
Abstract
This review elucidates the complex interplay among oxidative stress (OS), macrophage polarization, and stem cell-driven osteogenesis, emphasizing the regulatory influence of reactive oxygen species (ROS) on bone repair and regeneration. It demonstrates that an imbalance in ROS can impede bone healing by disrupting the equilibrium between pro-inflammatory (M1) and pro-repair (M2) macrophage phenotypes. Furthermore, the review delineates the mechanisms through which ROS can influence mesenchymal stem cell differentiation and osteoclast activity, while also highlighting the body’s antioxidant defenses that counteract OS. Innovative strategies are explored, particularly the use of biomaterials and nanomedicine, which aim to modulate ROS levels and macrophage polarization, thereby fostering a conducive microenvironment for bone regeneration. The integration of nanotechnology, biomaterials, and cellular biology emerges as a promising frontier for advancing bone regeneration therapies, with the necessity for clinical validation underscored throughout.
Impact Statement
This review establishes redox modulation as a paradigm-shifting strategy for bone regeneration. We elucidate how engineered biocomposites precisely recalibrate reactive oxygen species (ROS) to resolve osteo-inflammation, directing macrophage polarization from pro-inflammatory (M1) to pro-regenerative (M2) phenotypes. This immune reprogramming synergistically enhances mesenchymal stem cell osteogenesis and suppresses osteoclastogenesis. By integrating cutting-edge biomaterial design—including enzyme-mimetic nanozymes and organelle-targeted antioxidants—we highlight clinically viable solutions for diabetic bone defects, osteoporosis, and rheumatoid arthritis. Our framework bridges immunology, nanotechnology, and tissue engineering, offering transformative therapeutic avenues for inflammatory osteopathies.
Introduction
Bone inflammatory diseases, such as osteoarthritis (OA) and osteoporosis (OP), pose significant global health challenges due to their complex pathogenesis, which involves oxidative stress (OS) and chronic inflammation. The immune microenvironment, particularly the role of macrophages, is critical to bone healing and regeneration. Macrophages, essential components of the immune system, perform diverse functions, including pathogen clearance, inflammation resolution, and tissue repair.1,2 Depending on environmental cues, they polarize into either pro-inflammatory (M1) or pro-repair (M2) phenotypes. While M1 macrophages play a role in immune defense, M2 macrophages promote tissue regeneration, making their balance pivotal for effective bone healing. 3 Excessive reactive oxygen species (ROS), however, disrupt this balance, inducing OS, exacerbating inflammation, and impairing macrophage polarization, which hinders bone repair. 4 Modulating ROS levels has thus emerged as a promising strategy for enhancing bone regeneration.
ROS, while essential for normal cellular signaling and immune responses, becomes detrimental when dysregulated under pathological bone conditions such as diabetes or chronic inflammation. Elevated ROS levels induce OS and inflammatory responses, disrupting cellular homeostasis and impeding the healing process. Advanced biomaterials capable of scavenging ROS have shown great potential in addressing these challenges. These materials not only neutralize oxidative damage but also regulate the immune microenvironment, promoting effective bone repair.
To mitigate ROS-induced damage, both enzyme-based and nonenzymatic antioxidant materials have been developed. Metal-based nanozymes, such as cerium (Ce) oxide, Prussian blue, platinum (Pt), and manganese (Mn)-based nanomaterials, mimic endogenous antioxidant enzymes to scavenge ROS and modulate macrophage polarization as well.5,6 For example, Ce oxide nanozymes, leveraging reversible redox cycling, effectively reduce OS and promote M2 macrophage polarization.7,8 However, issues such as nanoparticle (NP) aggregation and degradation have driven the development of hybrid materials and doping strategies to enhance their stability and functionality. Similarly, bioenzyme-based materials, incorporating natural enzymes such as superoxide dismutase (SOD) and catalase (CAT), offer superior biosafety and specificity for ROS scavenging, with innovations such as Zeolitic Imidazolate Framework-8 (ZIF-8)-coated SOD achieving improved stability and targeted delivery.9,10
Nonenzymatic antioxidant materials, including ROS-responsive and ROS-scavenging nanomaterials, also provide promising solutions. These materials directly neutralize ROS and influence macrophage polarization to enhance tissue repair. In addition, targeted approaches involving organelle-specific materials, such as those directed at mitochondria or the endoplasmic reticulum (ER), further improve therapeutic precision by reducing intracellular ROS levels and minimizing off-target effects. While previous reviews have summarized the application of antioxidant materials for treating bone-related diseases,11,12 several critical limitations remain unaddressed. These reviews primarily treat OS as an isolated variable, failing to elucidate how antioxidants promote osteogenic repair through the spatiotemporal coupling mechanism within the immune-osteogenic network. Crucially, they neglect the pivotal role of immune cell metabolic reprogramming. As highlighted by the recent study of Lin et al., antioxidant materials can modulate macrophage reprogramming via mitochondrial metabolism, thereby reactivating bone marrow-derived mesenchymal stem cells (BMSCs) to enhance osteogenic repair in diabetic mandibular defects.13,14 Furthermore, existing reviews omit discussion on the potential of synergistic antioxidant–antibacterial materials for the integrated treatment of bone infection and regeneration. They also fail to address the key challenges facing the future development and clinical translation of antioxidant materials for managing osteo-inflammatory diseases.
This review highlights recent advancements in antioxidant materials for bone regeneration, focusing on (1) enzyme-based and (2) nonenzymatic materials that enhance therapeutic efficacy. By addressing OS and modulating the immune microenvironment, these materials offer innovative solutions for bone tissue engineering. Future research should focus on optimizing their design and biocompatibility to ensure successful clinical application in treating bone-related inflammatory diseases.
OS-Mediated Pathogenesis of Osteopathology
ROS are normal by-products of cellular metabolism. Pathologically, excessive ROS induces OS, which induces osteopathology through three principal mechanisms as follows: BMSC dysregulation, osteoblast death, and osteoclast hyperactivation (Fig. 1).

