
Abstract
Outbreaks of conifer bark beetles in Europe and North America have increased in scale and severity in recent decades. In this study, we identify existing fossil records containing bark beetle remains from the end of the Last Glacial Maximum (~14,000 cal. yr BP) to present day using the online databases Neotoma and BugsCEP and literature searches, and compare these data with modern distribution data of selected tree-killing species. Modern-day observational data from the Global Biodiversity Information Facility (GBIF) database was used to map recorded distributions from AD 1750 to present day. A total of 53 fossil sites containing bark beetle remains, from both geological and archeological sites, were found during our searches. Fossil sites were fewer in Europe (n = 21) than North America (n = 32). In Europe, 29% of the samples in which remains were found were younger than 1000 cal. yr BP, while in North America, remains were mainly identified from late Glacial (~14,000–11,500 cal. yr BP) sites. In total, the fossil records contained only 8 of 20 species we consider important tree-killing bark beetles in Europe and North America based on their impacts during the last 100 years. In Europe, Ips sexdentatus was absent from the fossil record. In North America, Dendroctonus adjunctus, Dendroctonus frontalis, Dendroctonus jeffreyi, Dendroctonus pseudotsugae, Dryocoetes confusus, Ips calligraphus, Ips confusus, Ips grandicollis, Ips lecontei, Ips paraconfusus, and Scolytus ventralis were absent. Overall, preserved remains of tree-killing bark beetles are rare in the fossil record. However, by retrieving bulk material from new and existing sites and combining data from identified bark beetle remains with pollen, charcoal, tree rings, and geochemistry, the occurrence and dominance of bark beetles, their outbreaks, and other disturbance events can be reconstructed.
Keywords
Introduction
Conifer bark beetles (Curculionidae: Scolytinae) often inflict density-dependent tree mortality (i.e. population growth rates and associated levels of tree mortality are partially regulated by the density of suitable hosts) and help maintain a diversity of tree species, ages, sizes, and spatial heterogeneity (Berryman, 1976). Endemic populations create small gaps in the forest canopy by killing trees usually stressed by age, drought, defoliation, or other factors. During outbreaks, large amounts of tree mortality may occur in short periods (e.g. 1 to several years) negatively affecting timber and fiber production, water quality and quantity, fish and wildlife populations, recreation, grazing capacity, biodiversity, endangered species, carbon sequestration and storage, and cultural resources, among others (Morris et al., 2018). In recent decades, bark beetle outbreaks in Europe and North America have increased in severity and scale (e.g. Hicke et al., 2016; Thorn et al., 2017) and several recent outbreaks are recognized among the most severe in recorded history, with high economic impacts (Bentz et al., 2009; Fettig et al., in press). Bark beetles are highly sensitive to thermal conditions conducive to population survival and growth (Bentz et al., 2010), and temperature-related drought stress negatively affects host tree vigor (Kolb et al., 2016). As such, outbreaks have been correlated with recent shifts in temperature and precipitation attributed to climate change. Forest densification has exacerbated this effect in many locations (Fettig et al., 2007).
Detailed, historical records document numerous outbreaks of different species of bark beetles in forests of Europe and North America since the 19th century (Bentz et al., 2009; Schelhaas et al., 2003; Wood, 1982). In Europe, outbreaks of the European spruce bark beetle (Ips typographus) on Norway spruce (Picea abies) are the most important cause of timber losses with millions of hectares of forest impacted annually (Schelhaas et al., 2003; Skuhravý, 2002). However, during the 20th century, the Great spruce bark beetle (Dendroctonus micans) has been expanding its geographic range and impacts across Europe (O’Neil and Evans, 1999). While the forest structure and economic damage from bark beetle outbreaks in Europe is substantial (Lieutier et al., 2004), the damage in North America is much higher (e.g. Hicke et al., 2012). There are several species in North America, from four different genera, which cause high tree mortality (Drooz, 1985; Fettig, 2016; Furniss and Carolin, 1977). The most notorious of these are in the genus Dendroctonus (Erichson). For example, species like Dendroctonus ponderosae and Dendroctonus rufipennis have had major impacts on forests in western North America in recent decades, destroying large numbers of trees and leading to major changes in forest function, structure, and composition (e.g. Berg et al., 2006; Kurz et al., 2008; Negrón and Fettig, 2014). Over the past 30 years, tree mortality caused by bark beetles in the western United States has exceeded tree mortality caused by wildfires (Hicke et al., 2016), raising concerns about the sustainability of some western forests to provide certain ecological goods and services over time. Most notable, D. ponderosae impacted ~10.3 million hectares of forest from 2000 to 2016, which represents almost half of the total area impacted by all bark beetles combined in the western United States during this period (Fettig et al., in press).
