
Abstract
Previous studies have found that individuals with autism spectrum disorders show impairments in mentalizing processes and aberrant brain activity compared with typically developing participants. However, the findings are mainly from male participants and the aberrant effects in autism spectrum disorder females and sex differences are still unclear. To address these issues, this study analyzed intrinsic functional connectivity of mentalizing regions using resting-state functional magnetic resonance imaging data of 48 autism spectrum disorder males and females and 48 typically developing participants in autism brain imaging data exchange. Whole-brain analyses showed that autism spectrum disorder males had hyperconnectivity in functional connectivity of the bilateral temporal-parietal junction, whereas autism spectrum disorder females showed hypoconnectivity in functional connectivity of the medial prefrontal cortex, precuneus, and right temporal-parietal junction. Interaction between sex and autism was found in both short- and long-distance functional connectivity effects, confirming that autism spectrum disorder males showed overconnectivity, while autism spectrum disorder females showed underconnectivity. Furthermore, a regression analysis revealed that in autism spectrum disorder, males and females demonstrated different relations between the functional connectivity effects of the mentalizing regions and the core autism spectrum disorder deficits. These results suggest sex differences in the mentalizing network in autism spectrum disorder individuals. Future work is needed to examine how sex interacts with other factors such as age and the sex differences during mentalizing task performance.
Keywords
Introduction
A key character of autism spectrum disorders (ASD) is the persistent deficits in social communication and social interaction across multiple contexts, which cause significant impairment in social functioning (Diagnostic and Statistical Manual of Mental Disorders (5th ed.; DSM-V), American Psychiatric Association (APA), 2013). It has been shown that the deficits may be related to the dysfunction of the mentalizing system that plays an important role in understanding others’ intentions and goals (Fishman et al., 2014; Libero et al., 2014; Marsh and Hamilton, 2011).
Previous studies found that the mentalizing system includes several regions in the anterior and posterior parts of the brain, including medial prefrontal cortex (mPFC), temporal-parietal junction (TPJ), precuneus, and posterior cingulate cortex (PCC) (Fletcher et al., 1995; Van Overwalle and Baetens, 2009; Vogeley et al., 2001). The mentalizing system is involved in a broad range of tasks that require the understanding of others’ mental state, action goals, beliefs, personality traits, and emotions (for reviews, see Van Overwalle, 2011; Van Overwalle and Baetens, 2009).
In autism research, studies have shown that individuals with ASD demonstrated aberrant activation effects in the mentalizing regions during imitation and observation tasks compared with typically developing (TD) individuals (Dapretto et al., 2006; Jack and Morris, 2014; Libero et al., 2014; Marsh and Hamilton, 2011; Martineau et al., 2010; Poulin-Lord et al., 2014). For instance, Marsh and Hamilton (2011) found that in a hand action observation task, rational actions elicited stronger activity in the bilateral mPFC than irrational actions only in the TD group. In addition, behavioral assessment of mentalizing and gesture recognition showed that the ASD group was less able to select appropriate mental state description than the TD group, and that the ASD group made more errors and had longer reaction time (RT) than the TD group in the gesture recognition task. Martineau et al. (2010) indicated that hand action observation elicited stronger activity in the bilateral precuneus in TD participants only. Poulin-Lord et al. (2014) revealed that during hand action imitation, TD individuals showed stronger activation than ASD individuals in the bilateral TPJ, whereas ASD individuals showed stronger activity in the bilateral precuneus and right mPFC. Freitag et al. (2008) found that in a biological motion observation task, the TD group had strong activation in the left PCC, whereas the ASD group had strong activity in the bilateral PCC and left precuneus. Additionally, in a separate behavioral assessment of imitation and adaptive motor abilities, the ASD group demonstrated longer RT in recognizing biological motion as well as scrambled motion and weaker imitation ability than the TD group.
The above studies reported no behavioral measurements during their action observation or imitation tasks in functional magnetic resonance imaging (fMRI), which might be due to the difficulty in analyzing the data (e.g. imitation performance) or the potential confounding effect of motor activity from button response. However, some studies conducted additional behavioral tests to examine basic action imitation and observation skills in ASD and TD groups (Freitag et al., 2008; Marsh and Hamilton, 2011), and they found that the ASD group was less capable in these skills. These behavioral results and brain activation findings together suggest that the mentalizing system is involved in imitation and observation, and that ASD individuals might have dysfunctions of the system.
Researchers using other tasks that focus on judgment or comprehension of social behavior also found different brain effects in ASD individuals and TD individuals (Carter et al., 2012; Kana et al., 2015; Pantelis et al., 2015). In Carter et al. (2012), ASD and TD children were asked to judge in which of the two pictures a boy’s behavior was bad (social condition) or in which of the two pictures an action took place outdoors (physical condition). The fMRI results showed that TD participants had greater neural activity in the mPFC, bilateral inferior frontal gyrus (IFG), and left superior temporal pole (TP) than children with ASD. In Kana et al. (2015), participants were presented with animations, and answered questions about the behavior of the figures in the animations. In a theory of mind (ToM) condition (figures had an interaction involving feelings and thoughts, e.g. coaxing), ASD participants had significantly less neural activity in the mPFC, the mirror network, and the cerebellum than TD participants. In addition, in ASD participants, there was underconnectivity between frontal and posterior regions during task performance. In a study about assessing the awkwardness of different social interactions, Pantelis et al. (2015) found that when watching videos that contain many socially awkward moments, both ASD and TD participants had activity in the mentalizing regions, but ASD participants had significantly decreased activation round the right TPJ, which extended into the posterior end of the right superior temporal sulcus (STS). The authors suggest that the right TPJ and right STS regions have a critical role in navigating social situations.
Although the above studies showed that ASD individuals had aberrant activity in the mentalizing network compared with TD participants, the findings are mainly from ASD males, and the sex differences in the aberrant effects of the mentalizing network are still unclear. This is because the prevalence of ASD in males is much higher than that in females (Newschaffer et al., 2007), and studies often recruited ASD male participants or mixed groups with few ASD female participants. Only recently, some studies started to focus on sex differences in the neurology involved in social understanding (Kirkovski et al., 2015; Schneider et al., 2013). For instance, Kirkovski et al. (2015) found no group differences in neural activity in the mPFC or TPJ during a ToM task. However, males with ASD had less neural activity in the posterior STS than TD males when processing social information. This difference was absent in TD females and females with ASD. The authors suggest sex differences in the activity of the posterior STS in ASD. Schneider et al. (2013) explored the empathy abilities in ASD males and ASD females. Participants watched story videos and judged their own emotions and the actor’s emotions. The fMRI results showed that ASD males had stronger activity in the bilateral mPFC than ASD females, and that ASD females had decreased activation in the midbrain and limbic regions compared with TD females. The authors suggest that ASD males and females may have different ways to process empathy: ASD males may have impairments in cognitive aspects of empathy, whereas ASD females may have alternations of social reciprocity.