Relationship between the balance of ROS and bone diseases. The balance between the generation of ROS and the effect of intracellular antioxidant enzymes is crucial for the homeostasis of the body. Oxidative stress occurs when the production of ROS exceeds the antioxidant capacity. This leads to bone diseases, which trigger the downregulation of BMSCs, death of osteoblasts, upregulation of osteoclasts, and reduction of osteocytes. Created by FigDraw. BMSCs, bone marrow-derived mesenchymal stem cells; ROS, reactive oxygen species; Keap1/Nrf2 axis, Kelch-like ECH-associated protein 1/Nuclear factor erythroid 2-related factor 2 axis; HO-1, heme oxygenase-1; SODs, superoxide dismutases; SIRTs, sirtuins.
OS triggers dysregulation of BMSC growth and differentiation
Interfering with the differentiation and growth of BMSCs is considered to be one of the effective ways due to its importance for bone growth and healing (Fig. 2). 15 When mitochondrial function is impaired, the PTEN induced kinase 1 (PINK1) -Parkin pathway is activated and ultimately increases the activity of E3 ubiquitin ligase, which leads to mitochondrial membrane protein ubiquitination and mitophagy. However, upregulation of protein 53 (P53) triggered by OS blocks the mitochondrial translocation of Parkin, thereby inhibiting the process of mitophagy, leading to intracellular accumulation of damaged mitochondria and apoptosis of BMSCs.16,17 Nonetheless, it should be noted that the effects of P53 on BMSCs are complicated. TP53INP2, a downstream target gene of P53, can promote osteogenic differentiation of BMSCs by activating the Wnt/β-catenin pathway. Under OS, TP53INP2 is degraded through the autophagy–lysosome system, resulting in the inhibition of osteogenic differentiation of BMSCs. 18 Some experiments have also shown that under the condition of OS, mitophagy is induced in BMSCs through c-Jun N-terminal kinase (JNK) in the early stage as a cell self-protection mechanism, whereas long-term OS inhibits JNK, thereby shutting down mitophagy and promoting apoptosis through the activation of caspase.
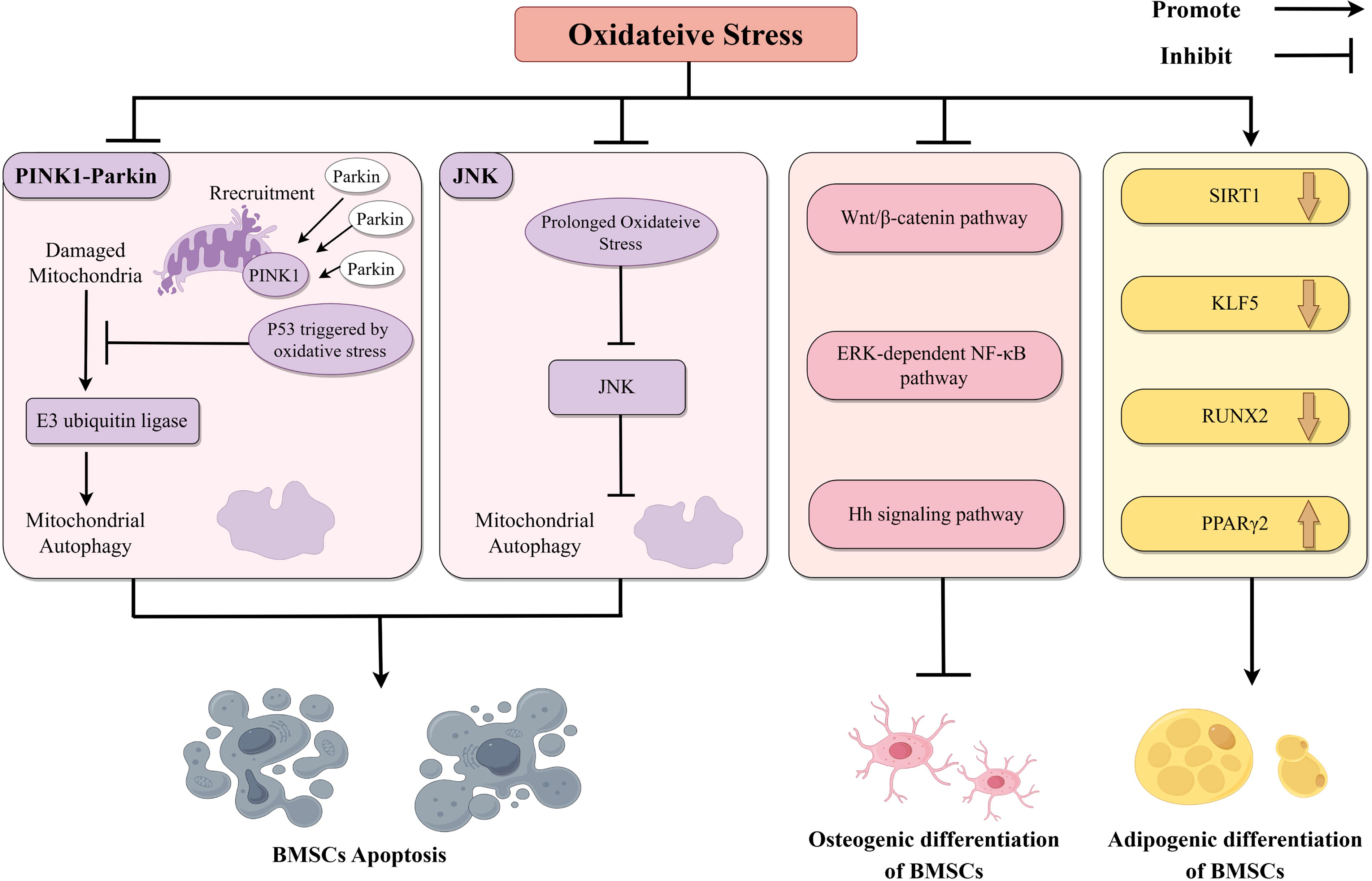
Oxidative stress mediates bone diseases by interfering with the growth and differentiation of BMSCs. One of the ways that oxidative stress can lead to bone disease is by affecting BMSCs. The mechanisms include promoting the apoptosis of BMSCs, inhibiting the osteogenic differentiation of BMSCs, and promoting the adipogenic differentiation of BMSCs. Created by FigDraw. Hh signaling pathway, hedgehog signaling pathway.
