
Abstract
Background
Alzheimer's disease (AD) is recognized as a multifactorial neurodegenerative disorder involving numerous cellular and molecular processes, such as sleep disturbance, imbalance in brain glucose metabolism and neuroinflammation; these dysregulations typically precede the onset of symptoms. Hence, the results of mono-target therapy after AD diagnosis are in many cases unsatisfactory.
Objective
Traditional Chinese medicine (TCM) presents significant potential for treating AD. Sleep disorders are one of the early symptoms of AD; however, there is no effective solution to sleep disorders caused by AD. Some TCMs have been shown to treat sleep disorders by regulating energy metabolism or improving neuroinflammation. This study aims to investigate if XX-F administrated in advance could alleviate AD by improving sleep quality and neuroinflammation.
Methods
Mice were given Xiexintongfu formula (XX-F) intragastrically for three months. Morris water maze and pentobarbital-induced sleep test were performed to evaluate cognition and sleep. Determine changes in energy metabolism related to glycolysis through western blot and specific assay kits. Using immunofluorescence and western blot to detect neuroinflammation.
Results
Shortened sleep duration and cognitive impairment were observed in 6-month-old APP/PS1 mice. XX-F significantly prolonged sleep duration and rescued cognition. In addition, XX-F reduced the number of amyloid-β (Aβ) plaques and ameliorated neuroinflammation, and inhibited glycolysis by reducing pyruvate kinase M2 (PKM2) and lactate levels while rescuing adenosine triphosphate (ATP) deficiency.
Conclusions
We demonstrate that XX-F can improve sleep and cognition of AD mice by regulating energy metabolism and reducing neuroinflammation. This is a potential treatment method for AD and requires further in-depth research.
Keywords
Introduction
Alzheimer's disease (AD), the most prevalent form of dementia, is characterized by a complex interplay of pathophysiological alterations including memory deficits, 1 and sleep disturbances. 2 A hallmark feature of AD-related sleep dysfunction involves the disruption of normal sleep architecture, manifesting as nocturnal sleep fragmentation, excessive daytime napping, and significant reduction in both slow-wave sleep (SWS, N3) and rapid eye movement (REM) sleep.3,4 These sleep impairments are not merely symptoms but actively contribute to cognitive decline through multiple pathways, including memory consolidation deficits, accelerated biological aging. 5 Furthermore, emerging evidence underscores sleep deprivation as a significant modifiable risk factor for AD development. 6 Notably, sleep deterioration has been observed even during the preclinical stages of AD, correlating with early AD pathology. 7 This relationship appears bidirectional: sleep disturbances impair clearance of amyloid-β (Aβ), leading to its abnormal accumulation, while Aβ deposition itself further exacerbates sleep dysfunction. 8 The resulting neuroinflammatory cascade, characterized by microglial activation and pro-inflammatory cytokine release, creates a vicious cycle that accelerates disease progression. 9
Traditional Chinese medicine (TCM) has accumulated substantial empirical evidence in treating sleep disorders over its 2000-year clinical history, offering distinct advantages including sustained therapeutic effects and favorable safety profiles.10,11 Current research demonstrates that TCM interventions exert significant effects on sleep regulation through diverse pathways: soy isoflavones ameliorate sleep deprivation-induced cognitive deficits via antioxidant and anti-inflammatory mechanisms. 12
The present study investigates a novel TCM formulation developed by integrating two classical prescriptions: one with well-established sleep-promoting properties and another demonstrating efficacy in improving cerebral energy metabolism in APP/PS1 mice. 13 This innovative combination embodies TCM's holistic treatment strategy, 14 potentially addressing both the symptomatic and pathological aspects of AD-related sleep disturbances. We hypothesize that early intervention with this formulation may prevent AD progression, offering a promising complementary approach to conventional AD therapeutics. Our research aims to bridge traditional medicine with modern neurobiological understanding, potentially yielding novel insights into integrative AD prevention strategies.
Methods
Animals
Three-month-old male APP/PS1 mice and C57BL/6 wild-type mice were purchased from SPF (Beijing) Biotechnology Co., Ltd Experimental procedure was approved by Beijing University of Chinese medicine (BUCM) Medical and laboratory animal ethics committee (BUCM-4-2021072501-3175). Mice were randomly grouped after 3 days of acclimatization in the SPF-level laboratory at Liangxiang Campus and weighed once a month for a total of four times. They were allowed to freely eat and drink throughout the experiment. The room lights were strictly monitored to maintain a regular circadian rhythm.