In this study, we investigated the sex differences in the brain network of the mentalizing system in ASD individuals, which still lacks evidence. We used resting-state fMRI technique to study the intrinsic functional connectivity (FC) of the mentalizing system in ASD. We chose this approach to examine the mentalizing networks based on advantages of this approach and disadvantages of task-state fMRI. First, mentalizing, which requires understanding the mental state of oneself or others, is a complex process and its activation can be modulated by task demands. So far, researchers have used various tasks to study mentalizing networks, such as story comprehension (e.g. Ciaramidaro et al., 2007; Walter et al., 2004); virtual taxi driving (Spiers and Maguire, 2006); passive viewing films or animations about goal-directed actions (Moriguchi et al., 2006); visual detection (Astafiev et al., 2006; Giessing et al., 2006); viewing goal-directed object motions (Martin and Weisberg, 2003; Schultz et al., 2004); ToM belief tasks (Abraham et al., 2008; Mitchell, 2008); and morality judgment (Borg et al., 2006; Moll et al., 2002; Young and Saxe, 2008). Some tasks activate only part of mentalizing networks (e.g. right TPJ in visual detection tasks; bilateral TPJ; and mPFC in goal-directed object motion tasks), whereas other tasks may activate broader networks (e.g. ToM belief tasks and morality judgment tasks). Furthermore, within the same type of task, the variety of baseline conditions can cause differences in brain activity. To avoid these task influences, we decided to derive mentalizing regions from a meta-analysis study that quantitatively summarizes the activation of mentalizing networks in various tasks (Van Overwalle and Baetens, 2009) and used the regions as seeds in FC analysis on resting-state datasets. Second, FC is defined as the interregional cross correlations in the brain (Fox and Raichle, 2007), and even in the resting-state, FC patterns still correspond to the brain networks involved in specific cognitive tasks (Beckmann et al., 2005; Fox and Raichle, 2007; Simmons and Martin, 2012; Smith et al., 2009). It is suggested that the intrinsic FC may reflect the functional specialization of the brain, which is formed by frequent co-activation during cognitive processes (Fair et al., 2007, 2009). Third, in resting-state fMRI studies, the signal-to-noise ratio is better than task-state fMRI studies (Fox and Raichle, 2007).
This resting-state FC approach has been used in autism research to study the intrinsic brain network of the mentalizing system in ASD population (Fishman et al., 2014). In that study, the authors found that compared with TD participants, ASD participants showed both hypoconnectivity and hyperconnectivity in the mentalizing network, and the degree of the aberrant effects was related with greater social impairment. In addition, the mentalizing network had excessive cross talk with the mirror neuron system, which was also associated with more severe social dysfunction.
This study examined the intrinsic FC of the main mentalizing regions (bilateral TPJ, mPFC, and precuneus) in ASD males as well as females and in matched TD participants. The resting-state fMRI data were derived from the autism brain imaging data exchange (ABIDE, Di Martino et al., 2014). The database contains resting-state fMRI data collected from 539 individuals with ASD and 573 TD individuals, which include a number of female participants. Previous studies have used ABIDE to test the sex differences in the brain structure of ASD males and females (e.g. Schaer et al., 2015; Supekar and Menon, 2015). In this study, we selected mentalizing regions revealed by a previous quantitative meta-analysis (Van Overwalle and Baetens, 2009) and conducted seed-based FC analyses in each participant group.
It should be noted that the mentalizing network and the default mode network (DMN) have a strong overlap. Previous research has shown that the DMN is activated during higher-order tasks that attribute mental states to others (Mars et al., 2012; Schilbach et al., 2008). For instance, using meta-analysis, Laird et al. (2011) found that brain activity from social cognitive tasks showed brain effects in the bilateral TPJ, precuneus/PCC, and mPFC. These effects were very similar to the DMN network defined by independent component analysis (ICA; e.g. Smith et al., 2009). In a recent meta-analysis, Mars et al. (2012) found that at the level of the network and the level of individual brain regions, there are to be consistent overlap between the DMN areas and the areas in mentalizing tasks. Schilbach et al. (2008) suggested that the physiological “baseline” of the human brain (i.e. the DMN) is linked to the psychological “baseline” (i.e. the mentalizing network), which means that thinking about self and others may be the default mode of thought. In this study, we selected seed regions in the mentalizing network based on the meta-analysis of mentalizing tasks (Van Overwalle and Baetens, 2009), and we did not particularly focus on the DMN. Nevertheless, we agree that there would be a strong overlap between the DMN and the mentalizing network because mentalizing tasks require episodic memory and mental simulations, which are prone to rely on the DMN (Buckner et al., 2008). We suggest that future research should focus on the precise anatomy and functions of the two networks and their similarities and differences.
In this study, we predicted that compared with TD participants, ASD participants would demonstrate reduced FC of the mentalizing regions (Kana et al., 2014, 2015; Libero et al., 2014), and the aberrant effects would be related to the core ASD deficits. Furthermore, ASD females and ASD males might show different atypical FC effects (Schneider et al., 2013). One possibility was that ASD males would demonstrate more severe aberrant effects compared with ASD females because the prevalence of ASD is higher in males than in females (e.g. Kirkovski et al., 2015).
Methods
Participant selection
Initial selection
The data selection started from female datasets since there were much fewer female participants than male participants in ABIDE. We selected datasets in which the ASD participants were diagnosed in accordance with the Diagnostic and Statistical Manual of Mental Disorders (4th ed., DSM-IV-TR) classifications of autism disorder, Asperger’s syndrome, or pervasive developmental disorder not-otherwise-specified (PDD-NOS), and performed the review of Autism Diagnostic Interview-Revised (ADI-R, Lord et al., 1994) and Autism Diagnostic Observation Schedule (ADOS, Lord et al., 2000). In addition, both ASD and TD participants had intelligence quotient (IQ) scores (either full-scale IQ scores or verbal IQ and performance IQ scores). The initial selection found 147 participants.
Data acquisition
Scan procedure and parameters can be found in supplementary material (available online).
Data preprocessing
The Analyses of Functional NeuroImages (AFNI) statistical software package (Cox, 1996) was used to analyze the data. For the preprocess of the resting-state fMRI data, we removed spikes in each voxel’s time series, conducted slice timing alignment on volumes, aligned the echo planar imaging (EPI) to the anatomy, conducted volume registration, blurred each volume with 4 mm full-width half-maximum, and band-pass filtered the EPI data (0.009 Hz < f < 0.08 Hz, Dosenbach et al., 2010). Linear regression was conducted to regress out the estimated motion parameters. The cleaned data were resampled to 3 × 3 × 3 mm3 voxel size and aligned in the MNI-152 template.
It is important to note that the despiking method in the preprocessing does not involve scrubbing. Instead, the despiking method used Fourier fit and identified spikes by comparing residuals against the standard deviation (SD) of the residuals. Spike signals were remapped so that no large changes would be produced in the despiked output (Jo et al., 2013). It has been shown that this method may improve volume registration and make motion parameters less variable (Jo et al., 2013).
For the resting-state fMRI data, head motion can significantly influence the FC results (Goto et al., 2015). To reduce the influence of motion artifacts, we first discarded individual data in which the maximum distance of translational head movement was larger than 2 mm or the maximum degree of rotational head movement was larger than 2°. After that, we used the motion correction method implemented in the afni_proc.py program (http://afni.nimh.nih.gov/pub/dist/doc/program_help/afni_proc.py.html), which adopts a strict censor threshold (0.2) decided by taking the derivative of the motion parameters and the sqrt (sum squares) per TR. The TR would be censored if this Euclidean norm exceeds the given limit. We additionally censored TRs when more than 10% of the automasked brain were outliers because these TRs were typically due to the motion. Since motion is not generally a whole-brain-between-TRs event, these TRs might capture motion where the motion parameters do not. Although motion censoring might disrupt the temporal structure of the data, so far it is still the most effective way to ensure that the data were not influenced by spurious, motion-related noise (Power et al., 2014). In addition, scrubbing can effectively be used to minimize the motion-related artifacts in seed-based correlation analyses (Power et al., 2014).
We also want to mention that in this study, the band-pass filtering was performed during linear regression to allow us to perform motion censoring at the same time. Furthermore, it is suggested that simultaneous band-pass filtering and regression could be a better way to control noise in data and avoid reintroducing synchronous noise into the data (Hallquist et al., 2013).
