
Abstract
Rice is among the world’s most important and ancient domesticated crops. However, the spatial and temporal pattern of the early rice domestication process remains unclear due to the lack of systematic study of wild/domesticated rice remains and corresponding dates during the early Holocene. Here, we collected 248 samples from five typical Shangshan cultural sites in the lower Yangtze region where is the most likely origin place of rice for phytolith analysis. The results showed the following. (1) Rice bulliform phytoliths from the five sites all present domestication traits, suggesting that the rice domestication process had begun across the region by the early stage of the Holocene. (2) The relative domestication rates reflected by the rice bulliform phytoliths were different between sites, the sites with higher domestication rates were distributed closer to the mainstream river. (3) The rice domestication process revealed by bulliform phytoliths can be divided into three periods during the early Holocene: from 10 to 9 ka, rice domestication began and stayed at a low level under 35%; from 9 to 8.5 ka, rice domestication level increased to 50%; and from 8.5 to 8 ka, rice domestication level was in a fluctuating state. (4) By 9 ka BP, rice double-peaked phytoliths from glume cells are present in most of the studied sites, which imply the presence of crop dehusking processing. This study reconstructed the spatial and temporal patterns of rice domestication during the early Holocene, which will improve our knowledge of early crop domestication and enhance our understanding of changes in rice status.
Introduction
Reconstructing the spatial and temporal pattern of crop domestication can help us understand environmental factors that govern the expansion of plant ranges and the relationship between crop domestication and human activity history (Gutaker et al., 2020). Rice (Oryza sativa L.), a staple component of the diet of half the world’s population, is among the world’s most important and ancient domesticated crops (Callaway, 2014). A recent study based on direct phytolith dating has shown that rice domestication may have begun along the lower Yangtze River at the beginning of the Holocene (Zuo et al., 2017). The origins of rice have been the focus of intensive study, while relatively little attention has focused on the early process of spatial and temporal rice domestication.
The main reasons that hinder the reconstruction of spatial and temporal patterns of early rice domestication are as follows. (1) It is difficult to find carbonized rice grains and spikelet bases that can be directly dated to 9000 years old (Fuller et al., 2007; Liu et al., 2007; Zhao, 2014; Zheng and Jiang, 2007; Zheng et al., 2007) due to the humid climate in the lower Yangtze region, resulting in poor preservation of organic materials. Thus, it is difficult to find enough samples associated with early occupations. (2) Phytolith evidence has been sued to reconstruct the basic framework of rice domestication from 10,000 to 2000 BP (Ma et al., 2016, 2018; Wu et al., 2014), but this work lacks sufficient samples that reflect the spatial variation and corresponding accurate dating results to reconstruct the earliest spatial and temporal pattern of rice domestication during 10,000–8000 BP.
Phytoliths, which are silica particles that make up intercellular or intracellular structures in plants, are particularly abundant in Poaceae plants and can be well preserved in soils (Piperno, 2006; Wang and Lu, 1993). Phytoliths have been a useful tool in exploring the domestication of crops (Ball et al., 2016). Rice plants produce a variety of phytoliths, and the types of bilobate, bulliform and double-peaked phytoliths exhibit taxonomical value (Gu et al., 2013; Lu et al., 2002; Wang and Lu, 1993, 2012). Recently, a study examined rice bulliform phytoliths from wild and domesticated rice paddy soils and found that in wild rice soils the proportion of bulliform phytoliths with ⩾9 fish-scale decorations was 17.46% ± 8.29% while in domesticated rice soil was 63.70% ± 9.22% (Huan et al., 2015). The study demonstrated that the proportion of bulliform phytoliths with ⩾9 fish-scale decorations can be adopted as a criterion for discriminating wild and domesticated rice and might reflect the relative domestication rate to a certain extent (Lu, 2017).
Since the 1970s, the lower Yangtze region has been considered the origin place of rice domestication (Jiang, 2004; Sun et al., 2007; Zhejiang Provincial Administration Committee of Cultural Relics and Zhejiang Museum, 1978; Zhejiang Provincial Institute of Archaeology, 2003). The Shangshan culture was by far the earliest Neolithic culture in this region and can be dated to 10,000–8000 BP (Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2007; Zuo et al., 2017). The Shangshan culture provides an archaeological context and key evidence for the study of early rice origin and domestication. However, most previous studies have only focused on individual site (Qiu et al., 2019; Zheng et al., 2016; Zuo et al., 2017), and lack regional investigations, which greatly hinders accurate study of the spatial and temporal pattern of the rice domestication process. In the last two decades, more than 10 sites associated with the Shangshan culture have been discovered (Jiang et al., 2017, 2018; Xu et al., 2016; Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2007), providing an opportunity to systematically study the spatial and temporal patterns of early rice domestication.
Here, we collected 248 samples from five typical archaeological sites of Shangshan culture and used phytolith analysis to trace the early rice domestication process in the lower Yangtze region in order to construct the spatial and temporal pattern of rice domestication during the early Holocene. This study improves our knowledge of early crop domestication and enhances our understanding of changes in the rice status.
Materials and methods
Sediment and sampling
The Shangshan culture is mainly distributed in the valley of the Qujiang River and Jinhua River, which cross the Jinqu Basin, the largest basin in Zhejiang Province (Xu et al., 2016). This study analysed samples from five Shangshan culture sites, namely, the Shangshan site, Miaoshan site, Hehuashan site, Huxi site and Qiaotou site. The Shangshan site is locate in the valley of the upper Puyang River, which is a small marginal basin outside of the Jinqu Basin. The Hehuashan site is located beside the Qujiang River, which is the upper mainstream of the Fuchun River. The Miaoshan site and Huxi site are located beside the Wuyi River. The Qiaotou site is located on the Dongyang River. The Wuyi River and Dongyang River merge in Jinhua city and form the Jinhua River, which is a tributary of the Qujiang River (Figure 1).