Formula preparation, quality control, and administration
Xiexintongfu formula (XX-F) was purchased from Shangren yijia Chinese Medicine Clinic Co and contains a total of 9 herbs. Based on previous studies on the biological activity range of the XX-F component and the pharmacokinetic characteristics of oral administration,15–19 XX-F was processed into three doses: 0.625 g/ml as the low dose, 1.25 g/ml as the medium dose (clinical equivalent dose) and 2.5 g/ml as high dose.20,21 Mice were orally administered different doses of XX-F (high dose: 50 g/kg; Medium dose: 25 g/kg Low dose: 12.5 g/kg. The clinical dose of XX-F in the human body is 1.46 g/kg/d (the total weight of all herbs in a single XX-F prescription is 102 g, and the average human body weight is 70 kg), and the moderate dose (25 g/kg) in mice is equivalent to the clinical dose converted based on body surface area. The administered doses were calculated by converting clinically effective human doses to mouse-equivalent doses using the body surface area normalization method.22,23 This approach is widely adopted in TCM pharmacological research. 24 The intermediate dose group represents the clinical equivalent dose, while the high and low doses were set at 2-fold higher and lower respectively to evaluate dose-dependent effects. Donepezil (a reversible AChE inhibitor) was selected as the positive control drug, an FDA-approved oral drug administered at a clinically equivalent dose. Donepezil was diluted with purified water to a concentration of 2 mg/kg and kept at room temperature. 25 Mice in the donepezil group were orally administered donepezil. According to the established plan, 26 we inserted the gavage needle into the mouth of the mouse, gently pushed the gavage needle towards the stomach, and injected the decoction formula into the stomach of the mouse. The time course of intervention was 3 months, once a day. Same volume of purified water was given to the mice in the WT and model groups. Administration was through intragastric route.26,27 The raw material of formula was added to the decoction pot and adequate water was added. After 40 min heating, the solution was collected, filtered, and the residue was removed by a filter. The collected solution was centrifuged for 10 min, and the supernatant was subjected into ultra-performance liquid chromatography for parameters optimization. Subsequently, the samples were analyzed by liquid chromatograph. A Waters ACQUITY UPLC-AB SCIEX Triple TOF 6600 mass spectrometer was used; ACQUITY CSH C18 column (2.1 × 150 mm, 1.7 μm); water containing 0.1% formic acid as mobile phase A and acetonitrile containing 0.1% formic acid as mobile phase B; gradient elution: 0–8 min: 2%–20% B; 8–20 min: 20%–35% B; 20–32 min: 35%–95% B; 32–33 min: 95%–2% B; flow rate: 0.3 mL/min; column temperature: 50 °C; injection volume: 3 μL. 20–32 min: 35%–95% B; 32–33 min: 95%–2% B; flow rate: 0.3 mL/min; column temperature: 50 °C; injection volume: 3 μL. Mass spectrometry was performed using an ESI ion source with positive and negative ion mode scanning; nitrogen as the nebulizer gas, Curtain gas at 35 psi; Gas 1 at 55 psi; Gas 2 at 55 psi; ISVF at 5.5 kV (positive ion mode), −4.5 kV (negative ion mode); CE at 40 V; DP at 80 V; TEM at 550 °C (positive ion mode), 550 °C (negative ion mode). mode), 550 °C (negative ion mode); ion scan range of m/z 100–2000.
Pentobarbital-induced sleep test
Sodium pentobarbital (55 mg/kg) was given intraperitoneally 1 h after last administration of XX-F, water or donepezil. The onset of sleep is the time that the mice stayed immobile and lost their righting reflex. The time interval between administration of pentobarbital and onset of sleep was considered. sleep latency the return is defined by the two times of righting reflex within 1 min. The time from the loss of righting reflex to its return were considered sleep duration.
Morris water maze
Morris water maze consists of a large circular pool of water with a fixed platform hidden just below the surface of the water. Four equally spaced locations around the edge of the pool divided the pool into four equal quadrants. The platform is rendered invisible by adding powdered milk. Geometric images are hung on the pool wall as visual cues for navigation. Before the experiment started, mice were housed in the water maze room to adapt to the surrounding environment. Learning trials: a method in which each mouse was given two trials per day for six consecutive days. The trial was initiated by placing mice in the water facing the pool wall in one of the four quadrants. The daily order of entry into individual quadrants was randomized so that all four quadrants were used once every two days. Mice were allowed to swim a maximum of 60 s to find the hidden platform. If unsuccessful within the allotted time period, the mice were given a score of 60 s and guided to the platform and stayed for 15 s. Probe trial: on day 7, a probe trial was conducted in which the platform is removed from the pool. Mice were placed in the pool to swim freely for 60 s. An annulus ring can be circumscribed around the previous target location (on the computer screen). The number of crossings through platform was recorded. After behavioral test, mice were anesthetized, sacrificed and grouped for different brain extraction process. Fresh brain tissue was collected from one group of mice and frozen, while the other group underwent paraformaldehyde fixation and sectioning for staining.
Tissue collection
Following the final behavioral test, mice were anesthetized by intraperitoneal injection of avertin. Cardiac perfusion was initiated through the left ventricle with saline until complete blood clearance was achieved. For a subset of mice, hippocampal tissues were immediately dissected and snap-frozen in cryotubes at −80 °C for subsequent western blot analysis. The remaining animals underwent whole-body perfusion with pre-chilled 4% paraformaldehyde (PFA) for tissue fixation. Following perfusion, brains were carefully extracted and post-fixed in 4% PFA solution for later Immunofluorescence staining.
Immunofluorescence
PBST solution (0.7% Triton-X-100 in PBS) and 10% blocking buffer (10% goat serum in PBST) was prepared. Brain sections were first incubated with blocking buffer overnight at 4 °C, followed by primary antibody incubation for 8 h at 4 °C. The sections were washed with PBST for three times and incubated with secondary antibody for 2 h at room temperature avoiding light. Next, sections were washed three times with PBST and processed through nucleus staining with DAPI for 10 min. The antibodies selected in this study are as follows: rabbit anti Iba-1, 1:1000 (Wako, 019-19741); mouse anti PKM2, 1:250 (Proteintech, 15822-1-AP); goat anti rabbit Alexa fluor 594, 1:500 (cell signaling technology); goat anti mouse Alexa fluor 647 1:500 (cell signaling technology). The images were acquired using a confocal microscope (Olympus).
For image acquisition, sections were mounted on Superfrost Plus slides and coverslipped with anti-fade mounting medium. Z-stack images (5–7 slices at 1-μm intervals) were captured using an Olympus FV3000 confocal microscope with consistent laser power and gain settings across all samples. Quantitative analysis was performed by researchers blinded to experimental groups using ImageJ software (NIH). Aβ plaques were manually counted in three non-overlapping hippocampal fields per section at 20× magnification. For Iba1 and PKM2 quantification, threshold-based area fraction analysis was performed after background subtraction using identical parameters across all images. Colocalization analysis of Iba1 and PKM2 was determined using the ImageJ JACoP plugin, with positive signals defined as pixels exceeding threshold values in both channels. Three sections per animal and 5–6 animals per group were analyzed to ensure statistical power.