Final sample selection
After the motion correction procedure, 42 participants with extreme head motion were excluded, and thus we had a final sample of 83 TD females and 24 ASD females. In all, 24 TD females whose age and full-scale IQ were matched with the ASD females were selected. Table 1 shows the demographic data of TD and ASD female participants. In the second step of participant selection, we selected ASD and TD male participants whose age and full-scale IQ were matched with ASD and TD female participants. First, we selected the imaging data of ASD and TD males from the same research institutes (University of Pittsburg, University of Michigan, Yale Child Study Center, Kennedy Krieger Institute, NYU Langone Medical Center, Stanford University, and University of California, Los Angeles) and preprocessed the data using the same steps. After head motion correction, we selected 24 ASD males and 24 TD males who were matched with the ASD and the TD females on age and IQ scores. Table 1 shows the demographic data of the included male participants.
Characteristics of the ASD and TD participants.
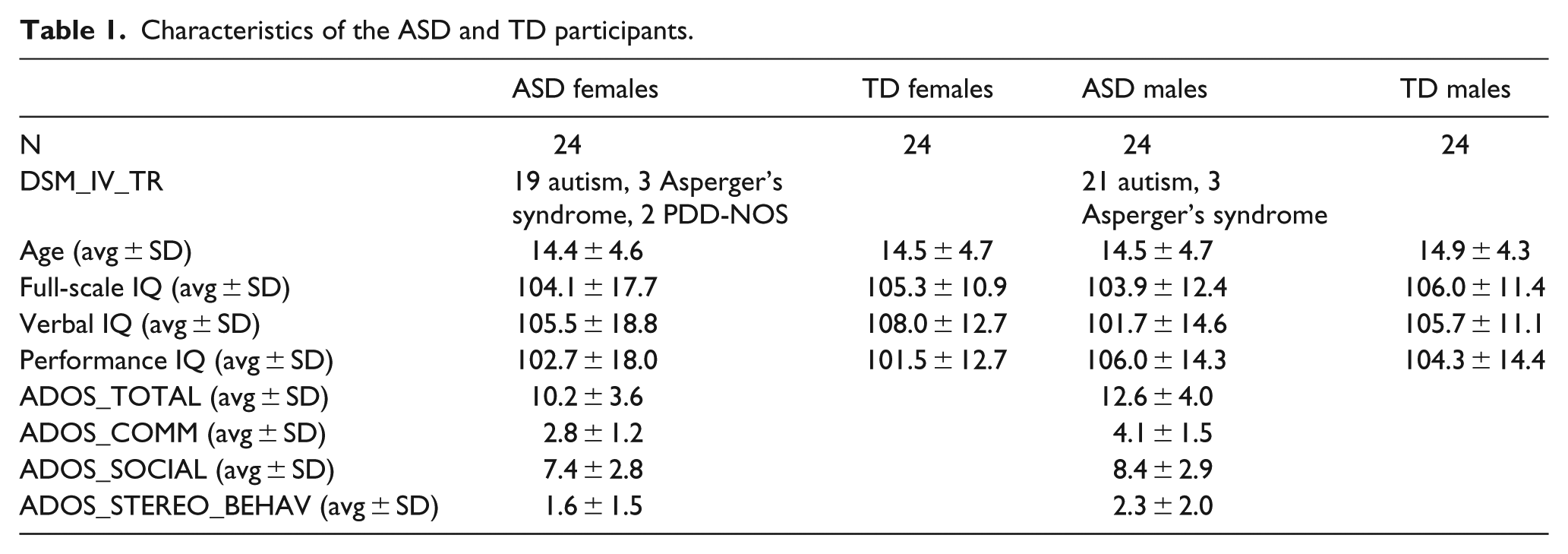
One-way analysis of variance (ANOVA) was conducted to examine the significant group differences in age and IQ. The results showed that no significant group effect was found for age (F(3, 92) = 0.056, p = 0.982), full-scale IQ (F(3, 92) = 0.134, p = 0.940), verbal IQ (F(3, 92) = 0.768, p = 0.515), or performance IQ (F(3, 92) = 0.409, p = 0.747).
FC analysis
The coordinates of seed regions in the mentalizing system were selected from Van Overwalle and Baetens (2009), a meta-analysis that investigated the mirror and mentalizing systems of understanding others’ action goals. Four mentalizing regions were selected, including the mPFC, bilateral TPJ, and precuneus (Fishman et al., 2014). All coordinates were in the MNI space: left TPJ (−52, −55, 30), mPFC (1, 57, 12), precuneus (2, −58, 46), and right TPJ (56, −54, 28).
In each participant, each seed region was created as 5-mm radius spheres with the coordinate as the center of the sphere. The average blood oxygen level–dependent (BOLD) time course was extracted from each seed region and then correlated with the time courses of all voxels in the rest of the brain (Pearson’s correlation). For each voxel-wise correlation, the r-map was generated and the r value was converted to z values using Fisher r-to-z transformation. False discovery rate (FDR) correction (p < 0.05) was used to correct the correlation results, and only significant effects were used in the group analysis.
Group analysis
To investigate the interaction between the factor sex (female vs male) and the factor autism (TD vs ASD), we conducted ANOVA computation for each seed region using a three-dimensional multivariate modeling (3DMVM) approach (Chen et al., 2014) with participants’ age, IQ scores, eye status during scanning, and site information (TR, voxel size, and length of scan) as covariate variables. The site information has been proved to influence fMRI analysis results (e.g. Birn et al., 2013; Hopfinger et al., 2000).
If significant interaction between sex and autism was found, we performed two-sample t-tests to examine the significant differences between ASD females and TD females and significant differences between ASD males and TD males. In addition, we performed one-sample t-tests to examine the significant FC effects in each participant group. In all t-test analyses, participants’ age, IQ, and site information were added as covariate variables.
For all whole-brain statistical analyses, we used a cluster-size threshold based on Monte Carlo simulations (3dClustSim, updated after December 2015). This method thresholding individual voxel at a specific p level uncorrected and resulted in a corrected threshold of p < 0.01. We chose p < 0.01 because there were totally four seeds (bilateral TPJ, mPFC, and precuneus) and thus the significantp value after the correction of multiple ANOVA testing (Poldrack et al., 2008) should be 0.05/4 = 0.0125. To achieve this p value, for all one-sample t-tests, the voxel-wise p value was p < 0.001 (uncorrected), and the cluster-size threshold was voxel number >18. For the F tests and the two-sample t-tests that examine the group differences, the voxel-wise threshold was p < 0.005 (uncorrected) and the cluster-size threshold was voxel number >24.
Region of interest analysis
We conducted region of interest (ROI) analyses to further examine the regional sex × autism interaction effects from ANOVA. For each ROI, the coordinate was the peak F value coordinate of the interaction effect. A 5-mm radius sphere around the peak F value coordinate was drawn and the mean FC in each participant was calculated. After that, ANOVA was conducted to test the interaction effect in the ROI. Significant effects were reported at p < 0.05.
Relation between ASD deficits and FC effects
To see the sex differences in the relation between ASD symptoms and FC, we examined the relation between autism deficits and FC effects in females and males with ASD. To achieve the goal, ADOS scores (ADOS_COMM, ADOS_SOCIAL, and ADOS_STEREO_BEHAV) were added into the general linear model as covariates for the FC effects of each seed region (regression analyses) in ASD female and male participants. A cluster-size threshold based on Monte Carlo simulation (p < 0.05) was used. At the voxel-level, the threshold was p < 0.005 (uncorrected) and the cluster-size threshold was voxel number >20.
Results
For each seed region, the FC effect in each participant group was shown in supplementary Figures 1 to 4 (available online).