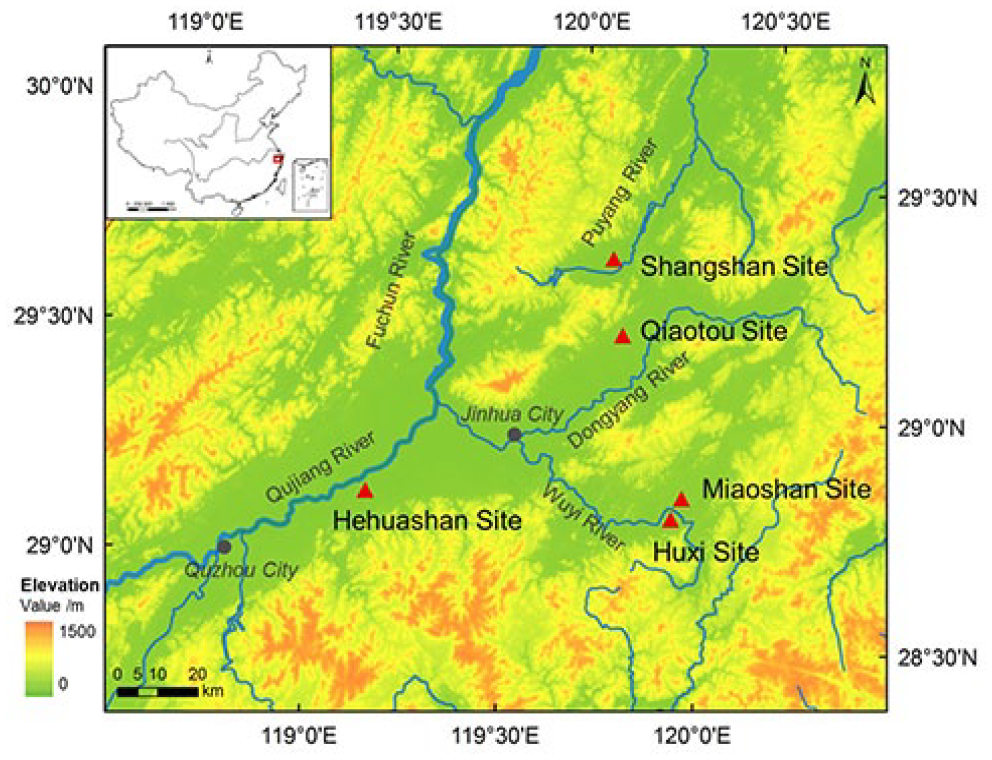
Locations of the studied Shangshan culture archaeological sites.
The Shangshan site (29.46°N, 119.97°E) is the earliest Neolithic site that has been systematically excavated in the lower Yangtze region to date (Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2007). The Shangshan site has eight stratigraphic layers, among which layers 8 and 7 contain pottery and stone artefacts from the early Shangshan cultural period, layers 6 and 5 contain pottery from the late Shangshan period, layer 4 contains Hemudu-Majiabang cultural materials, layers 3 and 2 contain materials of the historical period, and layer 1 is the modern topsoil. A total of 24 soil samples from T0611 were collected (including 20 samples at 2 cm intervals from layer 8 to 4 and 4 samples at 5 cm intervals from layer 3 to 1) for phytolith analysis (Figure 2).

The sampling profiles in the selected sites of Shangshan culture.
The Miaoshan site (28.92°N, 120.06°E) is also an early occupation of the Shangshan culture. The Miaoshan site has eight stratigraphic layers, among which layers 8 to 4 contain Shangshan cultural materials (Jiang et al., 2018). A total of 66 soil samples were collected from TG1, including 53 samples at 2 cm intervals from layer 8 to 4 and 13 samples at 5 cm intervals from layer 3 to 2 (Figure 2). Two charcoal samples were collected from layer 8 and 6 for radiocarbon dating.
The Hehuashan site (29.04°N, 119.25°E) has six stratigraphic layers, all of which contain middle Shangshan cultural materials (Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2016). A total of 50 soil samples were collected from T2 at 2 cm intervals (Figure 2).
The Huxi site (28.87°N, 120.02°E) is a late occupation of the Shangshan culture (Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2016; Zheng et al., 2016). The Huxi site has eight stratigraphic layers, among which layers 8 to 4 contain Shangshan cultural materials. A total of 75 soil samples were collected from ST3, including 68 samples at 2 cm intervals from layer 8 to 4, seven samples at 5 cm intervals from layer 3 to 2, and one sample from layer 1 (Figure 2).
The Qiaotou site (29.29°N, 119.97°E) is a late occupation of the Shangshan culture (Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2016; Zheng et al., 2016). The Qiaotou site has eight stratigraphic layers, among which layers 8 to 4 contain Shangshan cultural materials. A total of 33 soil samples from T1817 were collected at 5 cm intervals from layer 8 to 4 (Figure 2). Two charcoal samples were collected from layer 6 and 8 for radiocarbon dating.
Radiocarbon dating
Two charcoal samples from the Miaoshan site and two charcoal samples from the Qiaotou site were sent to Beta Analytic Inc. Laboratory for radiocarbon dating for this study. All samples successfully yielded radiocarbon dates. In addition, 19 previously published radiocarbon dates from the Shangshan site, Hehuashan site and Huxi site were also compiled for analysis (Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2016; Zuo et al., 2017). All 23 of these dates were calibrated by OxCal 4.4 using the IntCal 20 atmospheric curve (Reimer et al., 2020). The Shangshan culture lasted from 10,000 to 8000 cal. BP at these five sites (Figure 3).
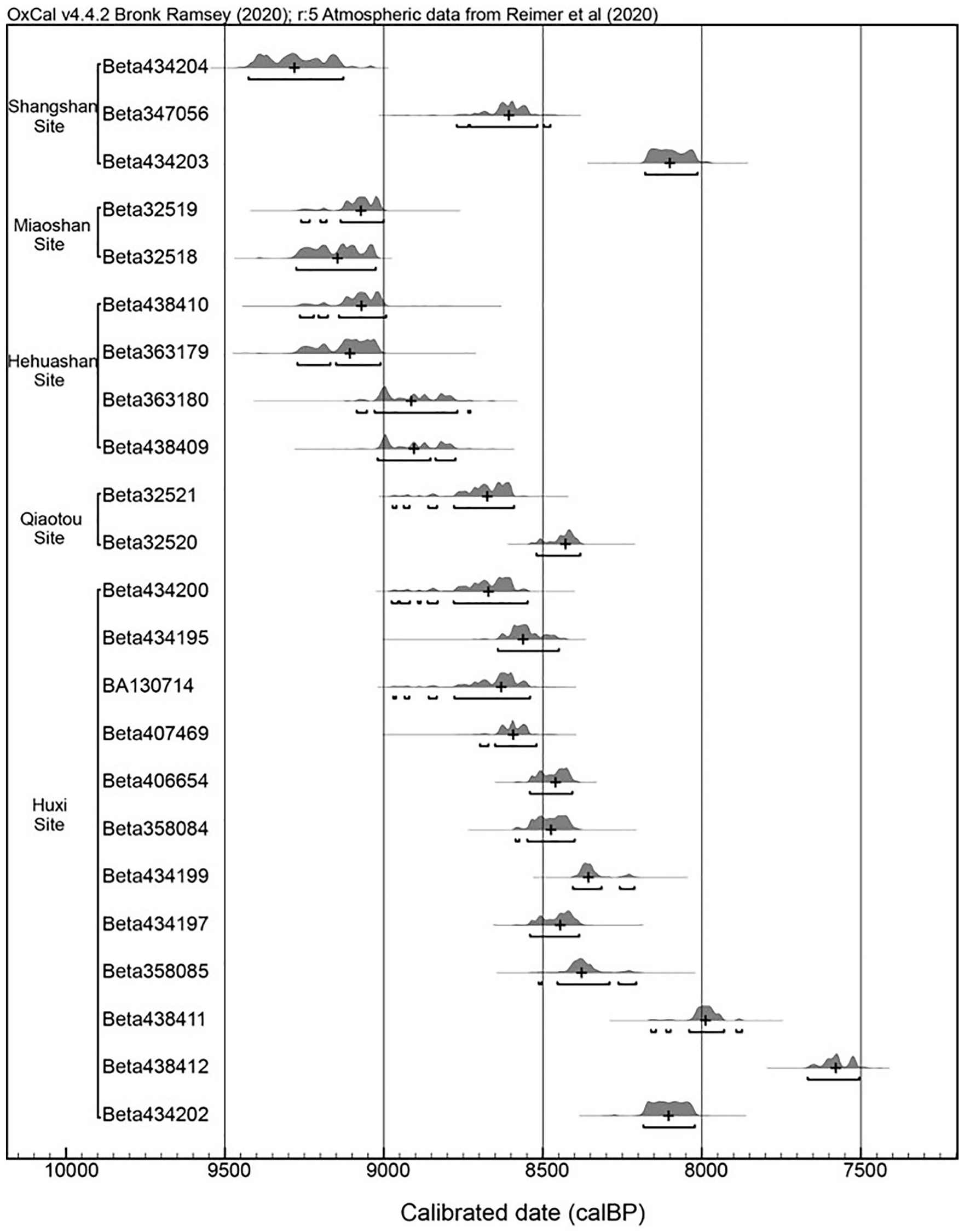
Calibrated radiocarbon dates from Shangshan culture sites.