Adenosine triphosphate (ATP) and lactate measurement
Frozen brain tissues were homogenized with lysis buffer and sonicated on ice, followed by centrifugation at 12,000 g for 10 min at 4 °C. The supernatants were collected, and the ATP as well as lactate levels were measured using a specific Kits (Lablead, CLC050 and ATP100) according to the manufacturer's instructions.
Western blot
Proteins were first extracted from fresh mouse hippocampal tissues using lysis buffer and then quantified using the BCA method. Proteins were run on SDS-PAGE and then transferred electroblotting onto PVDF membrane. The membranes were soaked and blocked with western blotting buffer and incubated with primary antibodies overnight at 4 °C. Afterwards, the membrane was washed and incubated with HRP-conjugated secondary antibody for 1.5 h at room temperature. The antibodies selected in this study are as follows: rabbit anti PKM2, 1:1000 (cell signaling technology, 4053S); mouse anti β-actin, 1:1000 (ZSGB-BIO, 16AF0207); rabbit anti TNF-α, 1:500 (ABclonal, A11534); goat anti-Rabbit IgG-HRP, 1:10000 (Lablead, S0101); goat anti-Mouse IgG (H + L)-HRP, 1:10000 (Lablead, S0100). The bound reactivity was visualized using ECL system.
Statistical analysis
All data are presented as mean ± standard error of mean (SEM). The significance of differences was assessed by one-way analysis of variance (ANOVA) followed by Tukey's multiple comparisons test. Significance was set as p < 0.05 and expressed as *p < 0.05, **p < 0.01, ***p < 0.001.
Results
XX-F consists of nine herbal drugs and quality control by high performance liquid chromatography-mass spectrometry
XX-F consists of 9 traditional Chinese medicines, including Huang-Lian (Coptis Chinensis), Huang-Qin (Skullcap), Fa-Ban-Xia (Rhizoma Pinellinae Praeparata), Dan-Nan-Xing (Arisaema Cum Bile), Zhi-Gan-Cao (Prepared Licorice), Ren-Shen (Ginseng), Gan-Jiang (rhizoma zingiberis), Fu-Ling (Poria Cocos), E-Jiao (donkey-hide gelatin) (Figure 1). XX-F decoction were analyzed by high performance liquid chromatography-mass spectrometry. We have discovered 26 known active ingredients, belonging to Coptis Chinensis, Skullcap, Ginseng, Prepared Licorice, and Poria Cocos, respectively. Among them, Coptis Chinensis, Skullcap, and Ginseng have the highest content of effective ingredients. We intragastrically administered the quality-controlled XX-F (Supplemental Table 1) to 3-month-old APP/PS1 mice for 3 months.
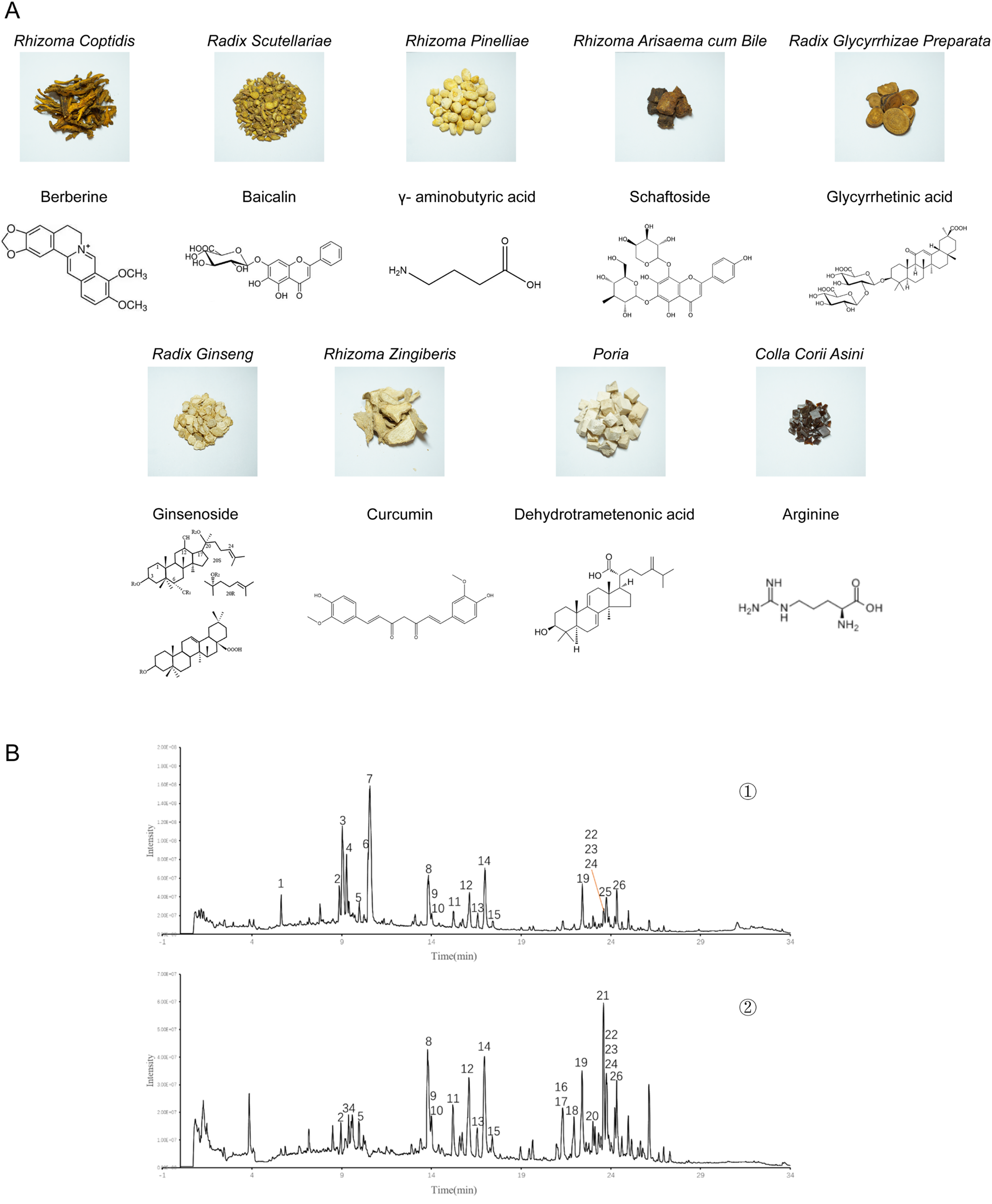
Composition and effective ingredients of XX-F. A. The traditional Chinese medicine and its main effective ingredients contained in the XX-F. B. UPLC-Q-TOF-MS for quality control. ①-②. Positive and negative mode, respectively.