Effects of sex and autism
The mentalizing region: left TPJ
For the FC effect of the left TPJ, ANOVA revealed a main effect for sex in the bilateral middle frontal gyrus (MFG), left superior medial gyrus, left supramarginal gyrus (SMG)/angular gyrus (AG), left precuneus, left TP and left IFG (p. orbitalis). No main effect for autism was found. Furthermore, a sex × autism interaction was revealed in the left inferior parietal lobule (IPL) and left supplementary motor area (SMA). Within the male groups, the whole-brain contrast showed that ASD males had increased FC in the left superior frontal gyrus (SFG)/MFG (Figure 1(a)). No significant group effect was found in the female participants.
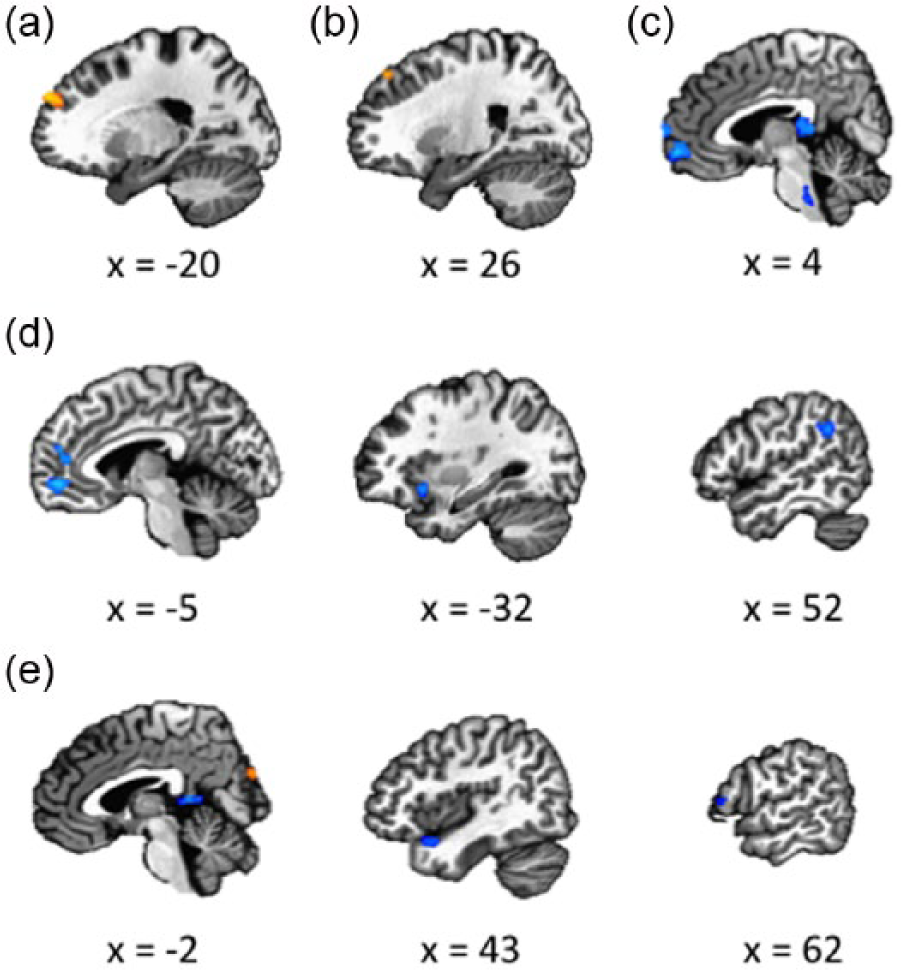
Results of whole-brain contrast analyses of the FC in females and males. (a) Seed region left TPJ: ASD males versus TD males. (b) Seed region right TPJ: ASD males versus TD males. (c) Seed region right TPJ: ASD females versus TD females. (d) Seed region mPFC: ASD females versus TD females. (e) Seed region precuneus: ASD females versus TD females.
The mentalizing region: right TPJ
For the FC effect of the right TPJ, ANOVA revealed a main effect for Sex in the left AG. No main effect for Autism was found. Furthermore, a sex × autism interaction was revealed in the left PCC. In the male participants, ASD males showed increased FC in the left SFG compared with TD males (Figure 1(b)). Within the female groups, the whole-brain contrast indicated that ASD females showed decreased FC in the PCC, right mid orbital gyrus, and right superior medial gyrus (Figure 1(c)).
The mentalizing region: mPFC
For the FC effect of the left mPFC, ANOVA revealed a main effect for sex in the left mid orbital gyrus, left AG, left PCC, left anterior cingulate cortex (ACC), and left IFG (p. triangularis). No main effect for autism was found. Furthermore, a sex × autism interaction was revealed in the bilateral SFG, left superior medial gyrus, left PCC/thalamus, left middle cingulate cortex (MCC), and right AG. Within the female groups, ASD females showed decreased FC in the left superior medial gyrus, left ACC, right AG, and left insula (Figure 1(d)). No significant group effect was found in the male participants.
The mentalizing region: precuneus
For the FC effect of the precuneus, ANOVA revealed a main effect for sex in the left middle occipital gyrus (MOG), left precuneus, left superior occipital gyrus (SOG)/MOG, right ACC, left postcentral gyrus, and left SMA. No main effect for autism was found. Furthermore, A sex × autism interaction was revealed in the right SOG, right IFG (p. triangularis), and left cuneus. Within the female groups, the whole-brain contrast analysis revealed that ASD females showed decreased FC in the left IFG (p. opercularis), left precuneus, and right TP. In addition, ASD females showed increased FC in the left cuneus and right SOG (Figure 1(e)). No significant group effect was found in the male participants.
Table 2 shows the results of ANOVA.
FC effects of each seed region.