Phytolith analysis
Phytoliths were extracted from the soil samples by the wet oxidation method. Initially, 2 g of soil samples was weighed. Subsequently, 30% hydrogen peroxide (H2O2) and 15% hydrochloric acid (HCl) were separately added to the samples to remove organic matter and carbonates. The samples were then subjected to heavy liquid flotation using ZnBr2 (density, 2.35 g/cm3) to separate the phytoliths, which were then mounted on a slide with Canada balsam.
The phytoliths were counted and identified under a Leica DM750 microscope at 400× magnification. More than 400 phytolith particles were counted in each sample. Phytolith abundance was expressed as the percentage of all phytoliths counted.
The bulliform phytolith selection criteria were based on Huan et al. (2015). For each rice bulliform phytolith, the number of fish-scale decorations around the edge was counted (Figure 4). Each sample was scanned until 50 rice bulliform phytoliths were encountered. In each case, the proportion of rice bulliform phytolith with ⩾9 fish-scale decorations was calculated to measure the rice domestication process.
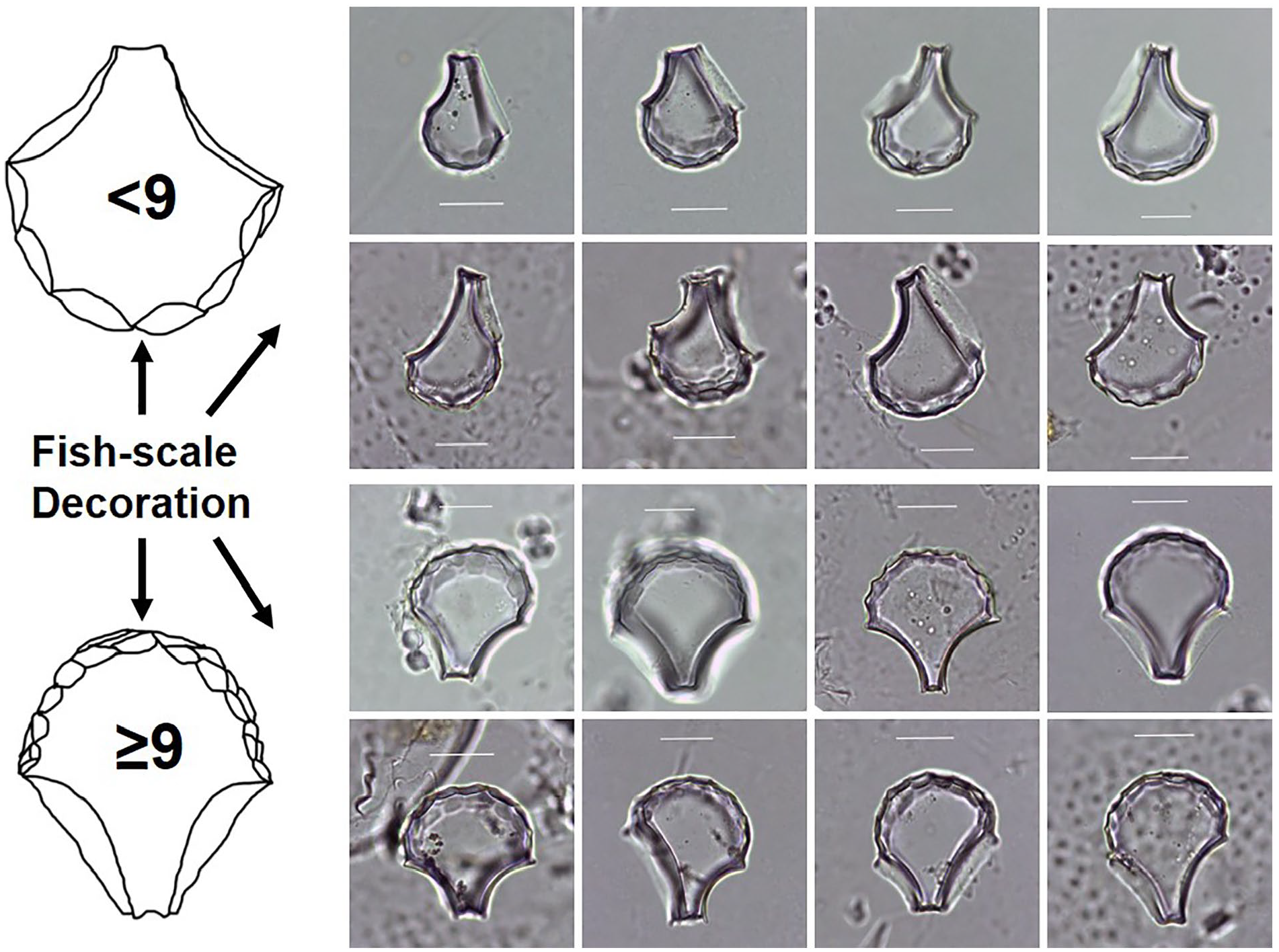
Fish-scale decorations on bulliform phytoliths from modern rice leaves.
Phytolith analysis results
All samples contain abundant phytoliths. Twenty-one phytolith morphotypes were identified, including long saddle, bilobate, rondel, trapeziform sinuate, elongate psilate, elongate echinate, square, rectangle, bulliform and others (Figure 5). Sufficient rice bulliform phytoliths with diagnostic domesticated and wild features were present at all five sites.

Major phytolith morphotypes in the five Shangshan cultural sites. (a and b) long saddle, (c and d) rondel, (e) multilobate, (f–i) bilobate, (j) parallel bilobate, (k) smooth elongate, (l) sinuate elongate, (m and n) trapeziform sinuate, (o) acicular hair cell, (p) square, (q) rectangle, (r) reed bulliform, (s) bulliform, (t) barnyard grass husk, (u and z) rice double-peaked, (v–y) rice bulliform with <9 fish-scale decorations, (aa–ad) rice bulliform ⩾9 fish-scale decorations.