XX-F regulated pentobarbital Sodium-induced sleep of APP/PS1 mice
The pentobarbital-induced sleep test showed that the sleep duration of mice in model group was significantly lower than that in wide-type (WT) group. The sleep duration in the low and medium-dose of the XX-F was significantly higher compared with the model group, and the mice in the medium-dose group had the longest sleep duration. Compared with the WT group, the sleep latency of mice in the model group had a tendency to increase, and the sleep latency in the medium-dose group was significantly shorter compared with the model group (Figure 2B, C). The above suggests that the XX-F is relatively safe at the given doses and can effectively prevent sleep disturbance that occurs in APP/PS1 mice at 6 months of age.
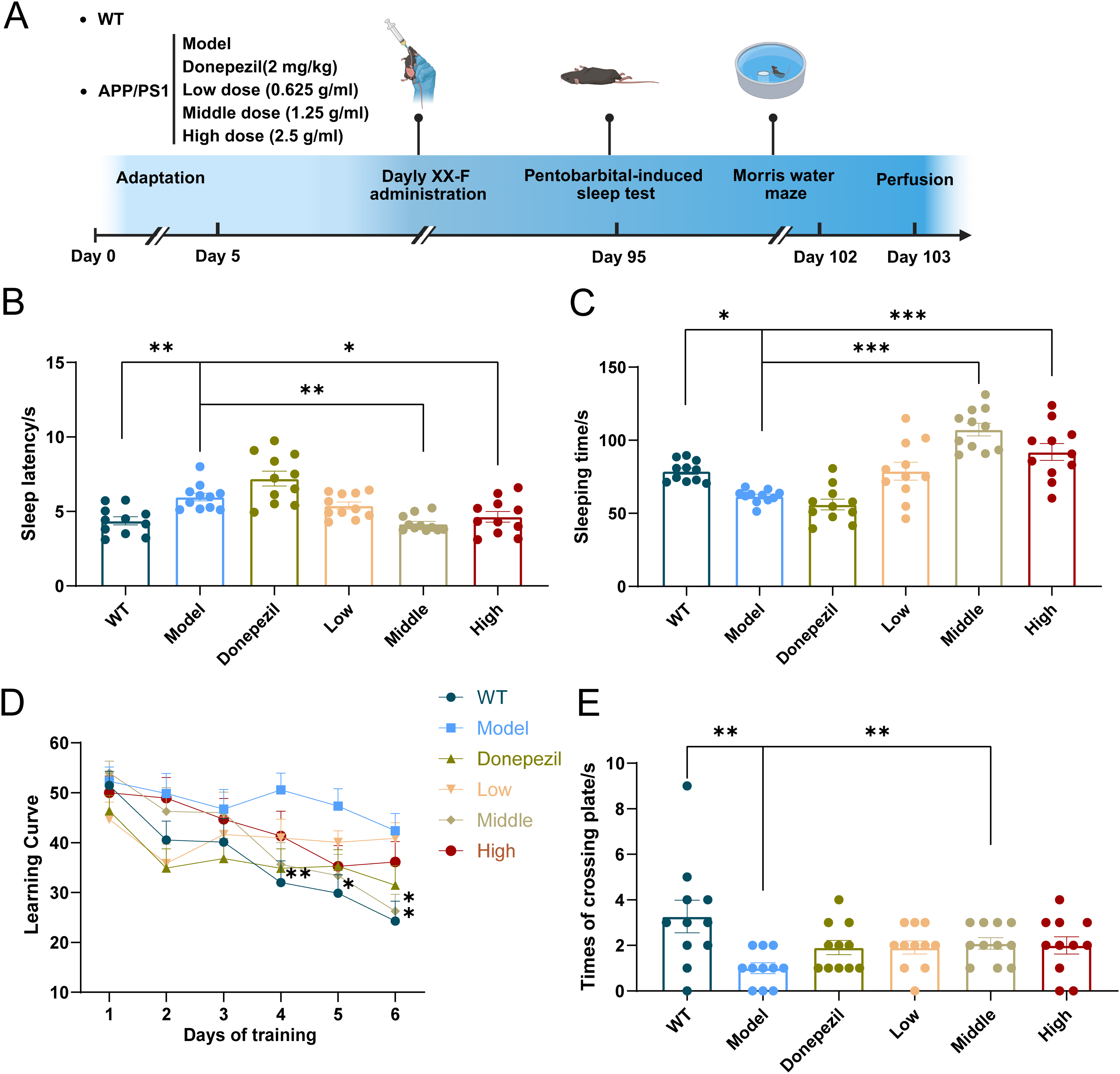
XX-F improves sleep and cognition of APP/PS1 mice. (A). Flowchart of the experimental intervention. (B) Sleep latency of the mice (s). (C) Sleep time of the mice (s). (D) Learning curve. (E) Numbers of mice crossing the platform. Data shown are mean ± SEM. n = 11 in each group. Compared with model group, ***p < 0.001; **p < 0.01; *p < 0.05.