In addition, the increase of ROS diverted the limited β-catenin to FoxO-mediated transcription instead of the T-cell factor/lymphoid enhancer factor (TCF/LEF) pathway, thus inhibiting the Wnt/β-catenin pathway-mediated osteoblast differentiation.19,20 Extracellular signal-regulated kinase (ERK) and ERK-dependent nuclear factor kappa-B (NF-κB) pathway and Hedgehog pathway have also proved to be involved in OS-mediated inhibition of osteoblast differentiation.21,22 The effect of OS on BMSCs is also reflected in the change of differentiation direction. Excessive ROS inhibits the expression of sirtuin 1 (SIRT1) and Kruppel-like factor 5 (KLF5), which increases peroxisome proliferator-activated receptor gamma 2 (PPARγ2) expression and decreases runt-related transcription factor 2 (RUNX2) expression and ultimately leads to a tendency of BMSCs to differentiate into adipocytes instead of osteoblasts. 23
OS induces death of osteoblasts
The promotion of osteoblast death has also been considered an effective way of ROS-induced bone destruction. OPA1 cleavage, which may be regulated by the AKT-GSK3β signaling pathway, contributes to osteoblast apoptosis by promoting OS-induced mitochondrial dysfunction. 24 Moreover, in a recent study investigating bone loss induced by glucocorticoids, dexamethasone (DEX) has been shown to elevate the levels of ROS, thereby inhibiting the activation of the phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) pathway. This leads to an increase in the expression of glycogen synthase kinase 3 beta (GSK3β), which ultimately promotes the apoptosis of osteoblasts. 25 Except for apoptosis, ROS can also be induced by iron overload in osteoblasts and are capable of promoting programmed cell death, partly through the receptor-interacting protein kinase 1 / receptor-interacting protein kinase 3 / mixed lineage kinase domain-like protein (RIPK1/RIPK3/MLKL) pathway. 26
OS promotes maturation of osteoclasts
Osteoclasts have the function of bone resorption. Therefore, their activation is closely related not only to the process of bone remodeling but also the development of some bone diseases. 27 Studies have shown that ROS plays an important role in the process of osteoclast differentiation. 28 The combination of receptor activator of NF-κB ligand (RANKL) and RANK leads to the activation of Rac1 and nicotinamide adenine dinucleotide phosphate (reduced form) (NADPH) oxidase (Nox), 29 which then elevates intracellular ROS levels. 30 Among them, NOX 1, NOX2, and NOX4 have been proven to be involved in this process.31–33 As an intracellular second messenger, ROS can activate the mitogen-activated protein kinase (MAPK) cascade (including ERK, p38-MAPK, and JNK), which induces the translocation of the downstream signals c-Fos and NFATc1 and promotes the expression of various osteoclast differentiation genes. 34 Furthermore, ROS are capable of activating NF-κB through the MAPK signaling cascade, which in turn upregulates the NOD-like receptor family pyrin domain containing 3 (NLRP3) inflammasome and fosters osteoclast maturation. 35 lipopolysaccharide (LPS)-induced, ROS-dependent osteoclastogenesis may originate from the activation of JNK and signal transducer and activator of transcription 3 (STAT3) and is negatively regulated by peroxiredoxin (Prx) II. 36 The research on MAPK-related signaling pathways is comprehensive and thus has been widely used as targets for drug development.
Beyond the MAPK-centric activation mechanisms, ROS can promote the release of calcium ion (Ca2+) from the inositol 1,4,5-trisphosphate pool and the stromal interaction molecule (STIM) -mediated Ca2+ influx, which consequently activates phospholipase Cγ1 and results in Ca2+ oscillations. 37 The sustained oscillation of Ca2+ concentration is considered to be crucial in the maturation of osteoclasts induced by RANKL and can be regulated through transmembrane protein 64, which relies on the activity of sarcoplasmic/ER calcium ATPase 2. 38
Except for its role as secondary messengers, ROS are implicated in the regulatory control of RANKL expression. For murine osteoblastic cells, ROS have been demonstrated to induce the phosphorylation of the cAMP response element-binding protein and activating transcription factor 2 (ATF2) complex, resulting in their binding to the cAMP response element domain of the RANKL promoter. Furthermore, the promotion of RANKL expression by ROS is also attributed to the phosphorylation of ERK. Notably, in the human osteosarcoma cell line MG63, ROS mediates the activation of the RANKL promoter through heat shock factor 2 (HSF2). 39
ROS is also helpful in the maintenance of osteoclast function. For mature osteoclasts, ROS serve to bolster their functional sustenance. ROS facilitate the conjugation and oxidation of Src homology region 2 domain-containing phosphatase-1 (SHP-1) and cellular Src proto-oncogene tyrosine kinase (c-Src), culminating in the inactivation of SHP-1 and the activation of c-Src. And c-Src is indispensable for maintaining the functions of osteoclasts. 40
Functional Alterations of Macrophages Under OS Conditions
The functional alterations of macrophages under OS influence tissue regeneration and pathological progression. Specifically, macrophages exhibit a two-way response to ROS. While ROS play key roles in driving macrophage recruitment, activation, and polarization, they retain the capacity to induce macrophage cell death.
