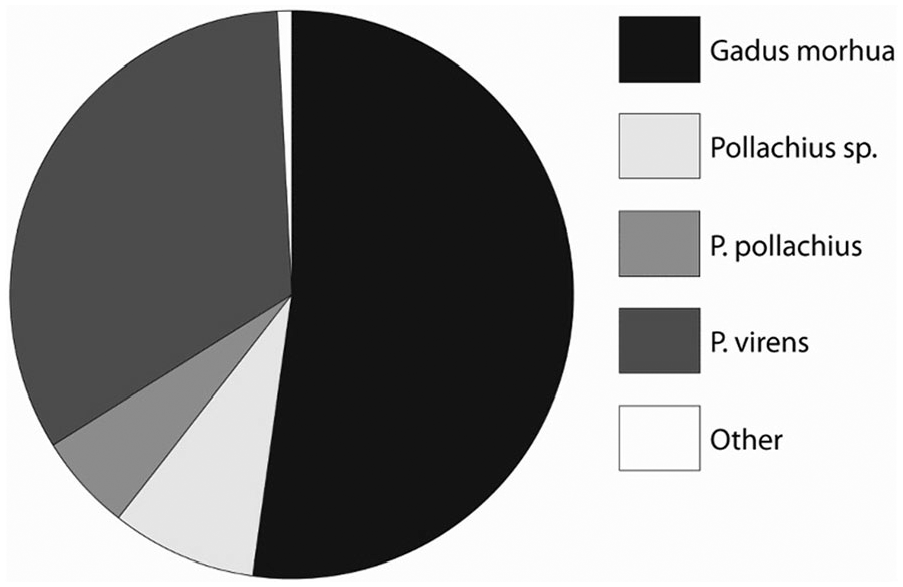
Relative abundances of gadid fish at Sævarhelleren. Figure originally in Ritchie et al. (2016).
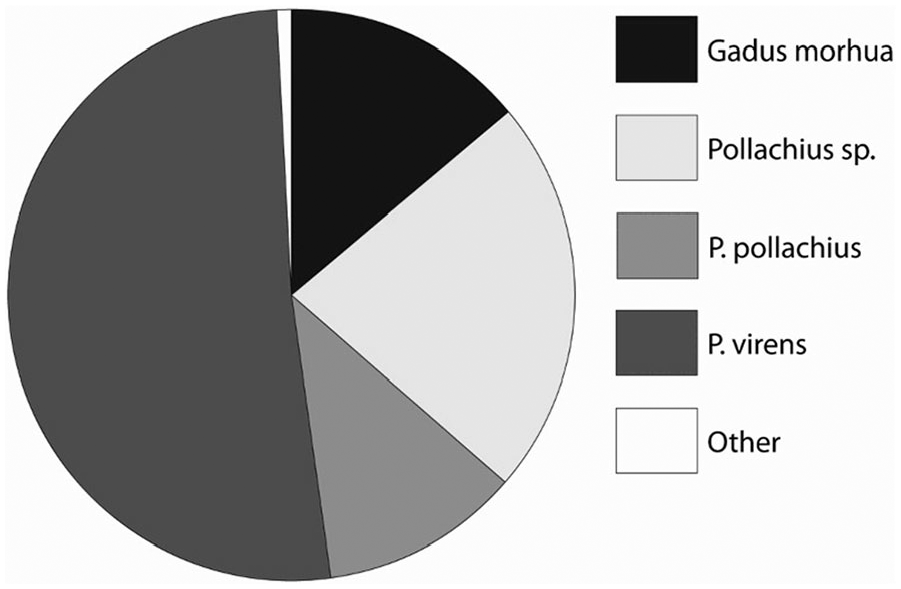
Relative abundances of gadid fish at Olsteinhelleren. Figure originally in Ritchie et al. (2016).
Final discussion and concluding remarks
While archaeological material has undergone numerous processes throughout the years and can in no sense be said to reflect the unbiased proxy of an environment at a particular point in time, it does provide several access points if the material is carefully selected. Given that the archaeological record is the only means of establishing a long-term data set of species variation and abundance in connection to climate change and human predation, the effort is worthwhile. Here, we have emphasized the human relation with marine, riverine and lacustrine environments, framed by a historical ecological perspective, to explore some of the parameters needed to create proxy data for studying and modelling the future effects of modern aquatic exploitation. We have highlighted several early-Holocene Scandinavian contexts and showed how the societies responsible for the archaeological remains can have had a long-term effect on the ecosystems they interacted with, which indeed can be useful for other disciplines not normally engaging with ancient societies and its long-term record.