XX-F improved learning and spatial memory of APP/PS1 mice
Cognitive function in the mice was evaluated using the Morris water maze test. In the learning session, from forth day the escape latency of mice in the WT group started to be significantly shorter compared with model group. Also from this time point, the escape latency of mice in the low-dose and donepezil group, rather than medium and high-dose group (data not shown), is significantly shorter compared with model group (Figure 2D). In the probe session, mice in the model group had significantly fewer platform crossings than those in the WT group. Mice in the medium-dose group crossed the platform significantly more times than the model group (Figure 2E). The above results demonstrate that APP/PS1 mice develop pronounced impairments in both learning and memory abilities at 6 months of age. A medium dose of the XX-F was effective in preventing cognitive decline and donepezil could only rescue dysfunction of learning instead of memory ability. Due to the poor effect of low and high doses, we focused on medium-dose XX-F for subsequent experiments.
XX-F alleviated Aβ pathology of APP/PS1 mice
We used antibody staining to detect Aβ pathology. There were robust amyloid plaque and Aβ1−42 in the brain of APP/PS1 mice and XX-F significantly reduced the number the plaque and Aβ. Besides, the GFAP positive area, which indicates astrocyte, was increased in the brain of APP/PS1mice and XX-F decreased the expression of GFAP. However, there was no significant difference of GFAP area as well as Aβ pathology between Donepezil and model group (Figure 3).
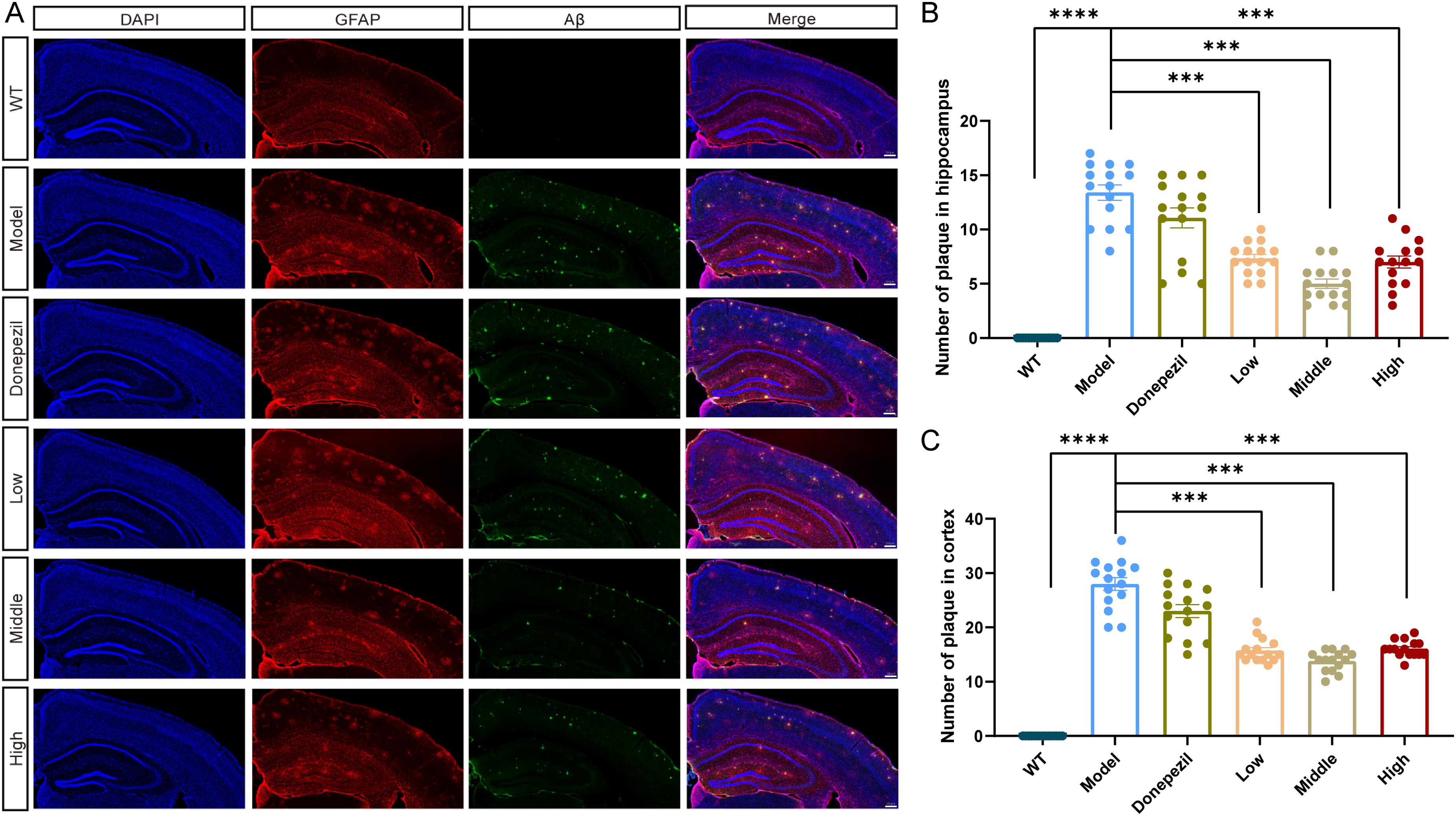
XX-F decreases Aβ pathology of APP/PS1 mice. (A) The representative immunofluorescence staining image of Aβ1-42 (Green) and GFAP (Red) in brain. B. Quantification of amyloid plaques in hippocampus. (C) Quantification of amyloid plaques in cortex. Data shown are mean ± SEM. n = 3 in each group. Compared with model group, ***p < 0.001; **p < 0.01; *p < 0.05.