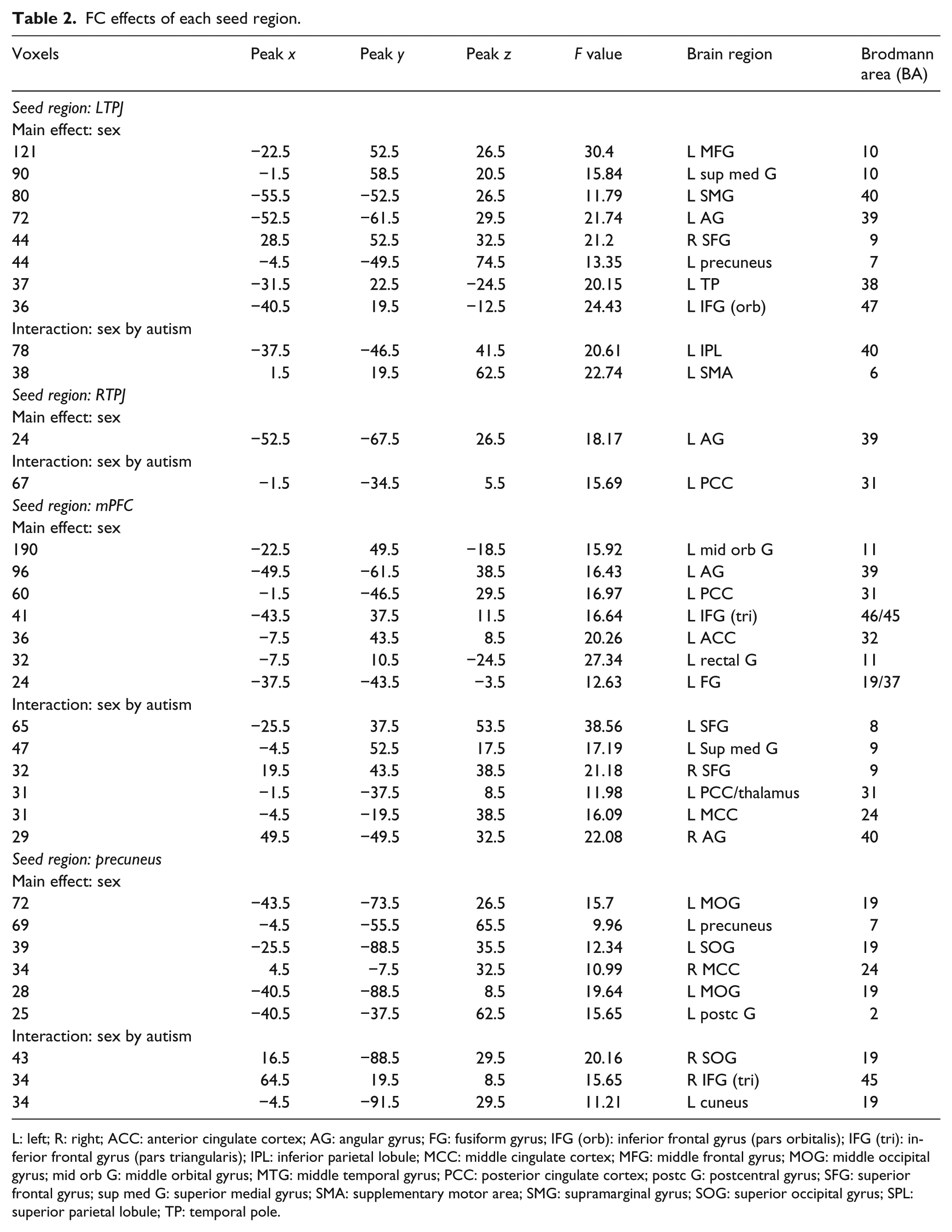
L: left; R: right; ACC: anterior cingulate cortex; AG: angular gyrus; FG: fusiform gyrus; IFG (orb): inferior frontal gyrus (pars orbitalis); IFG (tri): inferior frontal gyrus (pars triangularis); IPL: inferior parietal lobule; MCC: middle cingulate cortex; MFG: middle frontal gyrus; MOG: middle occipital gyrus; mid orb G: middle orbital gyrus; MTG: middle temporal gyrus; PCC: posterior cingulate cortex; postc G: postcentral gyrus; SFG: superior frontal gyrus; sup med G: superior medial gyrus; SMA: supplementary motor area; SMG: supramarginal gyrus; SOG: superior occipital gyrus; SPL: superior parietal lobule; TP: temporal pole.
ROI analysis of the FC interaction effects
The coordinates of the ROIs were in Table 2 (i.e. the interaction effects). For FC of the seed region left TPJ, significant interaction effects were found in the left IPL and left SMA. In the left IPL, the ROI analysis showed a significant interaction between sex and autism (F(1, 92) = 11.27, p = 0.001). Paired t-tests showed that within the females, the ASD group showed increased FC (t(23) = 2.67, p = 0.014), whereas in males, the ASD group showed reduced FC compared with the TD group (t(23) = 2.60, p = 0.016). In the left SMA, the ROI analysis indicated a significant interaction effect (F(1, 92) = 12.79, p = 0.001). Within the females, the ASD group showed reduced FC (t(23) = 2.85, p = 0.009), whereas in males, the ASD group showed increased FC (t(23) = 2.58, p = 0.017).
For FC in the seed region mPFC, several regions showed significant interaction effects. In the left MCC, the ROI analysis showed a significant interaction between sex and autism (F(1, 92) = 13.67, p < 0.001). The paired t-tests indicated that ASD females showed lower FC than TD females (t(23) = 2.94, p = 0.007), but ASD males showed higher FC than TD males (t(23) = 2.59, p = 0.016).
In the left MFG, the ROI analysis showed an interaction effect (F(1, 92) = 11.56, p = 0.001). The paired t-tests indicated no differences between ASD females and TD females (t(23) = 1.34, p = 0.19), but ASD males showed higher FC than TD males (t(23) = 4.14, p < 0.001).
In the left superior medial gyrus, the ROI analysis showed an interaction effect (F(1, 92) = 13.61, p < 0.001). The paired t-tests revealed that ASD females showed lower FC than TD females (t(23) = 3.81, p < 0.001), but no differences between ASD males and TD males (t(23) = 1.46, p = 0.16).
In the right AG, the ROI analysis showed an interaction effect (F(1, 92) = 15.02, p < 0.001). ASD females demonstrated reduced FC compared with TD females (t(23) = 4.32, p < 0.001). ASD males showed increased FC compared with TD males, but the difference was only marginally significant (t(23) = 1.91, p = 0.069).
In the right SFG, the ROI analysis indicated a significant interaction effect (F(1, 92) = 14.06, p < 0.001). The paired t-tests showed that ASD females had reduced FC compared with TD females (t(23) = 3.55, p = 0.002), but ASD males showed increased FC compared with TD males (t(23) = 2.64, p = 0.015).
For FC in the seed region precuneus, the left cuneus, right IFG (p. triangularis), and right SOG showed significant interactions in the whole-brain analysis. In the left cuneus, the ROI analysis showed an interaction effect between sex and autism (F(1, 92) = 11.49, p = 0.001). In the females, the ASD group showed lower FC than the TD group (t(23) = 2.77, p = 0.011). In the males, the ASD group showed increased FC compared with TD males, but the difference was only marginally significant (t(23) = 1.93, p = 0.066).
In the right IFG (p. triangularis), the ROI analysis showed an interaction effect between sex and autism (F(1, 92) = 15.12, p < 0.001). The paired t-tests showed that ASD females had reduced FC compared with TD females (t(23) = 3.54, p = 0.002), but ASD males showed increased FC compared with TD males (t(23) = 2.17, p = 0.04).
In the right SOG, the ROI analysis showed an interaction effect between sex and autism (F(1, 92) = 10.09, p = 0.002). ASD females demonstrated reduced FC compared with TD females (t(23) = 4.12, p < 0.001), but no difference was found between ASD males and TD males (t(23) = 1.24, p = 0.23).
For FC in the seed region right TPJ, the left PCC and left IFG (p. triangularis) showed significant interactions in the whole-brain analysis. In the left PCC, the ROI analysis showed an interaction effect between sex and autism (F(1, 92) = 5.69, p = 0.019). No difference was found between ASD and TD females (t(23) = 1.25, p = 0.22), but ASD males showed increased FC than TD males (t(23) = 2.69, p = 0.013).
In the left IFG (p. triangularis), the ROI analysis showed an interaction effect between sex and autism (F(1, 92) = 7.65, p = 0.007). In the females, the ASD group showed reduced FC compared with TD group, but the difference was only marginally significant (t(23) = 1.73, p = 0.093). In the males, the ASD group showed increased FC compared with the TD group, but the difference was marginally significant too (t(23) = 1.86, p = 0.076).
The ROI results were shown in Figure 2.