Although challenging, it is important to gain more insight in the relationships between other natural disturbance agents and bark beetle outbreaks (Dale et al., 2001; Fettig et al., 2013; Jenkins et al., 2014; Seidl and Rammer, 2017; Seidl et al., 2017; Thom and Seidl, 2016). A variety of models have been created to predict bark beetle outbreaks (e.g. Hicke et al., 2006; Stadelmann et al., 2013). These models allow us to calculate the chances of future bark beetle outbreaks on a regional as well as local scale, in order to assess their effects and to mitigate negative impacts (e.g. Fettig et al., 2007). The historical records of natural disturbances and bark beetle outbreaks, used in these models, go back as far as AD 1850 (Hicke et al., 2006; Schelhaas et al., 2003). In a few isolated studies, tree-ring records have been used in North America to identify local bark beetle outbreaks (Jarvis and Kulakowski, 2015; Zhang et al., 1999) which at certain locations date back to AD 1640. Dendroecological data sets from Central Europe (e.g. Čada et al., 2016) have been created to identify past natural disturbance events back to AD 1700. These records give us, up to a certain level, insight into what role bark beetles played in our forested landscapes before the significant increases in frequency and severity of outbreaks of the last several decades. In addition, identification of fossil bark beetle remains from sedimentary records can provide firsthand information of past outbreaks (Brunelle et al., 2008). Fossil beetle assemblages have been used to establish baselines of natural situations in managed parks (e.g. Mansell et al., 2014) as well as broader regional contexts (e.g. the reconstruction of Britain’s landscapes by Whitehouse and Smith, 2010). However, fossil records containing beetle remains are scattered and lacking from many regions, and an overview of fossil sites containing remains of tree-killing (primary) bark beetles is currently not available.
This study synthesizes available data of past occurrences of primary bark beetles by (1) summarizing the amount of fossil bark beetle data in publicly available databases and in the literature and (2) summarizing historical observational data from publicly available databases. Ultimately, we aim to improve knowledge on patterns (localities, frequency, mechanisms) of primary bark beetle outbreaks in Europe and North America.
Methods
Fossil records of bark beetles spanning the past 14,000 cal. yr BP were collated from sites in Europe and North America using the databases BugsCEP (Buckland and Buckland, 2006) and Neotoma (Williams et al., 2018). An additional data literature search with Elias (2010) provided a detailed overview of all sites <14,000 cal. yr BP containing bark beetle remains. Access to data and metadata unavailable as supplementary material in the literature was requested directly from the authors of the respective peer-reviewed publications. Additional unpublished data obtained directly from authors completed the final data collection (see unpublished; Supplemental Appendix C, available online). Taking the age constraint of 14,000 cal. yr BP and geographical limits into regard, ~500 sites from BugsCEP (Buckland and Buckland, 2006), 70 sites from Neotoma (Williams et al., 2018), and 20 additional sites were examined for primary bark beetle species remains. In Europe, the genera Ips, Dendroctonus, and Pityogenes were selected for query. Although species of the genus Pityogenes do not cause substantial damage to conifers in Europe, historical and recent outbreaks have been frequent (e.g. Göthlin et al., 2000; Grodzki, 1997; Zúbrik et al., 2008). Furthermore, as Pityogenes chalcographus favors the same host trees as I. typographus and D. micans (Göthlin et al., 2000; Novotný et al., 2002), its presence in the fossil record could indicate past conditions favorable for outbreaks of I. typographus or D. micans. The genera Ips, Dendroctonus, Scolytus, and Dryocoetes were selected for the query of North American sites. Metadata including location, latitude, longitude, altitude, site type (e.g. archeological site, peat bog), additional paleoecological proxies, age of sediments, number of samples, and reference were recorded (see Supplemental Data, available online). Sites were divided into time periods according to a formal subdivision of the middle Holocene and late Holocene by Walker et al. (2012): late Glacial (~14,000–11,500 cal. yr BP), early Holocene (~11,500–8200 cal. yr BP), middle Holocene (~8200–4200 cal. yr BP), and late Holocene (~4200–1000 cal. yr BP). In addition, the last ~1000 cal. yr BP to present were defined as an extra time period called ‘Historical’, which includes archeological sites as well as lake and peat bog sites. Samples with bark beetle remains were attributed to a certain time period by comparing the depth of the sample with the published age-depth model of the respective site. From the selected sites, not only data from the queried genera but all identified primary and secondary bark beetles were recorded. Identifications to genus level (e.g. Ips sp.) were also included in these records.