At the Shangshan site, 10,944 phytoliths were identified from 24 samples for phytolith assemblage analysis. The phytolith assemblage (Figure 6) is characterized by a high proportion of long saddle (21%), followed by square and rectangle (19.7%), rondel (18.6%) and bilobate (11.8%); the percentage of rice bulliform phytoliths is 0.8%.
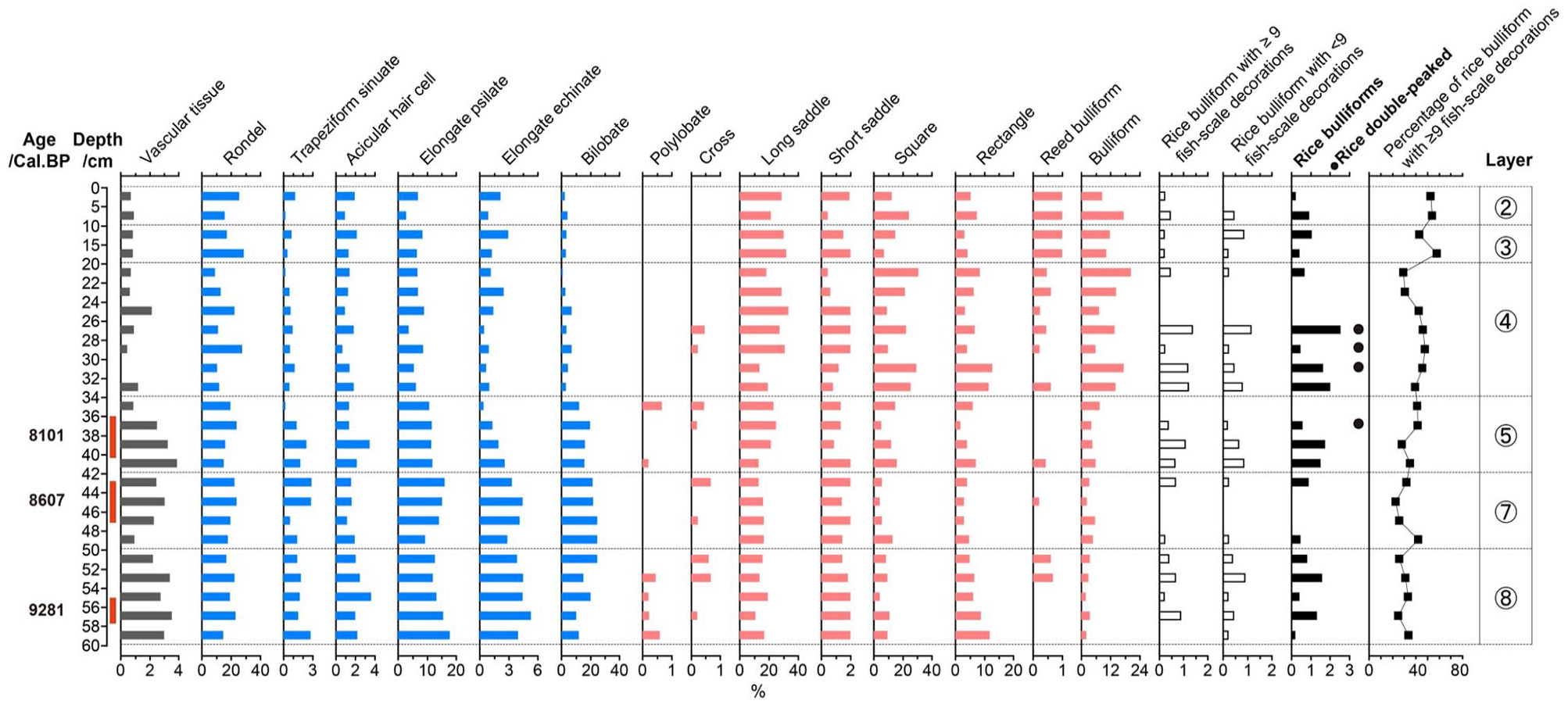
Selected phytolith percentage diagram of the Shangshan site.
A total of 1246 rice bulliform phytoliths were identified for the fish-scale decoration study. The proportion of rice bulliform phytoliths with ⩾9 fish-scale decorations showed an upward trend from early to late, except for a drop in layer 4. In addition, few rice double-peaked phytoliths appeared in four samples (SH12, SH15, SH16 and SH17). The results of phytolith dating in layer 8 and 5 were 9286 and 8101 cal. BP, respectively; the proportions of rice bulliform phytoliths with ⩾9 fish-scale decorations in these two layers are 25% and 42%; therefore, the relative domestication rate at the Shangshan site during Shangshan period was 1.4%/hundred yrs. The proportion of rice bulliform phytoliths with ⩾9 fish-scale decorations in layer 4 (Hemudu-Majiabang cultural period) was 40.4%.
At the Miaoshan site, 30,280 phytoliths were identified from 66 samples for phytolith assemblage analysis. The phytolith assemblage (Figure 7) was characterized by a high proportion of elongated psilate (25.5%), followed by square and rectangle (21.1%) and rondel (15.1%), and the percentage of rice bulliform was 2.1%. Rice double-peaked (0.59%) and parallel bilobate phytoliths were also found in the assemblage.