Mediation of macrophage recruitment by ROS
OS within the tissue can mediate the recruitment of macrophages to initiate subsequent responses. In tumor tissues, ROS can promote the production of cytokines (colony-stimulating factor 1 [CSF-1], chemokine (C-C motif) ligand 7 [CCL7], interleukin [IL8], vascular endothelial growth factor C [VEGF-C]) through the PI3K/Akt signaling pathway, thereby recruiting M2 macrophages. 41 In addition, the combined effort of ROS and matrix metalloproteinase 2 (MMP-2) contributes to the damage to the basement membrane, which facilitates the migration of macrophages into tumor tissues. 42 For epithelial cells, the assembly of their microRNAs (miRNAs) is influenced by OS. Specifically, OS mediates the O-GlcNAcylation of heterogeneous nuclear ribonucleoprotein A2/B1 (hnRNPA2B1), which allows it to bind to specific miRNAs. When released through extracellular vesicles (EVs), they are capable of recruiting and activating macrophages, resulting in an innate immune response. 43
Promotion of macrophage activation and function by ROS as intra/extracellular signaling molecules
Owing to its role as a signaling molecule, ROS plays a positive role in the activation and function of macrophages. For extracellular ROS, the increased production of extracellular ROS induced by malaria can directly promote the generation of pro-IL-1β in macrophages, which provides one of the signals for the activation of the NLRP3 inflammasome. This lays the foundation for the release of cytokines by macrophages. 44 Furthermore, ROS can also act indirectly on macrophages. Cytoplasmic hydroxyl radical is one of the most active forms of ROS. It generates oxidized phospholipids in the liver and kidneys, thus activating the NF-κB signaling pathway by binding to Toll-like receptor 4 (TLR4). This leads to the activation of macrophages, mediating apoptosis and fibrosis of the aging tissues. 45 However, the positive effects of ROS on macrophages rely on the optimal concentration of ROS. Otherwise, inappropriate concentrations have been demonstrated to exert inhibitory effects on macrophages. 46
Within the cell, ROS act as a second messenger involved in macrophage activation, and the pathways mediated by ROS vary in different diseases. For instance, in the context of bacterial infections, the increase in the production of ROS within phagolysosomes mediates the activation of CD38, resulting in the upregulation of nicotinic acid adenine dinucleotide phosphate. The release of Ca2+ facilitates the nuclear translocation of transcription factor EB (TFEB) through protein phosphatase 3 (PPP3) /calcineurin and promotes the subsequent macrophage-mediated inflammatory response. 47 In sterile inflammation, such as abdominal aortic aneurysm, angiotensin type I receptor and ROS-dependent NLRP3 activation contribute to the release of cytokines from macrophages. 48 Considering tissue-specific macrophages, the signaling pathways involving ROS may differ due to the unique characteristics of both the macrophages and their tissue environment. For example, in microglia, LPS-induced ROS may be dependent on lipoxygenase-12/15 (LOX-12/15), but not cyclooxygenase (COX), which is more common in other macrophages.49–51
Modulation of macrophage polarization by ROS
Under the influence of different microenvironments, macrophages polarize into M1 and M2 phenotypes after activation, exhibiting varied characteristics and functions. The role of OS in promoting the polarization of M1 macrophages has been demonstrated in a large number of inflammation-related diseases. In addition, due to the lower expression of oxidation-associated enzymes and higher expression of the antioxidant system, M2 macrophages have a decreased ROS level compared with M1 macrophages. However, the polarization of M2 macrophages still requires an adequate amount of ROS for its sustenance. 52 ROS have also been shown to promote the M2 polarization of macrophages in specific diseases. In the case of asbestos-induced pulmonary fibrosis, macrophages upregulate copper (Cu) and zinc (Zn)-SOD levels, which in turn mediate the generation of hydrogen peroxide (H2O2). This process leads to a reduction in the expression of jmjd3 due to the mutation of a cysteine residue in STAT6, resulting in the macrophage polarization toward an M2 phenotype that accelerates the progression of fibrosis. 53 For lung cancer, ROS are involved in the polarization of M2 macrophages mediated by the low expression of Prx5, which indicates a poorer prognosis. 54 In conclusion, optimal levels of ROS maintain homeostasis, whereas dysregulation drives pathology in macrophage polarization.
Induction of macrophage death by ROS
Although OS plays a positive role in the recruitment, activation, and polarization of macrophages, it still retains the capacity to induce the death of macrophages, thus ensuring immune homeostasis. Specifically, mtROS makes the cysteine residues of gasdermin-D (Gsdmd) oxidized, which increases the cleavage of Gsdmd by caspase-1. This promotes the release of the N-terminal domain of Gsdmd and subsequently mediates the pyroptosis of macrophages. 55 In addition, OS is regarded as an effective way to maintain the balance between monocytes and macrophages. Because of its deficiency in DNA repair, monocytes are susceptible to ROS produced by themselves as well as ROS produced by macrophages. This susceptibility can weaken their ability to differentiate into macrophages and may even induce their death. In contrast, macrophages exhibit the opposite situation. These help to prevent the excessive generation of macrophages. 56
Pivotal Role of Macrophage Polarization in Stem Cell-Driven Osteogenesis
The functional significance of macrophage polarization in stem cell-driven osteogenesis cannot be simply summarized by the static categorical framework. 57 Accumulating evidence indicates that successful bone regeneration relies critically on the initial pro-inflammatory M1 phase, the timely phenotypic switch to M2 dominance, and mutual effect between macrophages and stem cells—as opposed to unidirectional regulation (Fig. 3).

Different polarization of macrophages in stem cell-driven osteogenesis. Macrophage polarization always plays different roles in stem cell-driven osteogenesis. M1 macrophages promote the recruitment of BMSCs through the secretion of cytokines. And M2 macrophages induce the osteogenic differentiation of BMSCs through cytokine secretion and EVs. Created by FigDraw. M1, pro-inflammatory; M2, pro-repair.