Modern observational data
Modern bark beetle distribution data were collated from the Global Biodiversity Information Facility (GBIF, 2018a). Recordings without coordinates were deleted and recurring coordinates were combined if these observations were recorded during the same year. Records from museums were removed from the results as well, as these do not reflect the original locality of the beetles. For North America, additional observational data were added from approximated localities described by Wood (1982). The locations of fossil sites with identified primary bark beetle remains were compared with the observed modern species distribution by plotting point locations in R (R Core Team 2018, version 3.5.0). In addition, the historical observation data from GBIF (2018a) were divided into 20-year time periods (AD 1750–2018).
Results
A total of 53 fossil records, 21 from Europe and 32 from North America, were identified as containing bark beetle remains of the queried genera (Figure 1; Supplemental Appendix A, available online). The age of sites ranged from late Glacial to present day, and all but two sites contained radiocarbon dated samples. Site classifications included archeological sites, (paleo) beach deposits, river deposits, lake deposits, and peat deposits. Three of the 21 sites found in Europe were records from archeological studies, while none of the 32 sites in North America were archeological sites. Many of the sites found in North America were used to identify the extent and timing of deglaciation (e.g. Ashworth et al., 1981; Mott et al., 1981). This difference in research foci is reflected in the ages of the different sites in Europe and North America. While only 2 of the 21 sites containing bark beetle remains of the queried genera in Europe included late-Glacial sediments (Ponel et al., 2001, 1999), 12 of the 32 sites in North America were late Glacial sites. In North America, only four fossil sites contained samples younger than 1000 cal. yr BP, but none of these samples contained any of the bark beetle species which are considered to cause substantial damage (Bentz et al., 2009; Fettig et al., in press; Morris et al., 2017; Table 1).

Sites with fossil records in Europe and North America, dated 14,000 cal. yr BP to present, where Ips, Dendroctonus, and/or Pityogenes (Europe) or Dendroctonus, Ips, Scolytus, and/or Dryocoetes species (North America) remains were identified. Based on the data obtained from BugsCEP (Buckland and Buckland, 2006) and Neotoma (Williams et al., 2018).
Bark beetle species (Scolytinae) that caused substantial damage to stands of conifer trees in Europe and North America in recent decades.
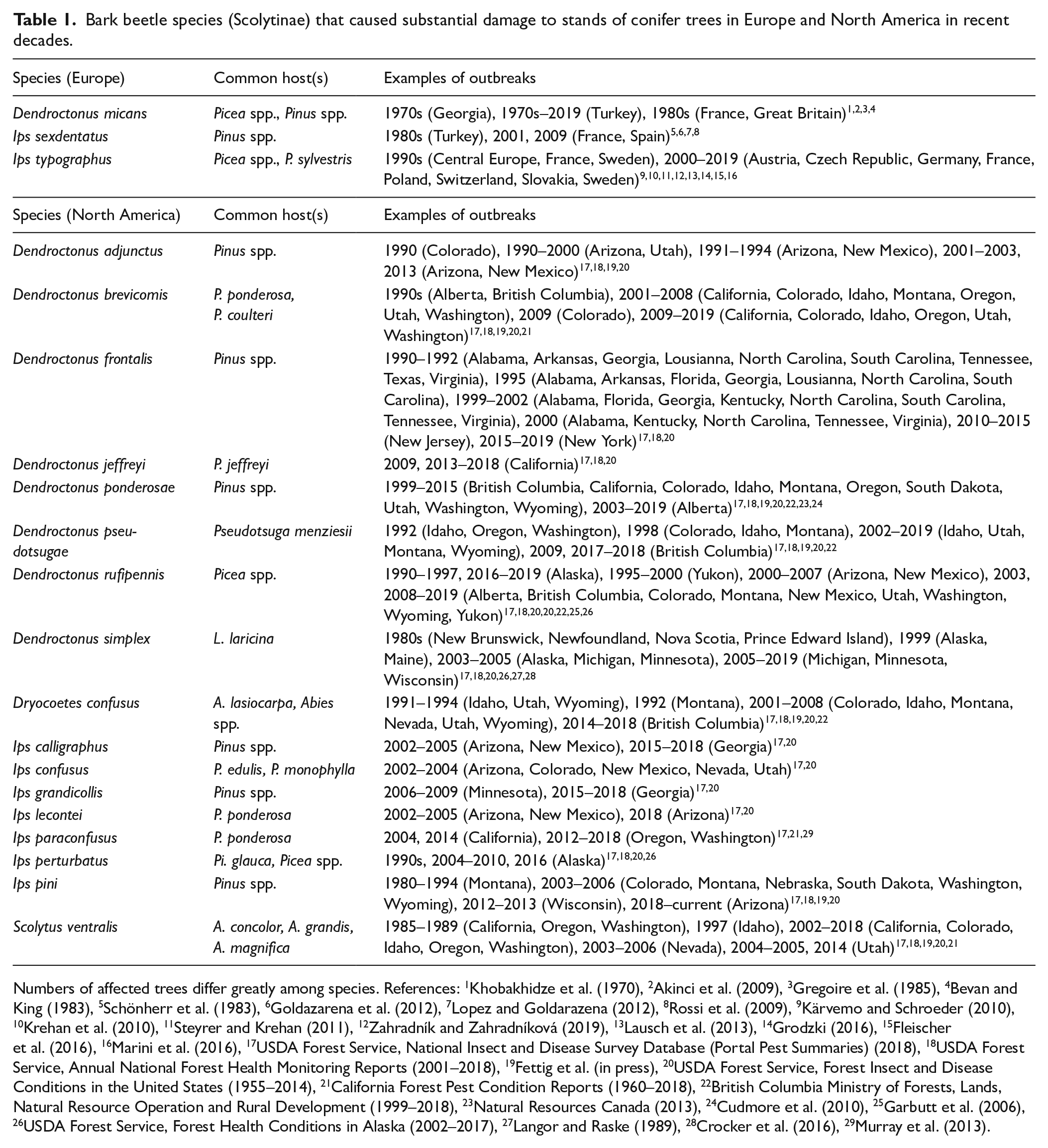