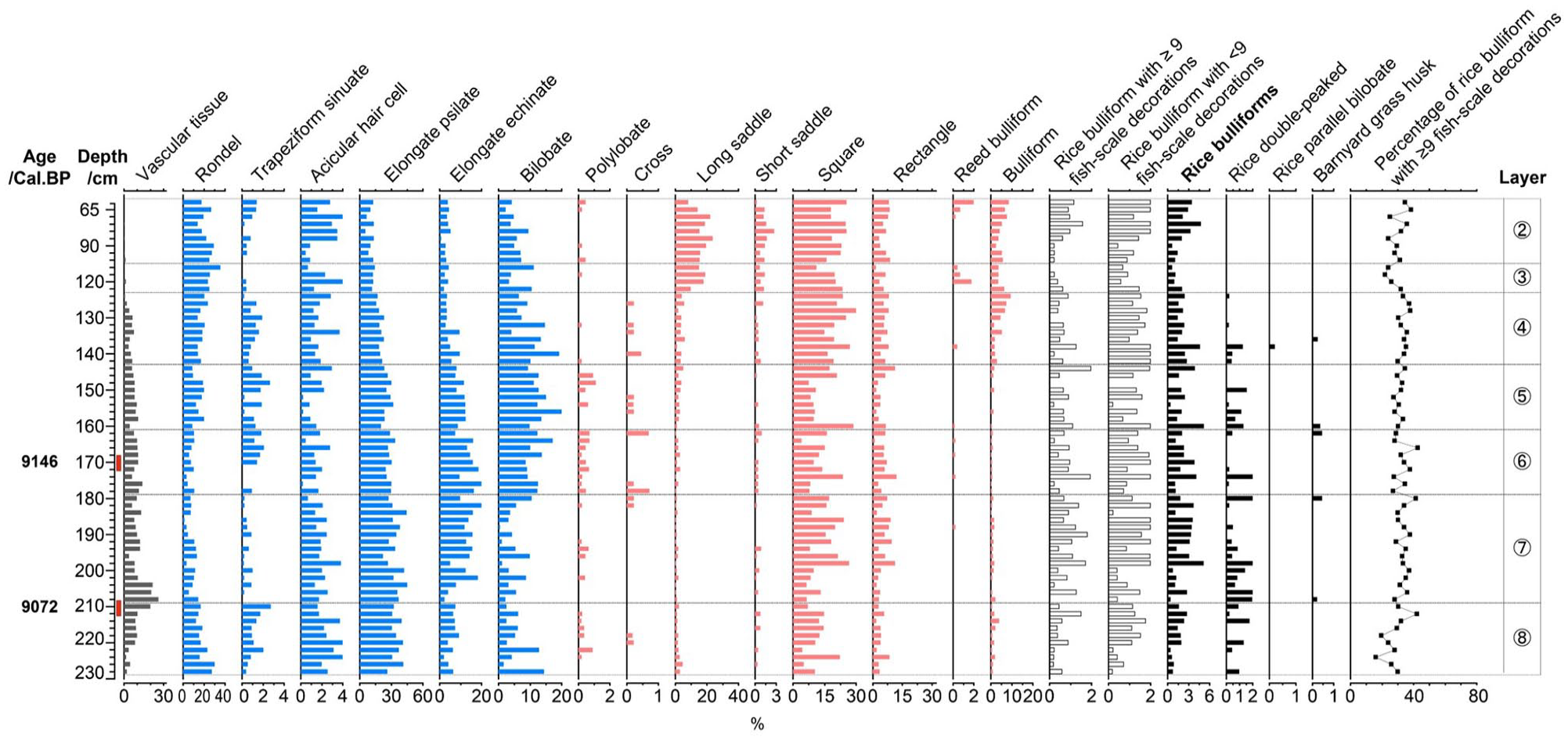
Selected phytolith percentage diagram of the Miaoshan site.
A total of 3434 rice bulliform phytoliths were identified for the fish-scale decoration study. According to the phytolith dating results and proportions of bulliform phytolith with ⩾9 fish-scale decorations from layer 8 (9072 cal. BP; 30.36%) and layer 6 (9146 cal. BP; 34%), so the proportion of rice bulliform phytoliths with ⩾9 fish-scale decorations remained steady from the early period to the late period.
At the Hehuashan site, 22,232 phytoliths were identified from 50 samples for phytolith assemblage analysis. The phytolith assemblage (Figure 8) was characterized by a high proportion of elongated psilate (25.0%), followed by rondel (23.1%) and square and rectangle (15.5%); the percentage of rice bulliform phytoliths was 1.5% and that of rice double-peaked phytoliths was 0.1%.
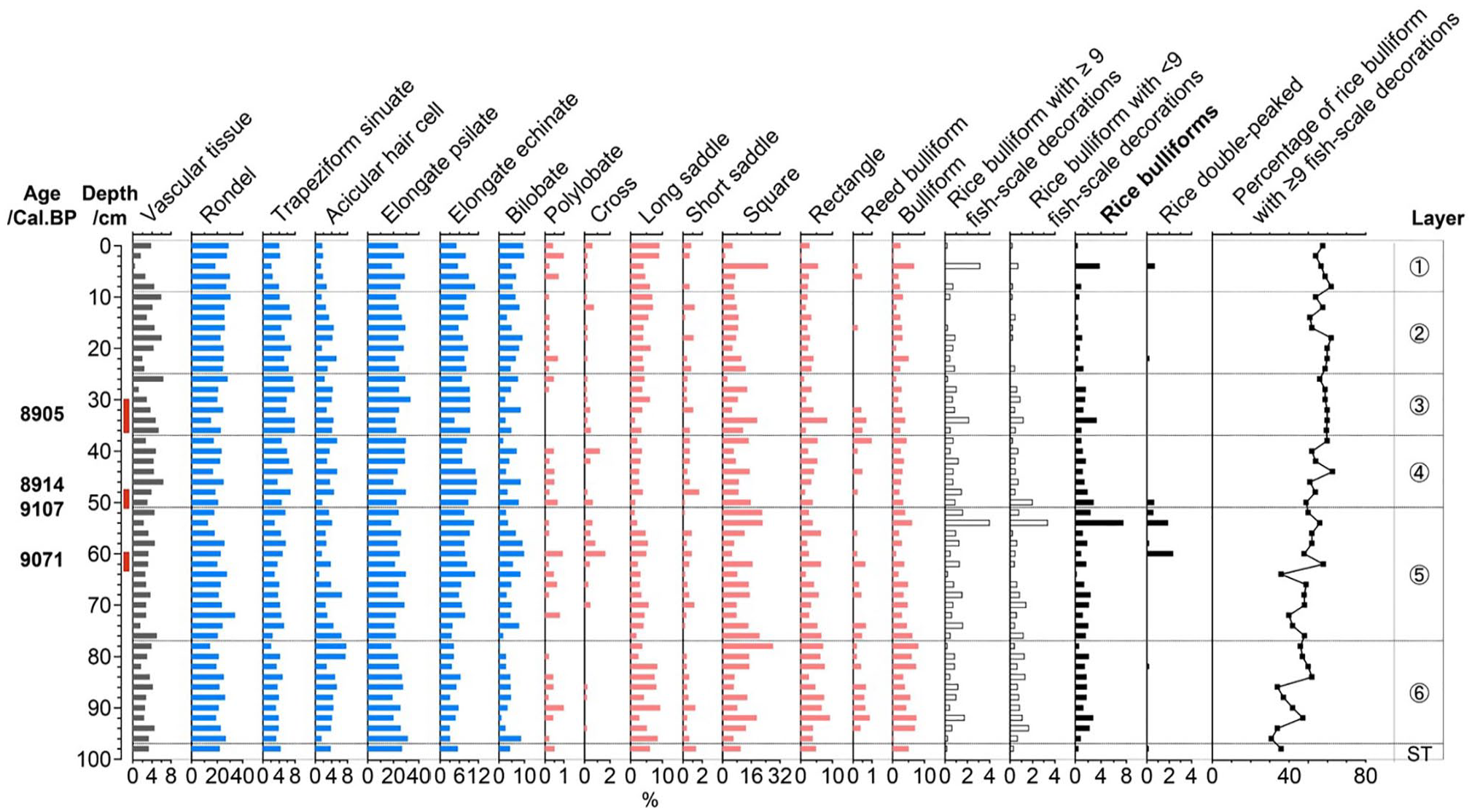
Selected phytolith percentage diagram of the Hehuashan site.
A total of 2550 rice bulliform phytoliths were identified for the fish-scale decoration study. The proportion of rice bulliform phytoliths with ⩾9 fish-scale decorations presented an increasing trend from the early to late phases. In addition, we also found rice bulliform phytoliths in the transition layer from cultural layer to natural deposit, and approximately 36% of these bulliform phytoliths with ⩾9 fish-scale decorations. According to the phytolith dating results and proportions of bulliform phytolith with ⩾9 fish-scale decorations from layer 5 (9071 cal. BP; 53%) and layer 3 (8905 cal. BP; 60%), the relative domestication rate at the Hehuashan site was 4.2%/hundred yrs.