M1 macrophages are considered to facilitate the recruitment and migration of stem cells and increase the abundance of stem cells, thus benefiting the subsequent osteogenesis. In detail, due to its capacity to release inflammatory mediators (tumor necrosis factor-α [TNFα], IL-6, IL-1β), M1 macrophages can induce cytoskeletal rearrangement and cellular elongation in BMSCs, both of which are related to enhanced cell migration.58–60 However, the effect of M1 macrophages in the subsequent process of osteogenesis is likely to be controversial. Some experiments have shown that the COX-2-prostaglandin E2 (PGE2) pathway and the proteins secreted by M1 macrophages, such as oncostatin M (OSM), IL-6, and TNFα, are essential for the initiation of BMSCs-driven osteogenesis.61,62 Nonetheless, more people believed that M1 macrophages may exert a negative effect on bone formation. M1 macrophages can release EVs that are loaded with miR-155. These EVs downregulate the BMP signaling pathway, which hinders the osteogenic differentiation of BMSCs. 63 In addition, M1 macrophages have been demonstrated to alter the differentiation direction of BMSCs, steering them toward adipogenesis rather than osteogenesis. 64 Although it is still controversial whether M1 macrophages have a positive effect on the proliferation and differentiation of stem cells, it is undeniable that the initial pro-inflammatory process induced by M1 macrophages is necessary but transient for stem cell-driven osteogenesis. Compared with M1 macrophages, M2 macrophages play a more important role in the continuous stimulation of osteogenesis. Therefore, a timely transition from the M1 to M2 phenotype is quite essential, 65 and recent evidence demonstrates that forkhead box protein O1 (FoxO1) is associated with the phenotypic transition. 66
The positive effect of M2 macrophages in promoting the proliferation and differentiation of stem cells and facilitating osteogenesis is relatively well-established. On the one hand, M2 macrophages can release anti-inflammatory cytokines, which have been widely recognized and studied in promoting osteogenesis. The secretion of these anti-inflammatory cytokines creates an immunological microenvironment conducive to bone formation and can also directly or indirectly influence stem cell-mediated osteogenesis. 67 Transforming growth factor (TGF)-β1 recruits BMSCs through the Smad2/3 signaling pathway and binds to receptors on BMSCs, which mediate the activation of Smad3. This leads to the upregulation of the expression of osteogenesis-related genes (ALP, Col I, BMP2). 68
M2 macrophages may also secrete BMP2 to promote osteogenic differentiation of BMSCs. Specifically, under the stimulus of mechanical drawing, the Yes-associated protein in M2 macrophages is activated and translocates into the nucleus, which leads to the increased expression of bone morphogenetic protein 2 (BMP2). 69 interleukin-10 (IL-10), insulin-like growth factor 1 (IGF-1), and OSM have also been demonstrated to enhance the expression of osteogenic-related genes and promote the osteogenic differentiation of stem cells.59,70,71 Moreover, VEGF secreted by M2 macrophages induces angiogenesis, which supplies the material foundation required for bone formation. This contributes indirectly to the osteogenesis process. 72
On the other hand, M2 macrophages can also regulate osteogenic differentiation through EVs. miR-378a, which acts on the BMP pathway, is encapsulated in the EVs secreted by M2 macrophages. Once these EVs are received by BMSCs, the expression of osteogenesis-related genes in BMSCs is promoted. 63 M2 macrophages use exosomes, a subtype of EVs, to transport miR-142-3p into BMSCs that suffer from dysfunction due to radiation exposure. In BMSCs, miR-142-3p inhibits the expression of TGF-β1, leading to an increase in osteogenic differentiation and a reduction in fibrogenic differentiation of BMSCs. These indicate that exosomes secreted by M2 macrophages can reverse the differentiation imbalance of BMSCs induced by radiation. 73 Furthermore, exosomes carrying miR-690 can upregulate the expression of insulin receptor substrate-1 (IRS-1) and transcriptional co-activator with PDZ-binding motif (TAZ) in BMSCs, thus inhibiting adipogenic differentiation and promoting osteogenic differentiation of BMSCs. 74
In terms of the process of bone formation, there is an interaction between stem cells and macrophages, rather than the unidirectional influence of polarized macrophages on stem cells. The upregulation of KDM6B from BMSCs can activate the brain and muscle ARNT-like 1 (BMAL1) gene in macrophages. This leads to the inhibition of the TLR2/NF-κB signaling pathway, thereby reducing the ratio of M1 to M2 macrophages. The reduction in ratio can promote the osteogenic differentiation of BMSCs, which makes the whole process a positive feedback loop. 75 And recent research indicates that exosomes derived from BMSCs regulate tripartite motif-containing protein 25 (TRIM25) -mediated triggering receptor expressed on myeloid cells 1 (TREM1) degradation, consequently enhancing M2 macrophage polarization and osteogenic differentiation. 76 In addition, adipose-derived stem cells (ADSCs) are capable of promoting the M2 polarization of macrophages, thus facilitating the differentiation of ADSCs into osteoblasts. 77
In summary, further determining the contributions of macrophage subtypes in bone healing and implementing research targeting both M1-to-M2 phenotypic transition and bidirectional macrophage-stem cell cross talk may be critical for this field.