Numbers of affected trees differ greatly among species. References: 1Khobakhidze et al. (1970), 2Akinci et al. (2009), 3Gregoire et al. (1985), 4Bevan and King (1983), 5Schönherr et al. (1983), 6Goldazarena et al. (2012), 7Lopez and Goldarazena (2012), 8Rossi et al. (2009), 9Kärvemo and Schroeder (2010), 10Krehan et al. (2010), 11Steyrer and Krehan (2011), 12Zahradník and Zahradníková (2019), 13Lausch et al. (2013), 14Grodzki (2016), 15Fleischer et al. (2016), 16Marini et al. (2016), 17USDA Forest Service, National Insect and Disease Survey Database (Portal Pest Summaries) (2018), 18USDA Forest Service, Annual National Forest Health Monitoring Reports (2001–2018), 19Fettig et al. (in press), 20USDA Forest Service, Forest Insect and Disease Conditions in the United States (1955–2014), 21California Forest Pest Condition Reports (1960–2018), 22British Columbia Ministry of Forests, Lands, Natural Resource Operation and Rural Development (1999–2018), 23Natural Resources Canada (2013), 24Cudmore et al. (2010), 25Garbutt et al. (2006), 26USDA Forest Service, Forest Health Conditions in Alaska (2002–2017), 27Langor and Raske (1989), 28Crocker et al. (2016), 29Murray et al. (2013).
Europe
In Europe, the species identified from fossil remains that cause substantial damage to trees were I. typographus (six sites) and D. micans (one site), but also fossil remains of Pit. chalcographus (nine sites) were found (Figure 2; Supplemental Appendices B and C, available online). All fossil records with these three identified species were younger than late Glacial. In samples covering the early Holocene, one site (Prašilske Lake, Czech Republic) contained remains of Pit. chalcographus. During the middle Holocene, two sites (Lac Long Inferior, France; Prašilske Lake, Czech Republic) contained remains from Pit. chalcographus, where at the Czech site, these remains were found together with those of I. typographus. Pit. chalcographus was found in samples from the late Holocene at two sites (Lac Long Inferior, France; Piilonsuo, Finland), and the Finnish site Piilonsuo (Koponen and Nuorteva, 1973) contained remains of I. typographus as well. Historical (~1000 cal. yr BP to present) sites in Europe contained most of the bark beetle remains of I. typographus, Pit. chalcographus, and D. micans. The Medieval archeological site of Novgorod, Russia (Hellqvist, 1999), contained fossil records of all three species I. typographus, Pit. chalcographus and D. micans. Five more historical (~1000 cal. yr BP to present) sites contained remains of Pit. chalcographus, three of these also contained remains of I. typographus (Oslo: Gamlebyen, Norway; Stavsåkra, Sweden; Storasjö, Sweden; Laka Lake, Czech Republic; and Diera Hollow, Slovakia; Oslo: Gamlebyen being an archeological site). Unpublished data from Diera Hollow in Slovakia (unpublished data by author) showed higher numbers of individuals of Pit. chalcographus between 950 and 400 cal. yr BP.