At the Huxi site, 34,419 phytoliths were identified from 75 samples for phytolith assemblage analysis. The phytolith assemblage (Figure 9) was characterized by a high proportion of elongated psilate (29.4%), followed by square and rectangle (16.8%), rondel (16.6%) and bilobate (11.7%); the percentage of rice bulliform phytoliths was 2.2%, and that of rice double-peaked phytoliths was 1.43%.
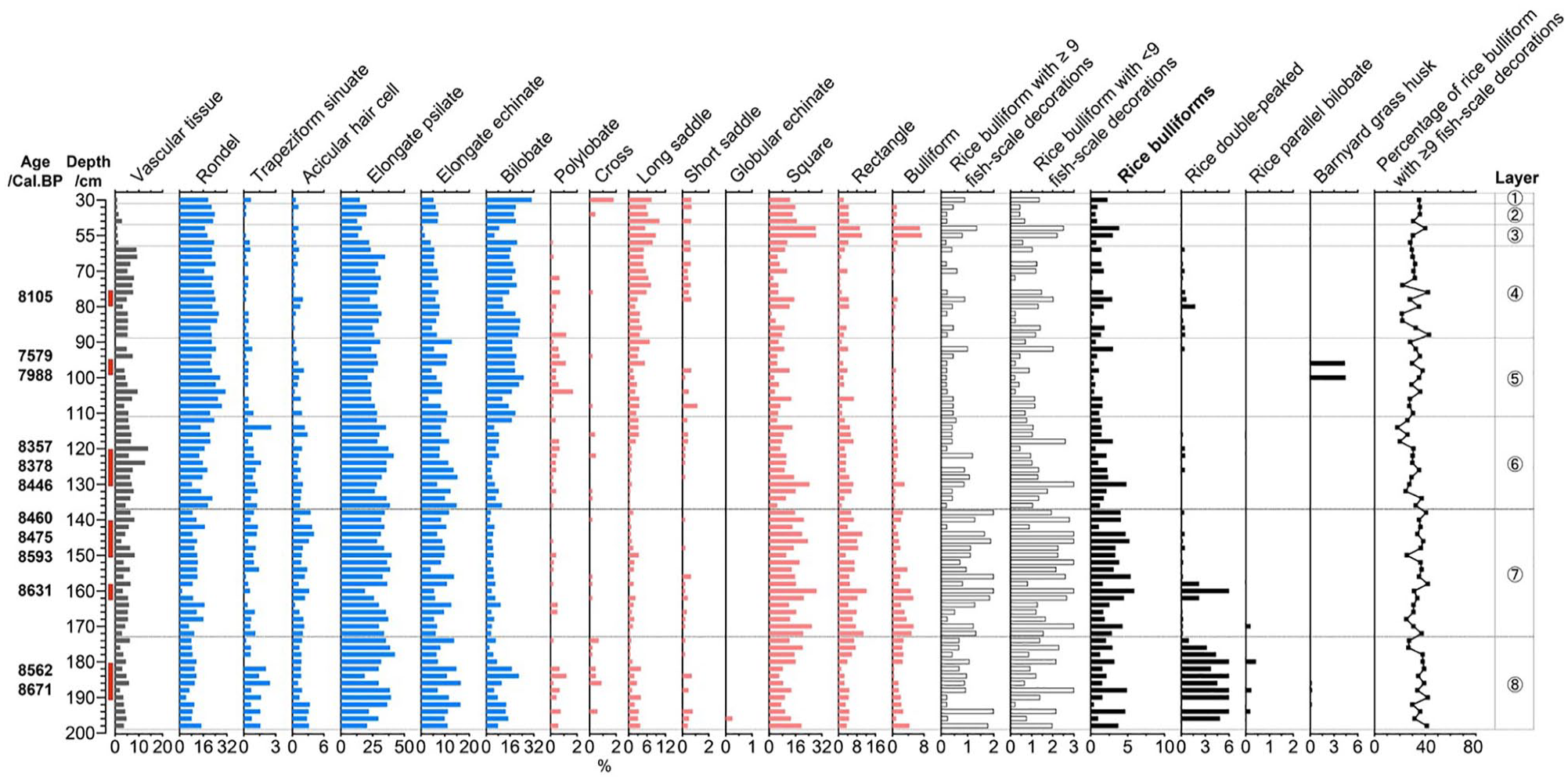
Selected phytolith percentage diagram of the Huxi site.
A total of 3937 rice bulliform phytoliths were identified for the fish-scale decoration study. The proportion of rice bulliform phytoliths with ⩾9 fish-scale decorations first decreased and then increased, but the overall change was not significant. The decorations of rice bulliform phytoliths and abundant rice double-peaked phytoliths in layer 8 were counted. According to the phytolith dating results and proportions of bulliform phytolith with ⩾9 fish-scale decorations from layer 8 (8671 cal. BP; 37.9%), layer 6 (8357 cal. BP; 29.2%) and layer 4 (8105 cal. BP; 32.1%); the domestication degree decreases from layer 8 to 6, and the relative domestication rate from layer 6 to 4 was 1.15%/hundred yrs.
At the Qiaotou site, 14,612 phytoliths were identified from 33 samples for phytolith assemblage analysis. The phytolith assemblage (Figure 10 ) was characterized by a high proportion of elongated psilate (32.9%), followed by square and rectangle (17.3%), rondel (14.8%) and bilobate (9.8%); the percentage of rice bulliform phytoliths was 2.1%, and that of double-peaked phytoliths was 0.4%.