In short, and highly simplified, through this study, we have demonstrated that fishing has been largely practised in Scandinavia from the early Holocene and onwards. Over time, this led to decreasing mobility and a corresponding increase of sedentary hunter-fisher-gatherer societies. Following the development of more stationary societies, specific areas became more important than others, which led to an increased territorialisation of the early- and mid-Holocene Scandinavian landscape. The increased territorialisation was supported and enabled by the practice of delayed-return subsistence strategies and the use of stationary food procurement technologies, which allowed the people to catch and store large quantities of food over long periods, which, consequently, anchored the human groups further to a particular area. The large-scale storage and the stationary food procurement equipment were connected to an increasing dependency on fish, which, in turn, can be connected to favourable conditions for aquatic life in early-Holocene Scandinavia. These favourable aquatic conditions, and an expanding sedentary lifestyle, led to a human population increase, which eventually led to increasing territorial displays and interpersonal violence, as the landscape became more crowded and the local aquatic abundancy declined. With a decline in aquatic abundancy followed a subsequent shift in main targeted species, illustrated through specific patterns in fish and aquatic mammal exploitation. This indicates that the early-Holocene societies of Scandinavia may have directly affected life in their local aquatic environments (Figure 8).

A simplified illustration of the events, indications, observations and implications influencing the human relationship with aquatic environments during early to mid-Holocene in Scandinavia.
However, to understand the full extent of ancient human interaction and impact on early-Holocene aquatic life and ecosystems, further analyses are needed. For example, the available fish bone assemblages need to be properly analysed, to detect temporal variations in species composition and intraspecies age variations, size reductions/increases and season of catch. Extensive stable isotope analyses (C and N) with a suitable temporal resolution should also be made to detect temporal changes in feeding habits among the different fish and aquatic mammal species themselves. Similar isotope analyses should also be made on the still unanalysed human and dog remains from early- and mid-Holocene contexts, to see whether the trend detected here is still valid when a larger sample becomes available.
If considering the evidence and data presented in this study, it might be argued that the intense use of aquatic resources was one of the driving forces behind the societal development in early-Holocene Scandinavia. Consequently, and given the criteria for creating comparable analogies as envisioned above, this ancient Scandinavian record meets the demands for what should be required of an archaeological material, if intended for use to develop models of future environmental scenarios in a world affected by global warming. Thus, we have demonstrated that the Scandinavian early- and mid-Holocene archaeological record is a good point of departure to gather appropriate proxy data useful for creating models of future aquatic ecosystem scenarios. Consequently, with this study, we are offering a long-term link between the ancient organic archaeological record and modern ecosystems. In other words, when appropriate analyses have been done, a true baseline, unbiased by earlier human predation on aquatic life, can be established. This baseline could be followed through time, and in different local environments, to form a record of how aquatic life develops when subjected to human predation. Furthermore, by correlating analyses on archaeological bone assemblages with climatic and environmental data obtained from, for example, lacustrine/marine sediments, peat bogs, tree rings and glacial records (cf. Bianchi and McCave, 1999; Björck et al., 1997; Jessen et al., 2005; Leng and Marshall, 2004), it is possible to relate the contingent changes observed in the zooarchaeological record, to fluctuations in temperature, in climate and in the natural environment. All considered, these data will be a valuable source of information and constitute a long-term (pre)historical ecological record. When this record is applied to modern contexts, it will enable the development of models to offer predictions of future environmental scenarios in different areas, with increases/decreases in temperature and other changes to the climate following the ongoing global warming. Finally, these models and prediction can also assist in assessing the future outcome when modern human exploitation of aquatic environments increases and/or if key species in our aquatic environments become locally extirpated.
This long-term link is, in its essence, based on the organic remains from the discarded waste of long-gone human cultures and societies. The future usefulness of this record is connected to how well we will be able to cope with the biases connected to the data. Unfortunately, the window of opportunity to mitigate the threat of failure to safeguard future aquatic systems from overexploitation and system collapses is constantly narrowing. As shown here, the ability to connect the long-term record of human aquatic exploitation patterns, during a period of rapid climate change, to changes in aquatic biodiversity and species abundance is intricately connected to the preservation and recovery of organic remains from appropriate sites and locations. Unfortunately, and often unknown even to researchers dealing with ancient remains on a daily basis, the ancient organic record itself is also threatened by our modern, post-industrial, way of life. Acid precipitation, pollution, extensive use of wetlands for agriculture, forestry, global warming, expanding residential areas and overall drainages to facilitate new constructions directly affect the ancient organic heritage. Recent studies indicate accelerated deterioration of organic archaeological remains (Boethius et al., in press; Boreham et al., 2011; High et al., 2016; Hollesen et al., 2018; Hollesen and Matthiesen, 2015; Jans et al., 2002; Kars et al., 2002; Milner et al., 2011; Nord et al., 2005, 2007; Nord and Lagerlöf, 2002; Nord and Tronner, 1991), suggesting that human encroachment into the environment has changed the prerequisites for in situ preservation. Also, the archaeological record in coastal areas is further threatened by erosion and sea-level rise as a consequence of the ongoing global warming (Erlandson, 2008). The general status of in situ archaeological organic remains is, however, still unknown in most areas and no major strategies have so far been suggested to stop the ongoing organic degradation. Consequently, the future of (pre)historic ecological modelling, using data derived from ancient organic remains, is threatened even before its full potential has been properly explored.
Footnotes
Acknowledgements
The authors would like to thank two anonymous reviewers for offering insightful comments on the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: The studies at Haväng and Hanöbukten were financed by the Swedish Research Council during the Blue Archaeology project (VR-2016-02027). The authors thank The Crafoord Foundation (grant number 20180631) for their part in financing this research.