Antioxidant Biomaterials for Multidimensional Regulation of the Bone Immune Microenvironment
OS and the associated ROS are highly correlated with the pathological process of numerous bone-related diseases, particularly in the context of diabetic bone defect and rheumatoid arthritis (RA),78–84 which is known to exacerbate OS and impair bone healing. The cellular redox balance, maintained by the interplay between ROS production and antioxidant defense mechanisms, is crucial for the regulation of macrophage polarization, a process that significantly influences bone regeneration.66,85,86
In the bone microenvironment, OS can disrupt the delicate equilibrium between M1 and M2 macrophage phenotypes, leading to impaired bone healing and regeneration. 66 The M1 macrophages, characterized by high ROS production, contribute to tissue damage and inflammation, whereas M2 macrophages, with lower ROS levels, promote tissue repair and angiogenesis. 87 The role of ROS in macrophage polarization is multifaceted. On one hand, ROS can activate inflammatory signaling pathways, such as the NF-κB pathway, leading to the upregulation of pro-inflammatory cytokines and the promotion of M1 macrophage polarization. On the other hand, moderate levels of ROS can also stimulate the expression of anti-inflammatory cytokines and the polarization of M2 macrophages, which is essential for tissue repair.88,89
In OA, the M1/T helper type 1 cells (Th1) activation occurs within an inflammatory milieu dominated by TLR and interferon (IFN) signaling, which facilitates the production of a plethora of pro-inflammatory cytokines (namely, TNFα, IL-1, IL-12, IL-18, and IFNγ), chemokines, Nuclear receptor subfamily 1 group D member 1, and MMPs, leading to osteoclastogenesis, erosion, and progressive joint destruction. Conversely, the activation of the M2/T helper type 2 cells (Th2) response dictates the release of growth factors and cytokines involved in anti-inflammatory processes, including IL-4, IL-10, IL-13, and TGF-β, thereby mitigating inflammation.87,90
Moreover, the cross talk between macrophages and stem cells, particularly mesenchymal stem cells (MSCs), is critical for bone regeneration. 91 Macrophage-derived cytokines and growth factors can influence the differentiation and function of MSCs, promoting osteogenic differentiation and the formation of new bone tissue. In addition, EVs, such as exosomes, can transfer microRNAs and other bioactive molecules from macrophages to MSCs, modulating their behavior and enhancing bone regeneration.91,92
In this context, the development of biomaterials and nanomedicines has opened new avenues for the regulation of OS and ROS levels in the bone microenvironment. Smart biomaterials, such as pH-responsive hydrogels and redox-sensitive NPs, can be engineered to release antioxidants or anti-inflammatory agents in response to the local OS, thereby modulating macrophage polarization and promoting a pro-regenerative microenvironment.80,93–98 Furthermore, the use of tissue engineering strategies, including references,99–102 has shown promise in enhancing bone regeneration by providing a conducive microenvironment for the migration, proliferation, and differentiation of MSCs. The design of such biomaterials requires a deep understanding of the complex interactions between ROS, macrophages, and stem cells, as well as precise control over the delivery of bioactive molecules.
Nonenzymatic antioxidant materials
In the realm of bone regeneration, the equilibrium of ROS is crucial for maintaining cellular functions and facilitating tissue repair. However, under diabetic or other inflammatory pathological conditions, an excess of ROS can be produced, leading to cellular OS, exacerbated inflammatory responses, and imbalanced macrophage polarization, thereby hindering the bone healing process. Therefore, in addition to nanomaterials that carry antioxidant enzymes and metal nanozymes, the development of nonenzymatic antioxidant materials capable of effectively scavenging ROS is equally significant for promoting bone regeneration.
In recent years, researchers have designed various nanomaterials with ROS-responsive or scavenging capabilities, which not only eliminate excessive ROS but also promote bone tissue repair by modulating the polarization state of macrophages. This section reviews the high-quality research advancements in this field in recent years and will discuss two special types of materials—nanomaterials that directly scavenge ROS (Fig. 4) and antioxidant materials targeting specific organelles (Fig. 5).
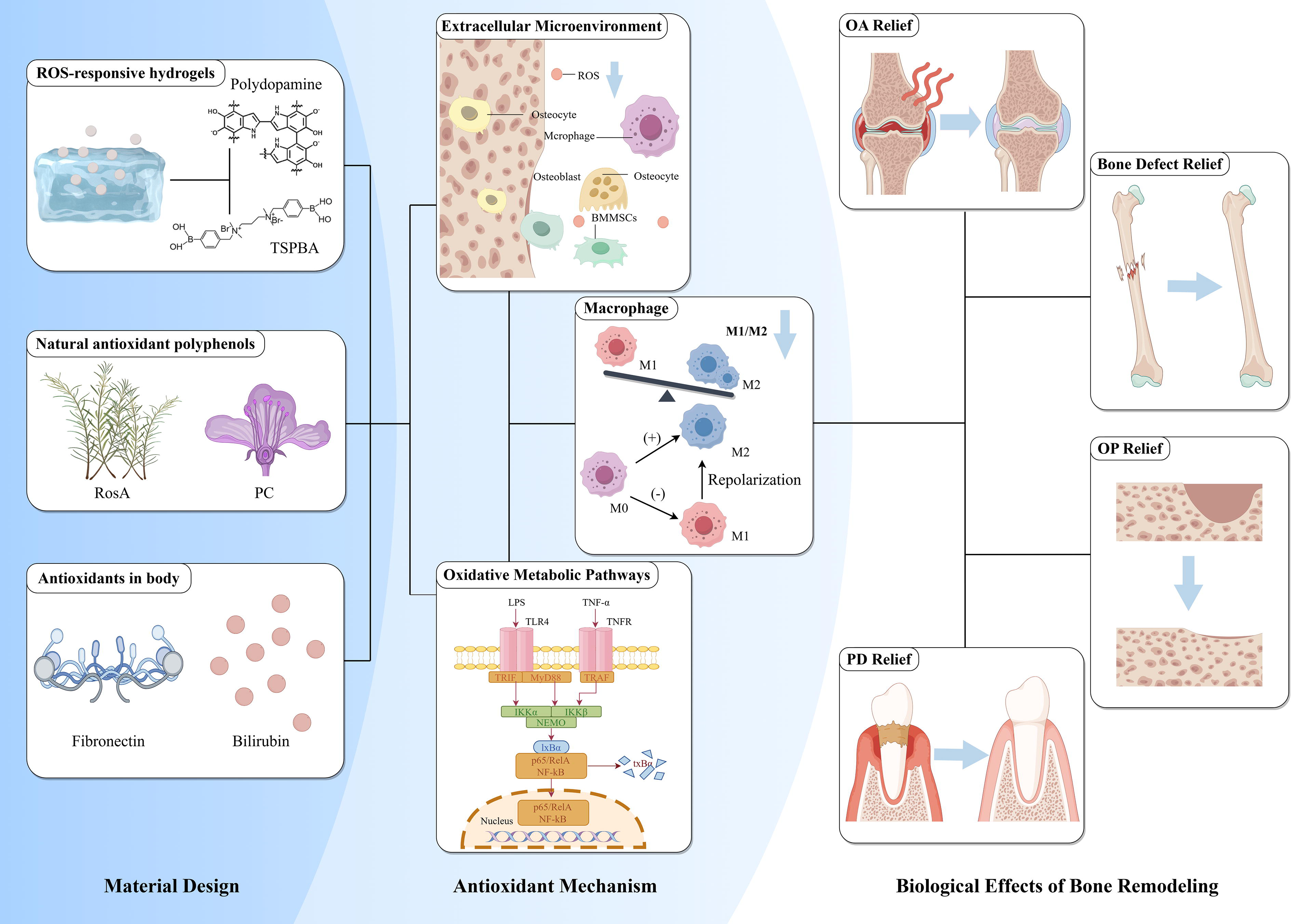
Mechanisms of bone regeneration mediated by nonenzymatic antioxidant biomaterials. Biomaterials with reactive oxygen species (ROS)-scavenging or ROS-responsive structures, those delivering natural antioxidants such as polyphenols, and those loaded with endogenous antioxidant proteins primarily promote bone regeneration by eliminating extracellular ROS and modulating intracellular oxidative metabolic signaling pathways. These mechanisms facilitate the polarization of pro-repair macrophages and, through interactions between immune cells and stem cells, ultimately achieve regeneration and repair of various bone inflammatory diseases. Created by FigDraw.