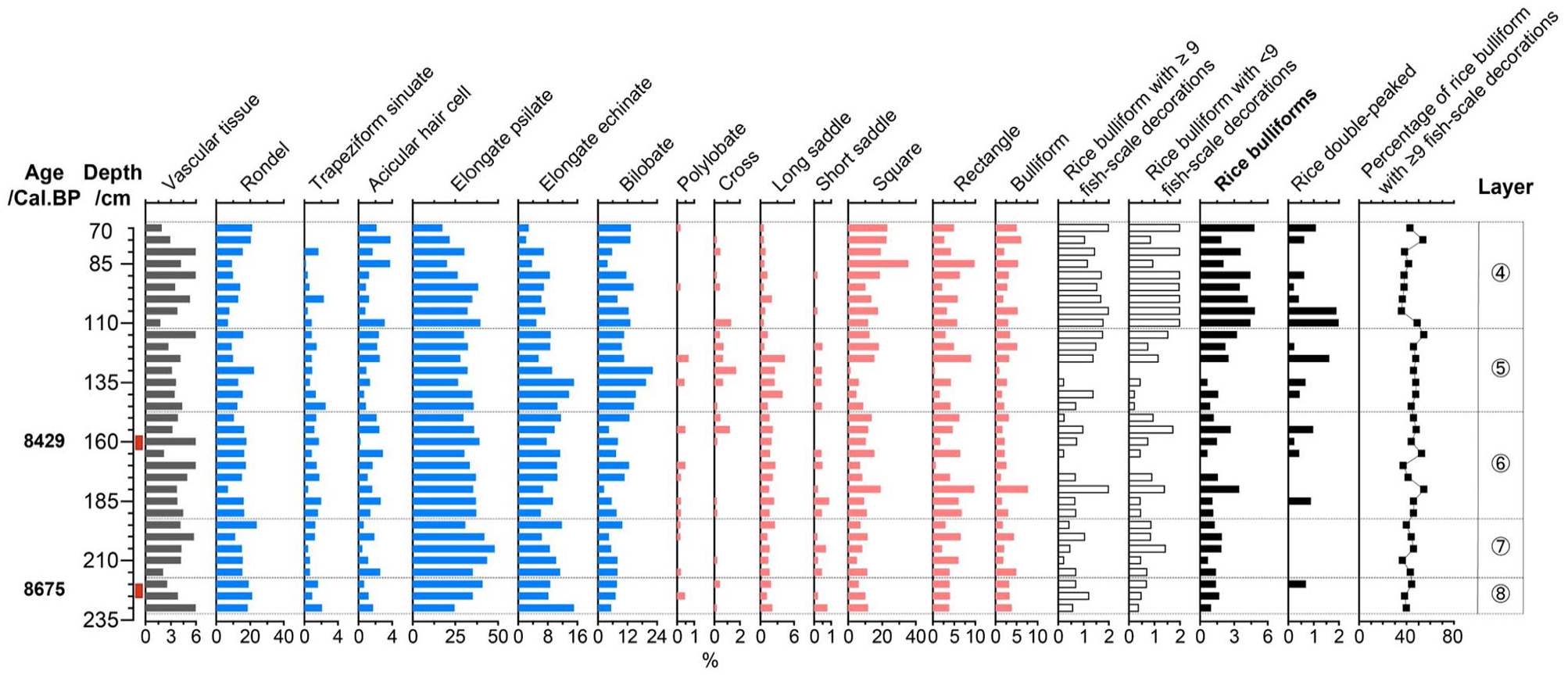
Selected phytolith percentage diagram of the Qiaotou site.
A total of 1703 rice bulliform phytoliths were identified for the fish-scale decoration study. The proportion of rice bulliform phytoliths with ⩾9 fish-scale decorations showed a slight increase and then a decreasing trend. While counting the decorations of rice bulliform phytoliths, rice double-peaked phytoliths were present in all layers. According to the charcoal dating results and proportions of bulliform phytolith with ⩾9 fish-scale decorations from layer 8 (8675 cal. BP; 38.46%) and layer 6 (8429 cal. BP; 44.23%), the relative domestication rate at the Qiaotou site was 2.3%/hundred yrs.
In summary, the proportions of rice bulliform phytoliths with ⩾9 fish-scale decorations in the five Shangshan archaeological sites are always higher than those in wild rice, and the rice domestication process appears to have differed among the five sites. Specifically, rice domestication at the Shangshan site presented a process of slowly increasing during the Shangshan culture period, decreasing during the Hemudu culture period, and then rapidly increasing. The degree of rice domestication at the Miaoshan site during the Shangshan culture period was generally low and stable, and there was a decrease until the end of the Shangshan culture. The degree of rice domestication at the Hehuashan site slowly increased and stabilized after layer 3. The degree of rice domestication at the Huxi site showed a trend of first decreasing, then increasing and then stabilizing during the Shangshan culture period and remained at a low-level stable state after the Shangshan period. The process of rice domestication at the Qiaotou site during the Shangshan culture period showed a process of first increasing and then decreasing.
Discussion
The domestication traits reflected by phytoliths from Shangshan rice remains
The Shangshan culture is the earliest known Neolithic culture in the lower Yangtze region (Zhejiang Provincial Institute of Archaeology and Pujiang Museum, 2007). However, the domestication traits of rice remain ambiguous throughout the Shangshan culture. The flotation results cannot provide systemic evidence of rice domestication due to the lack of rice remains in the cultural layers.
Abundant rice bulliform phytoliths were discovered in the Shangshan culture layers at all five sites. Through a systematic study of rice bulliform phytoliths, we found that the proportion of phytoliths in domesticated rise (rice bulliform phytoliths with ⩾9 fish-scale decorations) was always higher than that in wild rice in all early Shangshan culture samples. This result indicates that the rice domestication process began in the early stage of the Shangshan culture and continued through the middle and late stages of the Shangshan culture. In addition, many rice husks and rice leaves are present as tempers in the pottery sherds excavated at the Shangshan site (Zheng and Jiang, 2007); The results of microtrace analysis and phytolith analysis of stone tools unearthed at the Shangshan site also indicate that some of the stone tools were used to harvest grasses, including rice (Wang and Jiang, 2016), and harvesting crop with tools is a crucial method which could lead to increased domestication process (Hillman and Davies 1990).
Therefore, based on the evidence of rice bulliform phytoliths and archaeological excavations, rice was used not only as food but also as tempers in making pottery during the Shangshan culture, which means that rice was an important plant resource during the Shangshan period. As the main distribution area of the Shangshan culture, the lower Yangtze River region was likely an important origin of domesticated rice.
Spatial and temporal pattern of rice domestication in the Lower Yangtze Region
By comprehensively analysing the rice domestication process and dating results of the five sites (Figure 11), we found that evidence from early occupation of the Shangshan site and Miaoshan site indicated that the degree of rice domestication before 9 ka BP was approximately 35%; evidence from the Hehuashan site, Huxi site and Qiaotou site indicated that the degree of rice domestication from 9 to 8.5 ka BP had increased and was close to the domestication level; evidence from the Huxi site, Qiaotou site and Shangshan site indicated that the rice domestication degree from 8.5 to 8 ka BP was lower than that in the previous period but was still higher than that before 9 ka. In summary, during the early Holocene before 8 ka, the rice domestication process can be divided into three stages, characterized by a low level of domestication (10–9 ka BP), a high level of domestication (9–8.5 ka BP), and a fluctuating level of domestication (8.5–8 ka BP).