Modern distributions of three tree-killing bark beetle species in Europe, next to locations of the fossil sites with their identified remains which were dated Early Holocene (~11,500–8200 cal yr BP), Mid-Holocene (~8200–4200 cal yr BP), Late Holocene (~4200-1000 cal yr BP) and 1000 cal yr BP-present. Coordinates of modern observations were downloaded from GBIF.org. Ips typographus (green crosses), Dendroctonus micans (yellow circles), and Pityogenes chalcographus (blue diamonds). Depicted sites with Early Holocene fossils are Prašilske Lake (unpublished data); Mid-Holocene sites are Prašilske Lake (unpublished data), Lac Long Inferior (Ponel et al., 2001); Late Holocene sites are Piilonsuo (Koponen and Nuorteva, 1973) and Lac Long Inferior (Ponel et al., 2001); <1000 cal yr BP sites are Oslo: Gamlebyen (Kenward, 1988), Stavsåkra (Olsson and Lemdahl, 2009), Storasjö (Olsson and Lemdahl, 2010), Novgorod: Troitski (Hellqvist, 1999), Prašilske Lake (unpublished data), Laka Lake (unpublished data) and Diera Hollow (unpublished data).
North America
In North America, the species identified from fossil remains that cause substantial damage were D. ponderosae (two sites), D. rufipennis (eight sites ranging from late Glacial to late Holocene), Dendroctonus brevicomis (one site), Dendroctonus simplex (two sites), Ips perturbatus (two sites), and Ips pini (two sites) (Figure 3; Supplemental Appendices B and C, available online). During the late Glacial, four different species were found: D. rufipennis, D. simplex, I. perturbatus, and I. pini. During the early Holocene, D. rufipennis, I. perturbatus, and I. pini were found. Here, D. rufipennis was identified at three different sites in two different regions of North America (Rocky Mountains (Elias et al., 1986, 1991) and Eastern Canada (Ashworth, 1977)). One site (Lake Emma, Colorado: Elias et al., 1991) showed an indication of a higher abundance of D. rufipennis in two samples dated 10,000–9000 cal. yr BP. Two sites located in the Rocky Mountains in the United States contained remains from D. rufipennis (Elias, 1985; Elias et al., 1986), and two other sites in the same region contained remains from D. ponderosae during the middle Holocene (Brunelle et al., 2008). A fifth site located in Eastern Canada contained remains of D. simplex (Morgan et al., 1985). During the late Holocene, two sites in North America contained remains of D. rufipennis and D. brevicomis. One of the sites, located west of Hudson Bay, Canada (Elias, 1982), contained remains of D. rufipennis, and the other site, located in the Rocky Mountains in the United States (Elias et al., 1986), contained remains of D. rufipennis and D. brevicomis.

Modern distributions of six tree-killing bark beetle species in North America, next to locations of the fossil sites with their identified remains which were dated Late Glacial (~14,000-11,500 cal yr BP), Early Holocene (~11,500-8200 cal yr BP), Mid-Holocene (~8200-4200 cal yr BP) and Late Holocene (~4200-1000 cal yr BP). Coordinates of modern observations were downloaded from GBIF.org and supplemented with approximate locations from Wood (1982). Dendroctonus ponderosae (red diamonds), Dendroctonus rufipennis (blue triangles), Dendroctonus brevicomis (orange stars), Dendroctonus simplex (yellow diamonds), Ips perturbatus (green squares) and Ips pini (pink triangles). Depicted sites with Late Glacial fossils are St. Hillaire (Mott et al., 1981), Johns Lake (Ashworth and Schwert, 1992), Norwood (Ashworth et al., 1981) and Huntington Dam (Elias et al., 1991); Early Holocene sites are Seibold (Ashworth and Brophy, 1972), 18 Miles River (Ashworth, 1977), Gage Street Site (Schwert et al., 1985), Lake Isabella Delta (Elias et al., 1986) and Lake Emma (Elias et al., 1991); Mid-Holocene sites are Hoodoo Lake (Brunelle et al., 2008), Baker Lake (Brunelle et al., 2008), Au Sable River (Morgan et al., 1985), La Poudre Pass LP5 (Elias et al., 1986) and Lake Isabella Peat (Elias, 1985); Late Holocene sites are Ennadai II (Elias, 1982) and Roaring River (Elias et al., 1986).
In total, the fossil records contained 8 out of the 20 species that are considered (potentially) destructive bark beetles (Table 1). In Europe, no remains of Ips sexdentatus were found. In North America, the species Dendroctonus adjunctus, Dendroctonus frontalis, Dendroctonus jeffreyi, Dendroctonus pseudotsugae, Dryocoetes confusus, Ips calligraphus, Ips confusus, Ips grandicollis, Ips lecontei, Ips paraconfusus, and Scolytus ventralis were absent from the fossil record. Remains which were identified to genus level could account for some of these missing species, although this concerns a low number of identifications (data from two sites in Europe with identified Ips sp., and six sites in North America: five with identified Dendroctonus sp. and one with identified Ips sp.; see Supplemental Data, available online). Figures 4 and 5 give an overview of the collated data and show the number of sites with target species, remains identified to genus level, and different species from the same genera as were queried.