XX-F attenuated neuroinflammation in APP/PS1 mice
We used ionized calcium-binding adapter molecule 1 (Iba-1) as a marker to visualize microglia in the mice brain. Immunofluorescent staining showed more Iba-1 positive area in the brain of model group, especially around the Aβ plaques. The shape of the microglia in the model group exhibit enlarged cell body and shortened processes. XX-F administration exerted region-dependent anti-inflammatory effects, with medium-dose administration significantly reducing Iba-1/Aβ colocalization in the hippocampus, while all doses (low, medium, high) attenuated cortical microglial activation (Figure 4C, D).
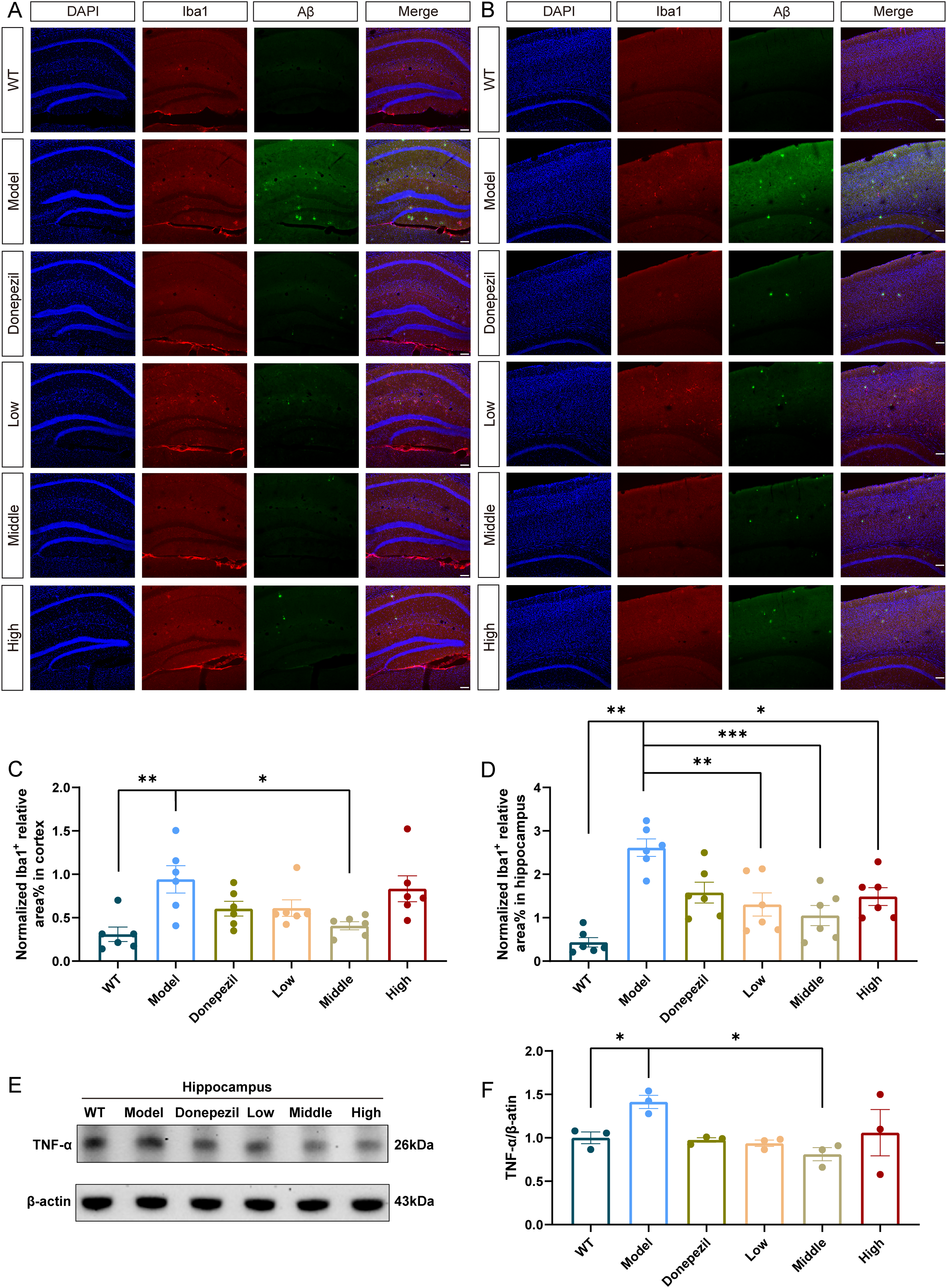
XX-F inhibits neuroinflammation of APP/PS1 mice. (A) The representative immunofluorescence staining image of Iba-1 (Red) and DAPI (Blue) in hippocampus. (B) The representative immunofluorescence staining image of Iba-1 (Red) and DAPI (Blue) in cortex. (C) Quantification of Iba-1 positive area in cortex. (D) Quantification of Iba-1 positive area in hippocampus. (E) The representative image of western blot. (F) Relative expression of TNF-α. Data shown are mean ± SEM. n = 3 in each group. Compared with model group, ***p < 0.001; **p < 0.01; *p < 0.05.