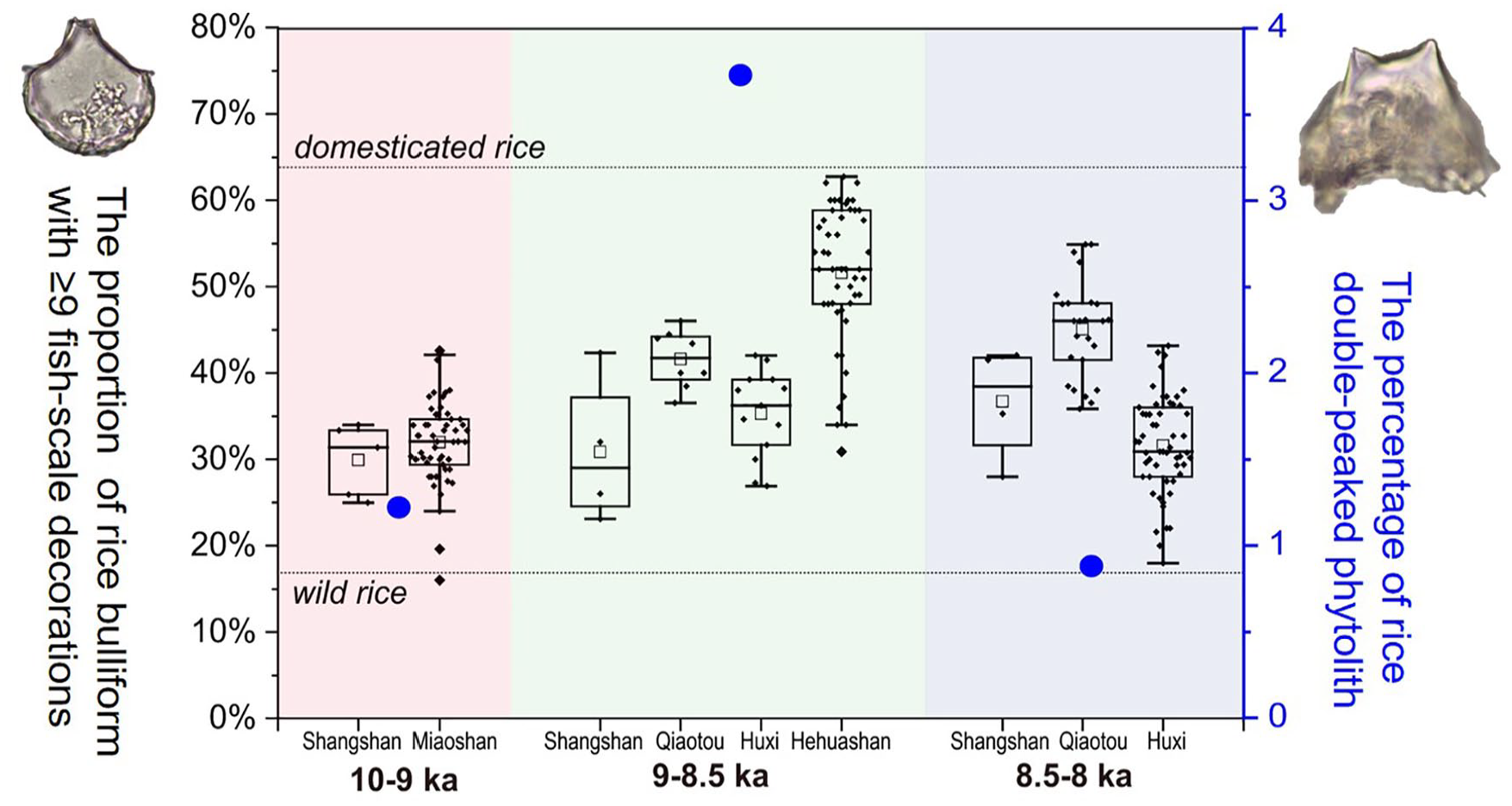
The rice domestication process during the early Holocene at the Shangshan cultural sites reflected by bulliform phytoliths (black box) and double-peaked phytoliths (blue dot).
During the early stage of rice domestication before 9 ka BP, when rice domestication was at a low level, rice double-peaked phytoliths, which come from rice husks, did not appear until approximately 9.2 ka BP, and even then, they appeared at only one site, Miaoshan (Figure 7). They then appeared at almost all layers of the Hehuashan site at approximately 9 ka (Figure 8). Eventually, they were widespread at the Huxi site, Qiaotou site and Shangshan site during 8.6–8 ka BP (Figures 6, 9 and 10). Double-peaked phytoliths can be left at sites during dehusking process of rice (Harvey and Fuller, 2005); therefore, the relatively higher proportion of double-peaked phytoliths in archaeological sites could be regarded as an indicator of crop processing.
Combining the rice domestication process reconstructed by bulliform phytoliths and the appearance time of rice double-peaked phytoliths, it is shown that although agriculture had already appeared, it was unstable during the Shangshan cultural period, as evidenced by the discontinuity in the presence of rice husks (Figure 6) in the early and middle phases of the Shangshan site. The continuous presence of rice husks was observed at the Qiaotou site, where a moat was discovered, indicating that the rice dehusking process consistently proceedin a higher hierarchically organized settlement.
We further analysed the rice domestication rate from the five sites during the Shangshan cultural period and found that the rice domestication rate had obvious regional characteristics during the early Holocene. The Shangshan site, with the lowest relative domestication rate (0.25%/hundred years), is locate in a small marginal basin outside of the Jinqu Basin. The Huxi site (1.15%/hundred years) and Qiaotou site (2.3%/hundred years) are both located in the upper reaches of tributaries of the Qujiang River, while the Hehuashan site (4.2%/hundred years) is located in the Qujiang River basin (Xu et al., 2016), which is the main channel of the upper Qiantang River (Figure 1). Therefore, it is not difficult to discern that the sites with higher domestication rates were distributed closer to the main stream. This might be due to (1) the main river channel has abundant and stable water resources, which is conducive to the continuous cultivation of rice; (2) The main river has wider terraces, which can support human activities on a larger scale and in a deeper intensity.
The fluctuating rice domestication process in the third stage (8.5–8 ka BP) may have been influenced by regional environmental change. Previous studies have shown that the increase of bilobate phytoliths is indicative of possible dry climatic conditions in the Yangtze Delta (Zhang et al., 2020; Zuo et al., 2016). Although limitations exist in the use of phytoliths unearthed from archaeological sites to reconstruct ancient environments, the synchronous of rice domestication fluctuations and bilobate content variations (Figure 6, 9 and 10) suggested that environmental changes might have affected the process of rice domestication.
In summary, the spatial and temporal pattern of the rice domestication process in the lower Yangtze River during the early Holocene was reconstructed by phytolith research. Before 9 ka BP, the degree of rice domestication was low and the sites were mainly distributed in the tributary areas of the mainstream rivers. After 9 ka, rice domestication rate accelerated and rice husks became widespread, meanwhile the sites expanded to near the main stream of the rivers. During 8.5–8 ka, rice domestication fluctuated which might have been caused by environmental changes.
Conclusions
Through systematic analysis of the phytoliths of five typical sites of the Shangshan cultural period, combined with accurate phytolith and charcoal dating results, the spatial and temporal pattern of the rice domestication process in the lower Yangtze River region during the early Holocene was reconstructed. Rice domestication process was begun across the region by the early stage of the Holocene and varied in space and time: In spatial, the sites with higher domestication rates were distributed closer to the mainstream river; In temporal, the early rice domestication process featured by low-high-fluctuating pattern; Besides, the crop dehusking process might have appeared by 9 ka in the lower Yangtze region.
Footnotes
Acknowledgements
The authors are grateful to the two anonymous reviewers for their helpful comments and suggestions.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Natural Science Foundation of China, Grant Nos. 41830322, 41907372; the Strategic Priority Research Program of Chinese Academy of Sciences, Grant No. XDB26000000.