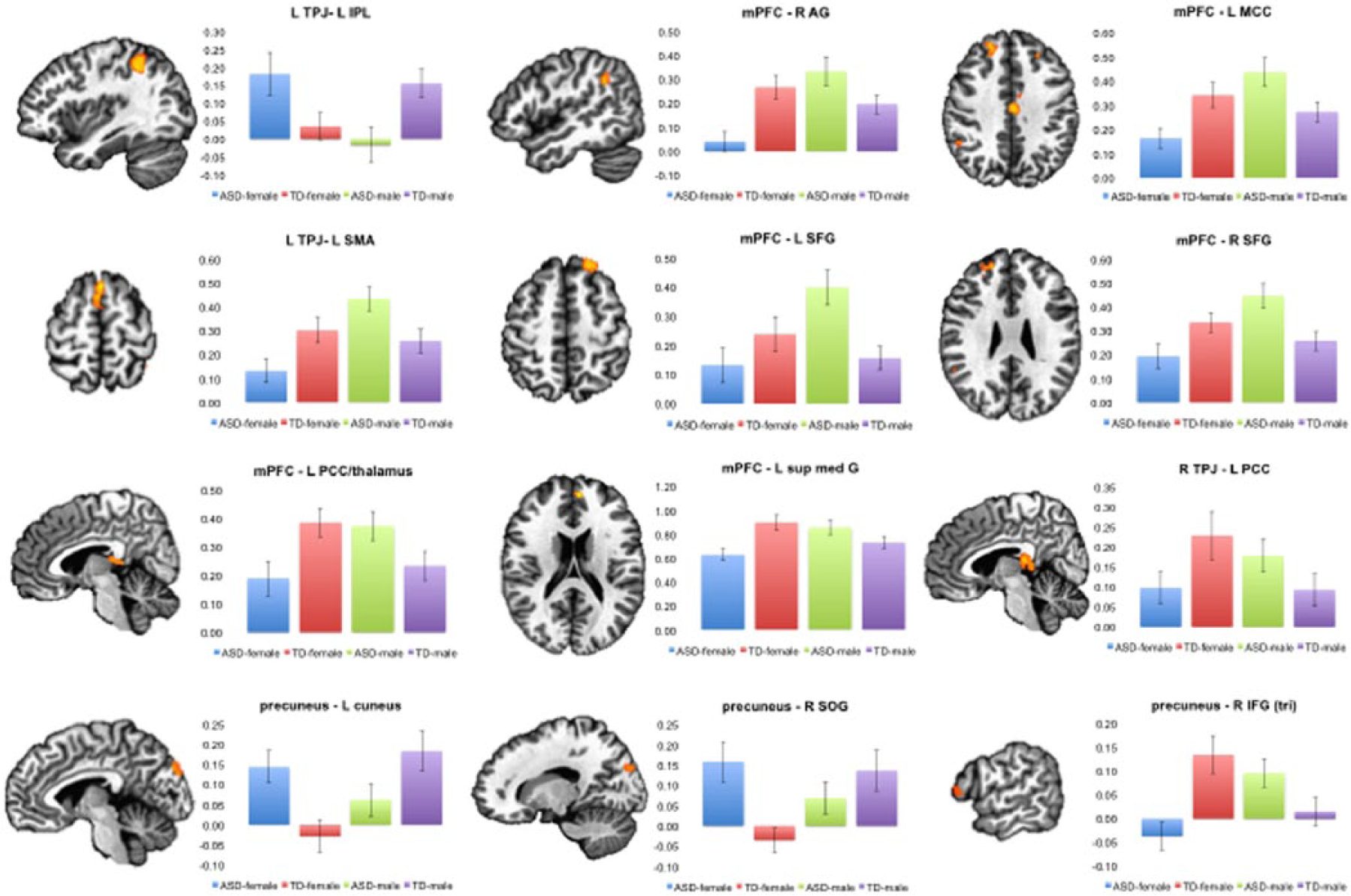
ROI analyses of the interaction effects from whole-brain ANOVA. In each histogram, the Y-axis indicates FC in the area. The error bars reflect standard errors.
Relation between ASD deficits and FC in ASD females and males
ASD females
For the precuneus, positive effects were found between ADOS_COMM and the FC in the right fusiform gyrus (FG) and right SFG (Figure 3(a)). No other effects were found.

Results of the regression analyses. (a)–(h): results in the ASD female group. (a) Seed region PCC: significant FC related to ADOS_COMM score. (b) Seed region mPFC: significant FC related to ADOS_COMM score. (c) Seed region mPFC: significant FC related to ADOS_SOCIAL score. (d) Seed region mPFC: significant FC related to ADOS_STEREO_BEHAV score. (e) Seed region left TPJ: significant FC related to ADOS_COMM score. (f) Seed region left TPJ: significant FC related to ADOS_SOCIAL score. (g) Seed region right TPJ: significant FC related to ADOS_COMM score. (h) Seed region PCC: significant FC related to ADOS_SOCIAL score. (i)–(j): results in the ASD male group. (i) Seed region left TPJ: significant FC related to ADOS_STEREO_BEHAV score. (j) Seed region right TPJ: significant FC related to ADOS_STEREO_BEHAV score.
For the mPFC, positive effects were found between ADOS_COMM and the FC in the right PCC and cerebellum (Figure 3(b)), and between ADOS_SOCIAL and the FC in the left superior parietal lobule (SPL; Figure 3(c)). In addition, a negative effect was found between ADOS_ STEREO_BEHAV and the FC in the left MOG (Figure 3(d)).
For the left TPJ, positive effects were found between ADOS_COMM and the FC in the left ACC/mid orbital gyrus, left IPL, bilateral thalamus, and left MFG (Figure 3(e)). Additionally, negative effects between ADOS_SOCIAL and the FC in the cerebellum, left IPL, and left MFG/SFG were found (Figure 3(f)).
For the right TPJ, positive effects were found between ADOS_COMM and the FC in the left SFG and left ACC (Figure 3(g)), and negative effects were found between ADOS_SOCIAL and the FC in the left MFG, bilateral superior medial gyrus, and right inferior temporal gyrus (ITG) (Figure 3(h)).
ASD males
For the left TPJ, positive effects were found between ADOS_ STEREO_BEHAV and the FC in the left precuneus, right mid orbital gyrus, right MOG, left SPL, and right IFG (p. opercularis) (Figure 3(i)). For the right TPJ, a positive effect was found between ADOS_ STEREO_BEHAV and the FC in the right mid orbital gyrus (Figure 3(j)). No significant effects were found between ADOS scores and FC for the mPFC and precuneus.
Table 3 and Figure 3 show the regression results.
Results of the regression analysis.