Fossil sites in Europe with occurrences in different time periods of the three queried taxa Ips, Pityogenes and Dendroctonus and the target species I. typographus, Pit. chalcographus and D. micans. A single fossil site can contain multiple time periods.

Fossil sites in North America with occurrences in different time periods of the queried taxa Ips and Dendroctonus and the target species I. perturbatus, I. pini, D. ponderosae, D. brevicomis, D. rufipennis and D. simplex. A single fossil site can contain multiple time periods.
Modern observational data
Modern observational data from GBIF (2018b-i, 2019a) for Europe and North America are shown in Figures 6 and 7. For North America, the lowest amount of data points for the query species was found for D. simplex (14 data points) and the highest for I. pini (144 data points), the latter one of the most common bark beetles in North America (Furniss and Carolin, 1977).

Historical and modern-day observations of Ips typographus (green crosses), Dendroctonus micans (yellow circles) and Pityogenes chalcographus (blue diamonds). Downloaded observational data from GBIF.org was split into one time period of 1750-1849 AD, one time period of 1850–1899 AD, and afterwards time periods of 20 years.

Historical and modern-day observations of Dendroctonus ponderosae (red diamonds), Dendroctonus rufipennis (blue triangles), Dendroctonus brevicomis (orange stars), Dendroctonus simplex (yellow diamonds), Ips perturbatus (green squares) and Ips pini (pink triangles). Downloaded observational data from GBIF.org was split into one time period of 1750–1849 AD, one time period of 1850–1899 AD, and afterwards time periods of 20 years.
Discussion
Distribution and abundance of fossil and historical primary bark beetles
Here, we present the first synthesis of fossil records of primary bark beetles in Europe and North America. Fossil sites that contained remains of primary bark beetles were scattered, and most consisted of a single identification of a species within a fossil record. In North America, the only exception was the site Lake Emma, Colorado (37.90211°N, 107.61537°W), where remains of 3 to 11 individuals of D. rufipennis were found in three sediment samples from ~10,000 cal. yr BP (Elias et al., 1991). In the region where Lake Emma is located], severe outbreaks of D. rufipennis have been occurring since AD ~2000 (Colorado State Forest Service, 2017). Such a high abundance of D. rufipennis in the fossil record of a site, which lies in a region where outbreaks are occurring currently, provides strong evidence that this area has been subjected to D. rufipennis outbreaks at least since the early Holocene. Due to the low abundances of bark beetle fossils in general, presence of species in the fossil record has previously been interpreted in a qualitative way (e.g. Schwert et al., 1985) to indicate the presence of certain plant or tree species or climatic conditions (although bark beetle species have been pooled with other species living in dead wood to form functional groups in paleoenvironmental reconstructions; e.g. Olsson and Lemdahl, 2010). In many of the studies where bark beetles were used to indicate the presence of trees or forests, no more than one or two individuals were found per sample and no species had a continuous presence throughout a stratigraphic sequence (e.g. Brunelle et al., 2008). Other species, such as D. frontalis, D. pseudotsugae, and S. ventralis, are missing completely in the fossil record, even though extensive outbreaks of these species have occurred historically (Table 1). For some species (D. frontalis, Ips spp.), this absence can be explained by a lack of fossil sites within their modern distribution range. For example, Neotoma (Williams et al., 2018) shows no fossil sites with insects <14,000 BP for a large portion of the southeastern United States. Possible explanations include a lack of suitable sites for preservation of remains and/or an absence of research interests (for an overview, see Elias, 2010). It is important to note that sites in this study might have been poorly dated, and although great care was taken to attribute samples with bark beetle remains to their appropriate time period, this has been highly dependent on the recorded age of the sites. Nevertheless, fossil records with primary bark beetles are scarce.
There were more sites with fossil records of primary bark beetles found in North America, but the European sites contain more occurrences. I. typographus and Pit. chalcographus co-occurred at six (29%) fossil sites. At Novgorod (Hellqvist, 1999), I. typographus was found together with both D. micans and Pit. chalcographus. Results from these fossil sites are a good indication that at least in historical times (since ~1000 cal. yr BP) primary bark beetle species were co-occurring in the same areas of Europe as they are today (Figure 2). A lack of occurrences of I. typographus in the fossil record of France, one of the countries that has experienced several notable outbreaks (Sallé et al., 2005; Viiri and Lieutier, 2004), is difficult to explain although the location of fossil sites likely has some influence as older publications report I. typographus populations were confined to mountainous areas (Komárek, 1925; Pfeffer, 1930). Despite this, fossil sites from France and Sweden, incorporated in the current data set, yielded remains from other Ips spp. such as Ips accuminatus (Storasjö, Sweden during the early Holocene; La Borde, France during the early Holocene; Tailefer Massif, France during the late Holocene). As I. accuminatus colonizes Pinus sylvestris in France (e.g. Herard and Mercadier, 1996), it might be worthwhile to consider this species in future research.