Parallel western blot analysis demonstrated that hippocampal tumor necrosis factor-α (TNF-α) expression was significantly elevated in vehicle-treated APP/PS1 mice, which was normalized by medium-dose XX-F administration (Figure 4E, F). These findings collectively demonstrate that XX-F effectively suppresses multiple markers of neuroinflammation in APP/PS1 mice, including microglial activation and pro-inflammatory cytokine production.
XX-F restored energy metabolism in APP/PS1 mice
Building upon our findings of XX-F's anti-neuroinflammatory effects, we investigated its impact on cerebral energy metabolism. Biochemical analyses revealed significant metabolic dysregulation in APP/PS1 mice, characterized by: (1) elevated protein expression of pyruvate kinase M2 (PKM2), the rate-limiting glycolysis enzyme; (2) increased lactate accumulation; and (3) reduced ATP levels (Figure 5). Medium-dose XX-F administration effectively normalized these metabolic parameters, reducing PKM2 expression and lactate levels, while restoring ATP production (Figure 5C-F).
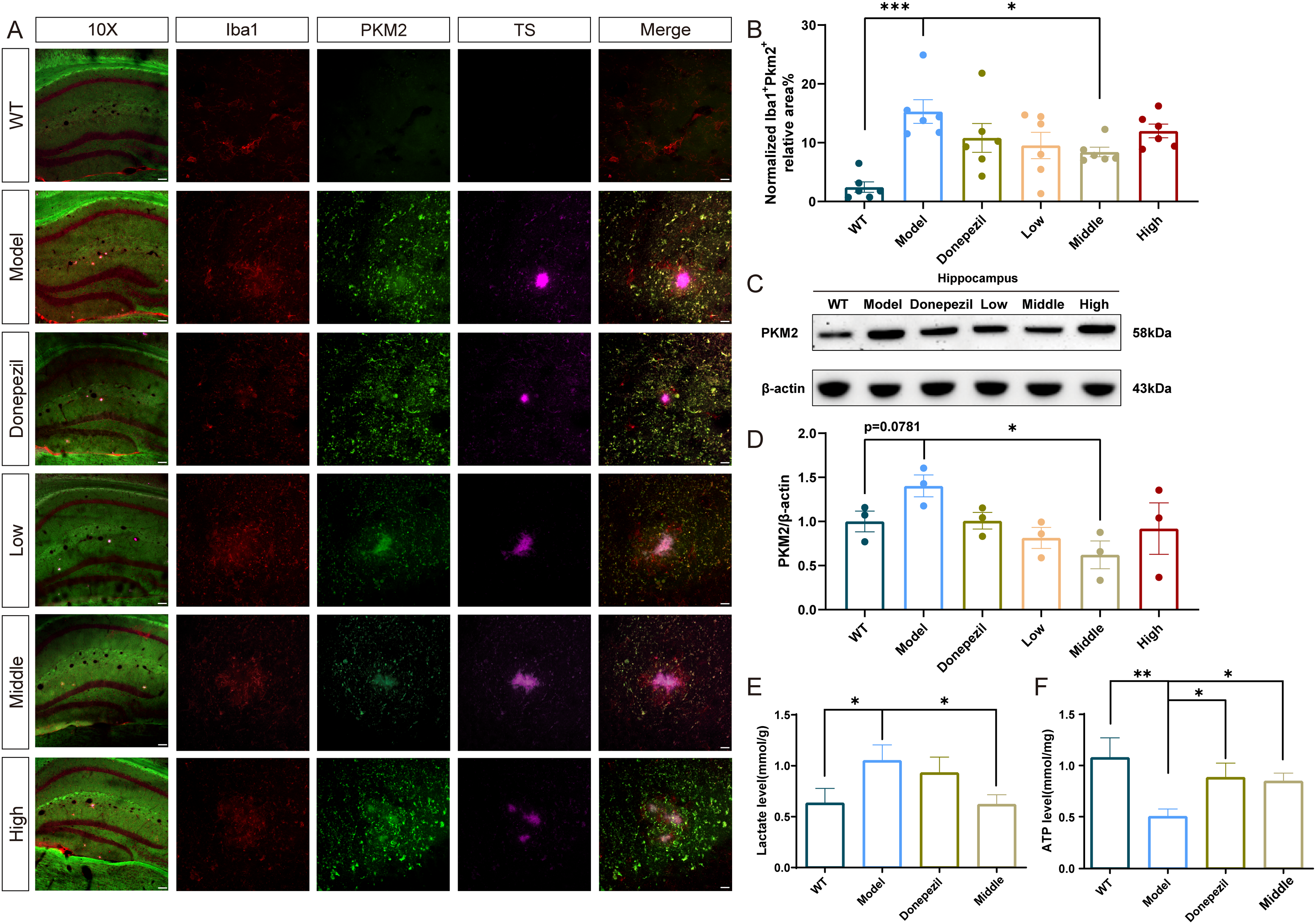
XX-F suppressed brain glycolysis of APP/PS1 mice. (A) The representative immunofluorescence co-staining image of Iba-1 (Red), PKM2 (Green) and amyloid plaque (Purple) in brain. (B) Quantification of Iba-1 and PKM2 co-localized area. (C) The representative image of western blot. (D) Relative expression of PKM2. (E) Quantification of lactate levels in brain. (F) Quantification of ATP levels in brain. Data shown are mean ± SEM. n = 3 in each group. Compared with model group, ***p < 0.001; **p < 0.01; *p < 0.05.
Immunofluorescence co-localization studies provided cellular-level insights, demonstrating intense PKM2 immunoreactivity (Figure 5A, green fluorescence) in Iba-1+ microglia surrounding Aβ plaques. Quantitative analysis showed a reduction in Iba-1/PKM2 co-localization in middle-dose XX-F treated mice (Figure 5B), suggesting microglia as a primary target for XX-F's metabolic regulation. These results demonstrate that XX-F ameliorates the pathological glycolytic shift in APP/PS1 mice, restoring energy homeostasis through PKM2 modulation in activated microglia.