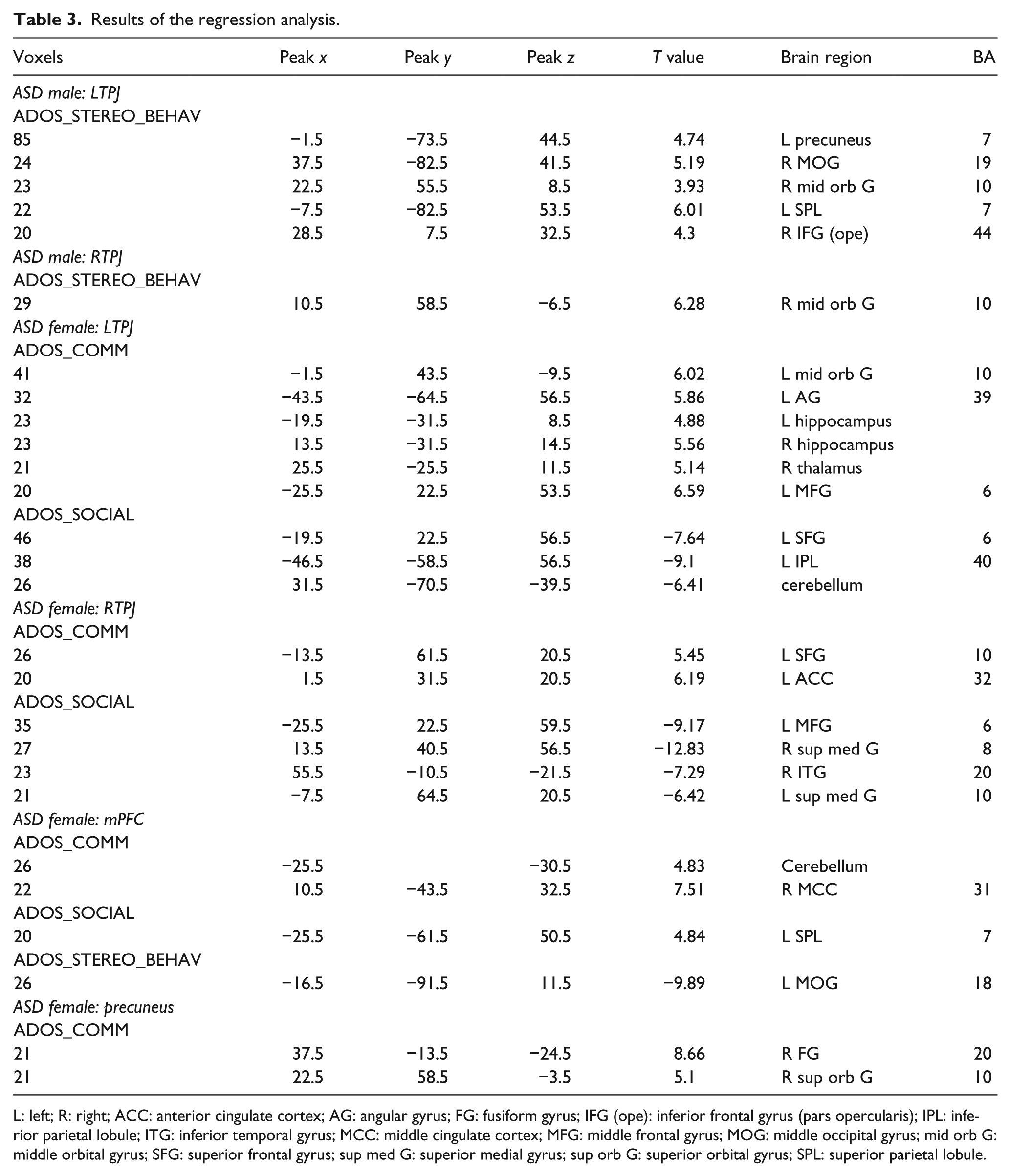
L: left; R: right; ACC: anterior cingulate cortex; AG: angular gyrus; FG: fusiform gyrus; IFG (ope): inferior frontal gyrus (pars opercularis); IPL: inferior parietal lobule; ITG: inferior temporal gyrus; MCC: middle cingulate cortex; MFG: middle frontal gyrus; MOG: middle occipital gyrus; mid orb G: middle orbital gyrus; SFG: superior frontal gyrus; sup med G: superior medial gyrus; sup orb G: superior orbital gyrus; SPL: superior parietal lobule.
Discussion
This study aimed to investigate the sex differences in the brain network of the mentalizing system in ASD individuals. To achieve this goal, we compared the intrinsic FC of the mentalizing brain regions in ASD males, ASD females, TD males, and TD females. Additionally, we calculated the relation between the FC effects in ASD males and females and the severity of their deficits.
The whole-brain ANOVA showed that compared with the TD participants, ASD females mainly demonstrated hypoconnectivity in the networks of mentalizing regions, whereas ASD males mainly showed hyperconnectivity in the mentalizing networks. Some previous studies (mainly recruiting male participants) have indicated that individuals with ASD showed hypoconnectivity compared with TD adolescents (e.g. Assaf et al., 2010; Cherkassky et al., 2006; Kana et al., 2009; Weng et al., 2010). Since previous research has rarely examined FC in females with ASD, it is hard to conclude whether the current result in ASD females is consistent with previous findings. However, the current result in ASD males (hyperconnectivity effects) seems inconsistent with the previous findings. We suggest that there might be several methodological reasons for the inconsistency.
First, the selections of seed regions are different in previous studies and this study. For example, in Assaf et al. (2010), ICA was used to analyze the DMN, and the seed regions were selected based on the activation in TD and ASD groups combined. In Weng et al. (2010), seed regions in the DMN were selected based on anatomical information. In this study, nevertheless, mentalizing seed regions were selected from a meta-analysis of fMRI studies about how mirror and mentalizing systems are involved in action comprehension (Van Overwalle and Baetens, 2009). The coordinates of seed regions in this study and the previous studies are different, which could cause different aberrant FC effects.
Second, this study and the previous studies used different fields of view (FOV) to analyze the FC effects. For example, in Assaf et al. (2010) and Kana et al. (2006, 2007, 2009), the FOVs were limited to action-derived ROIs, and in Weng et al. (2010), FOV was limited to anatomically determined ROIs (e.g. FC between a PCC seed and other areas of DMN was examined). In this study, however, the FC statistics were performed using a whole-brain FOV to maximize the search (i.e. FC between a mentalizing seed region and all other voxels in the brain was tested). The selection of FOV can significantly influence FC findings in ASD studies (Müller et al., 2011): studies reported global hypoconnectivity in ASD participants almost never reported whole-brain FC results and only focused on FC between ROIs; in contrast, many studies that do not support global hypoconnectivity in ASD used a whole-brain FOV. Thus, it is possible that the hypoconnectivity effects in ASD males reported in previous findings were because the FC effects were from a limited number of ROIs.
We also want to mention that within previous research that mainly recruited ASD males, the FC effects are not consistent. Although some findings supported general hypoconnectivity in ASD (e.g. Assaf et al., 2010; Just et al., 2004, 2007; Weng et al., 2010), other findings did not show general hypoconnectivity in ASD (Agam et al., 2010; Mizuno et al., 2006; Monk et al., 2009). Müller et al. (2011) suggested that some factors, such as low-pass filter, task-driven regression, and whole-brain FOV might influence the FC effect pattern.
The whole-brain contrast analyses within female and male participants showed that ASD males and females demonstrated different aberrant FC effects. First, the contrast analyses indicated that the two groups showed aberrant effects of different seed regions: in ASD males, the aberrant effects were in the FC of the bilateral TPJ, whereas in ASD females, the aberrant effects were in the FC of the mPFC, precuneus, and right TPJ. Previous research has shown that the TPJ is crucial for representing goals and intentions (Saxe and Kanwisher, 2003) and inferring others’ mental states (Van Overwalle, 2009), whereas the mPFC is involved in reasoning about actions and judgments (Keysers and Gazzola, 2007) and attribution (Van Overwalle, 2009). The precuneus and adjacent PCC may work as an interface between semantic network and the hippocampal memory system and help encode events into episodic memory (Binder and Desai, 2011), and researchers have found that the precuneus plays a key role in self-referential processing (Qin and Northoff, 2011; Van der Meer et al., 2010). The different aberrant FC effects in ASD males and females in this study may suggest impairments or dysfunctions in different social functions in ASD males and females (see section “Discussion” about the regression analysis).
Second, the contrast analyses revealed that the aberrant effects in ASD males and females had different directions. The ASD females mainly showed reduced FC in the networks of the mentalizing regions, whereas the ASD males showed increased FC effects. The ROI analyses demonstrated consistent results: in most regions showing significant sex × autism interaction effects, the ASD females had weaker FC than the TD females, whereas the ASD males had stronger FC than the TD males. It is suggested that hyperconnectivity may reflect a disorganized brain network that is inadequately selective, and that hypoconnectivity may show disrupted communication between brain areas (Courchesne and Pierce, 2005; Just et al., 2004, 2007, 2012; Minshew and Keller, 2010). In this study, ASD males showed long-distance hyperconnectivity between the TPJ and SFG/MFG, suggesting unselective information flow between remote regions in the mentalizing network. ASD females, on the contrary, not only had long-distance hypoconnectivity between the TPJ and mPFC, mPFC and AG, and precuneus and IFG, but also had short-distance hypoconnectivity between the TPJ and PCC, mPFC and ACC, and precuneus and PCC. This indicates that in ASD females, the communication is restricted in both local and global networks, and thus the information integration among widespread systems may not be effective (Courchesne and Pierce, 2005).