Differences in the types of fossil sites and original study objectives in Europe and North America seem to have resulted in differences between the fossil records from these continents, as well as low amounts of recorded bark beetle remains for certain time periods. In North America, some of the studies that recovered primary bark beetle remains focused on reconstructing late-Glacial to early-Holocene temperatures (e.g. Morgan and Morgan, 1979), while others focused on the retreat of the Laurentide Ice Sheet after the Last Glacial Maximum and subsequent landscape dynamics (e.g. Schwert et al., 1985). For these type of studies, single occurrences of primary bark beetle remains were sufficient evidence of the presence of host trees at the study site (e.g. Ashworth et al., 1981; Elias, 1982). In Europe, fossil insect remains have been used to reconstruct late-Glacial to early-Holocene climates (Coope et al., 1998). Many of these sites are located in Great Britain (Elias, 2010), a region where few remains of conifer bark beetles have been found. All identified species from records of Great Britain are colonizing Pinus spp. rather than Pi. abies. Moreover, the majority of bark beetle species found in Great Britain are species that colonize deciduous trees. This likely clarifies why the relatively large number of fossil sites in Europe yield a very low abundance of the queried species.
We only found one study, conducted by Brunelle et al. (2008), which attempted to specifically identify primary bark beetle remains in the fossil record of a site. This study was conducted at Hoodoo Lake, Idaho, US, and correlated the occurrence of remains with past disturbance events. Given the assumption that bark beetle outbreaks are likely to occur when stands of trees are experiencing stress, such as during drought (Kolb et al., 2016), their study focused on identifying bark beetle remains from lake sediments in an area that experienced frequent bark beetle outbreaks. The only remains of D. ponderosae were found originated from sediments that were dated at 8200 cal. yr BP, and as such it was speculated that bark beetle outbreaks may have occurred during the 8.2 Ka event (Brunelle et al., 2008). However, attempts to reproduce these results proved difficult as additional cores from Hoodoo Lake, and other nearby lakes, yielded no remains of D. ponderosae (Morris et al., 2015). In Europe, recent unpublished multiproxy data from Prašilske Lake and Laka Lake, Czech Republic and Diera Hollow, Slovakia (see unpublished in Supplemental Data, available online) indicate that fossil remains of primary bark beetles from lake sediments and peat cores could be used to detect fluctuations in bark beetle populations through time. In these three studies, bark beetle remains were used as one of the proxies to detect other natural disturbances.
The data from GBIF (2018b-i, 2019a) for Europe and North America provide a good overview of the regional representation of modern bark beetle distributions. For example, observational records from GBIF (2018b) for D. ponderosae appear to demonstrate its northward range expansion between 1980 and 2018 (e.g. Robertson et al., 2009). However, many areas that have experienced substantial outbreaks of D. ponderosae, as well as other primary bark beetles in North America, are not well represented in GBIF (2018b-g) but are available elsewhere (e.g. for the United States, see www.fs.fed.us/foresthealth/publications). Modern distribution data for I. typographus, D. micans, and Pit. chalcographus are concentrated in Western Europe and Scandinavia and lacking in Eastern Europe, coherent with the fact that the majority of GBIF data collectors reside in Western Europe and Scandinavia. As with similar databases, GBIF (2018a) depends on individuals for uploading data and some regions as well as certain species are better represented than others. Furthermore, it can be argued that a rapid increase of data from 1980 onwards could also be explained by improved methods of data collation and a rise in interest in the occurrences of certain bark beetle species.
Primary bark beetles as indicators of disturbance dynamics
Fossil bark beetle records hold great potential for inference of changes in forest composition and other disturbance regimes. However, records from the British Isles, where the majority of the studies on fossil beetle remains have been executed contained few conifer bark beetles. The few attempts to correlate trends in pollen fluctuations with bark beetle remains were criticized as circumstantial (Girling and Grieg, 1985). As no evidence exists for fluctuations in deciduous bark beetle populations from the numerous British records, the focus in historical landscape studies shifted to the ‘Urwald fauna’, beetle species indicative of primary forests (e.g. Whitehouse, 2006). At several sites in Britain, however, specimens of Pityogenes quadridens and Pityogenes bidentatus were found (see Supplemental Appendix B, available online), species with Pinus spp. as host or main host. As shown in GBIF (2019b), Pi. abies is not common in all regions of the British Isles, and future studies on conifer bark beetles in this region should include species colonizing Pinus spp.