Discussion
Here we identify the novel TCM formulation XX-F exerts dual therapeutic effects by simultaneously improving sleep quality and cognitive function in APP/PS1 mice. These findings support growing clinical evidence linking sleep disturbances to AD pathogenesis, where chronic sleep deprivation has been identified as a significant modifiable risk factor for AD development. 28 Notably, our results align with observations that sleep quality preservation may help maintain cognitive function during early AD stage, 29 and that sleep interventions can mitigate AD pathology in animal models.30,31 The current work extends these findings by revealing that XX-F's therapeutic benefits may stem from its unique ability to concurrently modulate two key pathological processes: cerebral energy metabolism dysregulation and neuroinflammation.
The metabolic alterations observed in APP/PS1 mice - characterized by enhanced glycolysis (evidenced by elevated PKM2 and lactate) coupled with impaired ATP production - mirror the brain's metabolic shift seen in sleep deprivation. 32 XX-F's capacity to normalize this metabolic imbalance suggests its potential to restore cerebral energy homeostasis, which may underlie its sleep-regulating effects. Furthermore, our findings support the proposed neuroinflammatory cascade wherein sleep disturbances promote oxidative stress and microglial activation,33,34 creating a vicious cycle that accelerates AD progression.
The neuroinflammatory cascade in AD involves extensive glial activation and the release of damaging cytokines, among which tumor necrosis factor (TNF) plays a central role. TNF, originally identified for its antitumor effects, is now recognized as a pleiotropic cytokine involved in immune regulation, sleep modulation, and metabolic control.35,36 Our findings demonstrate that XX-F effectively suppresses TNF-α overexpression in APP/PS1 mice, suggesting its potential to interrupt this inflammatory cascade.
A critical link exists between neuroinflammation and metabolic dysfunction in AD pathogenesis. The brain primarily relies on glucose metabolism, with oxidative phosphorylation generating ∼97.5% of ATP under normal conditions. However, in early AD, the brain undergoes a metabolic shift toward aerobic glycolysis (the “Warburg-like effect”). 37 This adaptation may initially protect neurons from oxidative stress but ultimately exacerbates disease progression. 38 Our results confirm this hypothesis, showing elevated PKM2 (a key glycolytic enzyme) and lactate levels in APP/PS1 mice, alongside reduced ATP production. Notably, XX-F treatment reversed these metabolic abnormalities, suggesting its ability to restore energy homeostasis.
Furthermore, emerging evidence highlights microglial metabolic reprogramming as a driver of neuroinflammation in AD. Aβ deposition triggers a glycolytic shift in microglia, promoting their pro-inflammatory activation while impairing phagocytic function. 39 Our immunofluorescence data reveal significant PKM2 expression in plaque-associated microglia, with XX-F treatment reducing this co-localization. This aligns with recent studies demonstrating that PKM2 mediates inflammatory activation in macrophages 40 and that glycolytic reprogramming contributes to AD pathogenesis through epigenetic mechanisms involving lactate-derived histone modifications.
XX-F demonstrated dose-dependent effects, with the medium dose (clinical equivalent) showing optimal efficacy in improving both sleep parameters and cognitive performance. Intriguingly, while donepezil improved learning in the acquisition phase, it failed to rescue probe trial performance or attenuate Aβ pathology and neuroinflammation. 41 There are several possible reasons: donepezil is associated with sleep disturbances (abnormal dreams, vivid dreams and nightmares) in clinical practice.42,43 In addition, for sleep deprivation model of cognitive impairment, donepezil was not able to reverse this performance decline.44,45 Moreover, the cholinergic mechanism of donepezil may not address the predominant pathology in Alzheimer's model mice, 46 consistent with its inefficacy in sleep deprivation-induced cognitive impairment and potential adverse effects in a subgroup of patients with frontotemporal dementia can experience worsening of symptoms with donepezil.47,48
The present study shows the complexity of AD and different animal models can only represent one aspect of AD pathology. This greatly limits the development of therapeutic drugs for AD disease. TCM has outstanding performance in treating complex diseases of multiple systems. As demonstrated in this study, by regulating energy metabolism and reducing neuroinflammation, the XX-F can improve sleep and cognitive impairment. And the exploration of deeper mechanisms of action may bring a promising new direction for the treatment of AD disease.
Conclusion
XX-F can improve sleep and prevent cognitive impairment in APP/PS1 mice. Regulating brain glycolysis and inhibiting neuroinflammation may be the main mechanisms of its action, and the glycolytic enzyme PKM2 may be a key target.
Supplemental Material
sj-docx-1-alz-10.1177_13872877251355596 - Supplemental material for A novel traditional Chinese medicine formula restores sleep and cognitive function in APP/PS1 mice by targeting glycolytic pathways and neuroinflammatory responses
Supplemental material, sj-docx-1-alz-10.1177_13872877251355596 for A novel traditional Chinese medicine formula restores sleep and cognitive function in APP/PS1 mice by Jiani Zhang, Chunxiang Wang, Kexin Chang, Tiantian Peng, Chengbang Liang, Junshi Cheng, Yu Shi, Xu Wang, Zhaoyang Wang, Yan Tan and Qian Hua in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
The authors have no acknowledgments to report.
ORCID iD
Ethical considerations
The Medical and Laboratory Animal Ethics Committee of Beijing University of Traditional Chinese Medicine approved all experimental procedures involving animals (BUCM-4-2021072501-3175).
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by the National Natural Science Foundation of China (No. U21A201401 to Qian Hua, No. 82374175 to Yan Tan, No. 82304915 to Xu Wang).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.