The regression analysis further confirms the sex differences in the aberrant FC effects in the ASD participants. For ASD males, only the FC effects of the bilateral TPJ were related to ASD deficits. Moreover, the relation was positive, that is, stronger FC corresponds to more severe deficits. This is consistent with the whole-brain ANOVA results that ASD males had hyperconnectivity in the networks of the bilateral TPJ. The interesting finding is that the FC between TPJ and mid orbital gyrus was related to ADOS_ STEREO_BEHAV that focuses on stereotyped behaviors and restricted interests in ASD males. The mid orbital gyrus is part of the orbitofrontal cortex (OFC), and the effect was adjacent to ACC, SFG, and medial frontal gyrus. OFC is anatomically synonymous with the ventromedial prefrontal cortex. In the human brain, its function is still not well understood. So far, researchers have proposed that the OFC is involved during sensory integration, the modulation of automatic reactions, and prediction as well as decision-making based on rewards (e.g. Kringelbach, 2005).
Several researchers have discussed the role of OFC in ASD deficits (Delmonte et al., 2013; Langen et al., 2011; Lewis and Kim, 2009; Uddin et al., 2015). For instance, Lewis and Kim (2009) discussed the neural basis of repetitive behavior in ASD, and suggested that cortical-basal ganglia circuitry plays an important role. In the circuitry, striosomal projection neurons receive input from limbic cortical areas, including OFC, ACC, and medial PFC, and directly mediate nigrostriatal dopamine pathway activity and in turn influence reinforced behavior. Langen et al. (2012) found that adults with ASD had aberrant fractional anisotropy of white matter tracts in the frontostriatal circuitry and worse go/no-go task performance (i.e. weaker inhibitory control) compared with healthy controls. Using resting-state fMRI, Delmonte et al. (2013) indicated that increased FC in frontostriatal circuitry was associated with restricted interests and repetitive behaviors in ASD. Taken together, these findings suggest that the OFC is involved in a network related to executive motor control and that the abnormality in the function or anatomy of the network is associated with the restricted repetitive behaviors in ASD.
The current result is in line with the above findings. Moreover, the result showed that the relation between OFC and restricted repetitive behaviors might mainly exist in ASD males. The positive relation between the TPJ-OFC FC and the score of restricted repetitive behaviors implies that higher-order cognitive functions, such as mentalizing, might play a role in this deficit. Turner (1999) divided restricted repetitive behaviors in ASD into two types: lower-order motor functions (e.g. stereotyped movements) and higher-order behaviors (e.g. compulsions and rituals). The higher-order behaviors have a distinct cognitive component by an adherence to a certain rule or mental set (e.g. having to do things in a certain way). The bilateral TPJ might be related with the higher-order behaviors because the regions are involved in thinking about self or others’ intentions, beliefs, and desires (Saxe and Kanwisher, 2003). In particular, the right TPJ is important in analyzing self-produced actions and signals from the external environment (Blakemore et al., 2002). The aberrant TPJ-OFC network in ASD males suggests that participants in this group might have dysfunction in mentalizing or executive control or both functions. It is also possible that mentalizing and executive control functions interact with each other and influence the behavior in ASD males. These possibilities require future work to test.
In ASD females, the regression results were quite different. The FC of all mentalizing regions was related to the ASD deficits, and both positive and negative effects were found. This might look strange since the main results of this study indicated that ASD females demonstrated reduced FC in the networks of mentalizing regions. We think there might be two reasons. First, although ASD females demonstrated decreased FC in many brain areas compared with TD females, they did show increased FC in some regions. For example, the whole-brain ANOVA revealed that for the seed region precuneus, ASD females showed increased FC in the left cuneus and right SOG. In addition, the ROI analysis showed that for the seed region left TPJ, ASD females showed increased FC in the left IPL. Thus, ASD females actually had a mixture of hyperconnectivity and hypoconnectivity in the networks for mentalizing regions. Second, in the regression analysis, we directly examined the relation between FC in the ASD participants and ADOS scores, rather than the relation between FC differences (ASD vs TD) and ADOS scores. We did try the latter but the effect could not survive the threshold. For the former analysis, we suggest that it reflects the relation between FC and ASD symptoms, regardless the aberrance of the FC effects. This means that even the FC and ADOS had a significant correlation, the FC might be still in the normal range and not significantly different from TD participants. In other words, the regression analysis indicated that some brain networks might have the potential to become aberrant. It is important to see that the related deficits focused on communicative and social deficits. For the communicative deficit, long-distance FC effects were related, including the FC between precuneus and SFG, mPFC and PCC, TPJ and ACC, and TPJ and MFG/SFG. These FC effects showed a positive relation with ADOS_COMM, suggesting that the global networks between the anterior and posterior regions play an important role in the communicative processes in ASD females. For the social deficits, the FC of the bilateral TPJ showed a negative relation, whereas the FC of the mPFC showed a positive relation. Both short- and long-distance FC effects were identified, including the FC between mPFC and SPL, TPJ and IPL, TPJ and MFG, TPJ and mPFC, and TPJ and ITG. These results suggest that both local and global networks of the mPFC and bilateral TPJ are involved in the social processes in ASD females. However, the relations between the whole-brain FC analysis and the regression analysis are not as clear as that in ASD males: in the whole-brain ANOVA, ASD females mainly demonstrated reduced FC of the mentalizing regions, but the regression analysis suggests that stronger FC could be associated with more severe deficits. To summarize, the regression analyses suggest that the dysfunction of the mentalizing system (i.e. aberrant FC effects) is associated with different aspects of ASD deficits in males (i.e. stereotyped behaviors) and females (i.e. communicative and social processes), and that the relation patterns are different.
Several issues need to be addressed in the future work. First, the relation between the hypoconnectivity and hyperconnectivity in ASD males and fameless and participants’ age is still unknown. Previous work has shown that age can influence the hypoconnectivity and hyperconnectivity effects in ASD (for a review, see Uddin et al., 2013). More specifically, ASD individuals who are about 9–12 years old tend to show hyperconnectivity (Di Martino et al., 2011; Lynch et al., 2013; Washington et al., 2014), whereas ASD individuals older than that tend to show hypoconnectivity (e.g. Anderson et al., 2011; Cherkassky et al., 2006; Ebisch et al., 2010). The real factor causing this pattern might be the onset of puberty, which can dramatically change individuals’ brain structurally and functionally (Uddin et al., 2013). This study revealed that sex could be another important factor that drive hyperconnectivity and hypoconnectivity in ASD individuals. How age and sex interact with each other during the development of the mentalizing system in ASD individuals requires future investigation.
Second, this study showed that ASD males and females had aberrant effects in the networks of different mentalizing regions, and that the aberrant effects were related to different ASD deficits. These results showed strong sex differences in the mentalizing network in ASD, but whether the same effects exist in mentalizing processes (e.g. ToM tasks, inference of intentions and goals, reasoning and judgment, and self-referential processing) still need to be tested in future studies.
Taken together, this study investigated the sex differences in the mentalizing network in ASD individuals. Using resting-state fMRI approach and FC analysis, we found that ASD males and females demonstrated different aberrant FC effects compared with TD participants. The sex differences were reflected in the networks of specific mentalizing regions, the direction of the effects, and the relations between the aberrant FC effects and the ASD deficits. Future work is needed to examine how sex factor interacts with other factors such as age and the sex differences during mentalizing task performance.
Footnotes
Funding
This work was supported by Macquarie University (grant no. 9201401500) and ARC Centre of Cognition and its Disorders Neural Markers Training Scheme Grant (funded out of CE110001021).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.