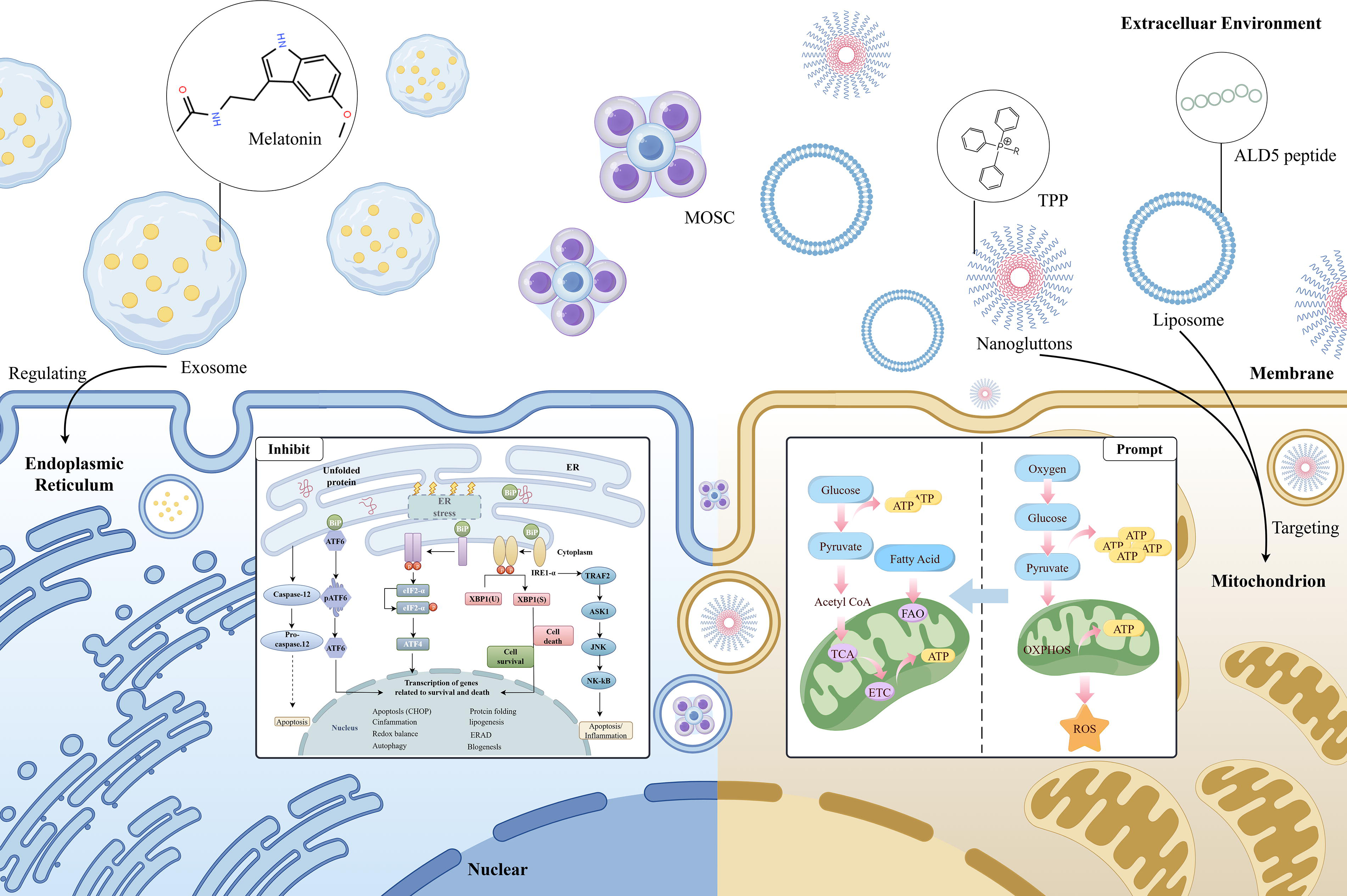
Mechanisms of organelle-targeted antioxidant nanomaterials. Nanomaterials functionalized with organelle-targeting peptides or other structures achieve metabolic reprogramming or regulation of the unfolded protein response in organelles such as mitochondria or the endoplasmic reticulum. These processes suppress oxidative stress, thereby contributing to their therapeutic effects. Created by FigDraw. TPP, triphenylphosphonium; MOSC, zinc-based metal-organic supercontainer; ALD5, aldehyde dehydrogenase 5.