Together with other proxies such as fossil pollen and sedimentary charcoal, fossil bark beetle remains may be used to reconstruct past disturbance history of sites (unpublished data by author). For example, fossil records with the absence of primary bark beetles but the presence of secondary bark beetles that only colonize dead or dying trees (see Supplemental Appendix B, available online) might indicate large amounts of dead wood resulting from other causes than a bark beetle outbreak. Combining all this information, the history and mechanisms of natural disturbances in conifer forests could be reconstructed. This could provide valuable new information for models to predict future bark beetle outbreaks (Morris et al., 2017). At many sites in Europe and North America, data from proxies such as pollen, charcoal, tree rings, and geochemical data already exist and have been used to reconstruct disturbance histories on local to regional scales (e.g. Tinner et al., 2008; Willis et al., 2000). Additional bulk material from waterlogged sites like peat bogs or small lakes could yield insect remains, in many cases in amounts large enough to make qualitative (small lakes) or even quantitative (peat bogs) reconstructions. These reconstructions could be used to establish baselines for primary bark beetle species abundance from the early to late Holocene, as well as to provide additional information about the distribution of these species in historical times (Figures 2–7).
Conclusion
In this study, we take the initial steps toward identifying and integrating fossil and historical data of primary bark beetle occurrences in Europe and North America. The scarcity of bark beetle remains in fossil records, especially in North America, represents a critical gap in our understanding of ecological history, and a fruitful area of future research in the field of paleoecology. Historical observational data from GBIF (2018b-g) for North America are also scarce. As such, we encourage researchers in North America to consider contributing more records to this and similar databases in hopes of increasing their overall utility. Both fossil and historical records in Europe are more numerous, but concentrated in Western Europe. New unpublished data from Central Europe show an abundance of primary bark beetle remains at sites located in coniferous forests. By retrieving bulk material from new and existing sites and combining data from identified bark beetle remains with pollen, charcoal, tree rings, and geochemistry, the dominance, presence, or absence of primary bark beetles during natural disturbance events could be reconstructed. Further synthesis of fossil and historical data, from geological sites as well as archeological sites, will increase our understanding of past disturbances.
Supplemental Material
Appendix_A_Sites – Supplemental material for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics
Supplemental material, Appendix_A_Sites for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics by Nick Schafstall, Niina Kuosmanen, Christopher J Fettig, Miloš Knižek and Jennifer L Clear in The Holocene
Supplemental Material
Appendix_B_Species_and_their_host_trees – Supplemental material for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics
Supplemental material, Appendix_B_Species_and_their_host_trees for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics by Nick Schafstall, Niina Kuosmanen, Christopher J Fettig, Miloš Knižek and Jennifer L Clear in The Holocene
Supplemental Material
Appendix_C_All_data – Supplemental material for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics
Supplemental material, Appendix_C_All_data for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics by Nick Schafstall, Niina Kuosmanen, Christopher J Fettig, Miloš Knižek and Jennifer L Clear in The Holocene
Supplemental Material
Appendix_D_References – Supplemental material for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics
Supplemental material, Appendix_D_References for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics by Nick Schafstall, Niina Kuosmanen, Christopher J Fettig, Miloš Knižek and Jennifer L Clear in The Holocene
Supplemental Material
Captions_for_supplementary_material_Schafstall_et_al_Holocene – Supplemental material for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics
Supplemental material, Captions_for_supplementary_material_Schafstall_et_al_Holocene for Late Glacial and Holocene records of tree-killing conifer bark beetles in Europe and North America: Implications for forest disturbance dynamics by Nick Schafstall, Niina Kuosmanen, Christopher J Fettig, Miloš Knižek and Jennifer L Clear in The Holocene
Footnotes
Acknowledgements
The authors thank Joey Pettit and Mélanie Saulnier for assistance in developing the distribution maps. They specially thank Thomas Atkinson for his help with the list of bark beetle species and their host trees in Supplemental Appendix B. The response of researchers to authors’ requests for additional information concerning their findings is greatly appreciated, and the authors specially thank Scott Elias for providing and checking unpublished beetle count sheets from various sites. Finally, they acknowledge Neotomadb.org, BugsCEP.com, and ![]() for providing freely accessible databases and all data contributors who provided their data to these databases for future use.
for providing freely accessible databases and all data contributors who provided their data to these databases for future use.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Nick Schafstall, Niina Kuosmanen, and Jennifer Clear were supported by the Czech Science Foundation (project no. 16-23183Y; PI: Jennifer L. Clear). Nick Schafstall was also supported by the Internal Grant Agency of the Faculty of Forestry and Wood Sciences of the Czech University of Life Sciences in Prague (IGA grant A06-18). Niina Kuosmanen also received support from project EXTEMIT-K: CZ.02.1.01/0.0/0.0/ 15_003/0000433. Christopher Fettig received support from the Pacific Southwest Research Station. A portion of the research of Miloš Knížek was supported by the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO0118.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.