ROS scavenging materials
ROS scavenging materials mitigate OS and inflammatory responses by directly reacting with ROS, reducing their accumulation within and outside cells. In recent years, a variety of nanomaterials have been developed for ROS scavenging due to their unique physicochemical properties. ROS-responsive biomaterials can intelligently control drug release in response to elevated ROS levels in the microenvironment, thereby consuming the ROS present. For instance, DEX nanomedicines stabilized with boric acid and assembled with poloxamer 188 have been shown to effectively suppress the production of ROS and nitric oxide in RAW264.7 macrophages through ROS-responsive drug release behavior and ROS scavenging capabilities. 98 ROS-responsive smart hydrogels are also a popular design direction; a logic-based diagnostic and therapeutic hydrogel has been reported with multistimulus responsiveness. The polyvinyl alcohol (PVA) network within the hydrogel reversibly breaks upon exposure to ROS or high glucose levels, thereby scavenging ROS. 83 Xiaoqing Lu et al. designed a bilayer hydrogel scaffold for cartilage defect repair, where the upper layer hydrogel containing polyvinyl alcohol crosslinked with phenylboronic acid degrades upon elevated ROS levels, releasing DEX, thus scavenging ROS and reprogramming macrophages. 103
Polydopamine (PDA), renowned for its abundant phenolic hydroxyl groups, exhibits exceptional antioxidant capabilities. PDA-mediated immunomodulatory patches and microneedles delivering PDA@Exo complexes have been ingeniously designed to leverage PDA’s antioxidant properties to scavenge ROS (Fig. 6b), mitigate inflammatory responses, modulate the secretion of pro-inflammatory and anti-inflammatory factors associated with the M1/M2 macrophage (Fig. 6c), and promote macrophage polarization toward the M2 phenotype by activating the PI3K-Akt-mechanistic target of rapamycin (mTOR) signaling pathway, thereby facilitating bone regeneration (Fig. 6a).81,104 In addition, a study has engineered dopamine nanoparticles (DAMM NPs) loaded with DEX, enhanced with arginine and Mn ions, to boost the particles’ antioxidant capacity and MRI signal, achieving sustained DEX release. This modulates the interaction between macrophages and chondrocytes, particularly through the TLR-3/NF-κB signaling pathway, to prevent chondrocyte apoptosis (Fig. 6f). 105 Yazhen Li et al. have also envisioned a DAMM-based conductive strategy, utilizing PDA to enhance the dispersion and stability of graphene oxide (PGO) and hydroxyapatite NPs within an alginate/gelatin network. PGO provides conductive pathways, while the scaffold possesses the ability to clear ROS and exhibits anti-inflammatory activity, inhibiting M1 macrophage polarization and activating M2 macrophages. It also mediates glycolysis and the Ras homolog family member A / Rho-associated coiled-coil containing protein kinase (RhoA/ROCK) pathway in macrophages to secrete osteogenic-related cytokines. 82

Characterization and in vitro experiments of nonenzymatic antioxidant materials that modulate macrophages to promote osteogenesis.
Furthermore, natural polyphenolic compounds have garnered widespread attention due to their potent antioxidant capabilities. These compounds are ubiquitous in plants, such as tea polyphenols, flavonoids, and proanthocyanidins, and they function by scavenging free radicals, inhibiting oxidative enzyme activity, and modulating intracellular antioxidant systems. Particularly in the treatment of inflammatory diseases, polyphenolic compounds not only clear excessive ROS but also modulate the polarization of immune cells, thus playing an anti-inflammatory role in the inflammatory microenvironment. A study has developed a multifunctional nanoplatform based on fifth-generation poly(amidoamine) dendrimers (G5 PAMAM) for codelivery of the antioxidant α-tocopherol succinate (α-TOS) and anti-inflammatory TNF-α small interfering RNA (siRNA) to macrophages. α-TOS reacts with oxygen radicals through its sixth hydroxyl group, producing a stable tocopherol semiquinone radical, thereby scavenging reactive oxygen. 106 Polyphenolic compounds such as rosmarinic acid (RosA) and procyanidins (PC) possess excellent ROS scavenging capabilities (Fig. 6d). Beilei Lu et al. have designed RosA NPs that clear ROS and promote the transition of macrophages from the M1 to M2 phenotype for the treatment of RA. 107 Furthermore, Piaoye Ming et al. have developed a biomimetic Se-nHA/PC composite microsphere, encapsulating selenoprotein-hydroxyapatite nanoparticles (Se-nHA NPs) and PC within a silk fibroin/sodium alginate (Se/SA) matrix using electrostatic injection technology. PC, through its internal reductive structure, namely the catechol group, can react with free radicals (OH-) via hydrogen atom transfer reactions, thereby scavenging ROS. 108
In addition to antioxidant enzymes within the human body, numerous endogenous substances possess antioxidant capabilities. The rational design and incorporation of these proteins can provide excellent antioxidant effects. Recently, a study has engineered a nanomedicine based on polymer NPs coated with fibronectin (FN), loaded with phosphodiester-terminated phosphoramidate dendrimers of the fourth generation (G4-TBP). This ROS-responsive nanomedicine is capable of drug release in an OS microenvironment, clearing excessive ROS through the antioxidant properties of G4-TBP and FN, thereby alleviating OS. 109 Two studies have designed a nanoplatform utilizing antioxidant properties of bilirubin (BR), using serum albumin-bilirubin-platinum nanoparticles (BSABR-Pt NPs) and immunoglobulin G-conjugated bilirubin/JPH203 self-assembled (IgG/BRJ) NPs to eliminate ROS. BR in BSABR-Pt NPs is used to eliminate ROS, whereas Pt exhibits a synergistic effect, clearing ROS and simultaneously generating oxygen. Ultimately, by inhibiting the hypoxia-inducible factor 1α (HIF-1α) pathway, BSABR-Pt NPs induce the repolarization of hypoxic M1-type macrophages to M2-type macrophages, modulating metabolic pathways from glycolysis to oxidative phosphorylation for the synergistic treatment of RA, by interrupting the harmful feedback loop between hypoxia and OS. 78 In contrast, internalized IgG/BRJ NPs decompose under increased OS, releasing BR and JPH203 to clear ROS, inhibiting the nuclear factor κ b pathway and the mammalian target of the rapamycin pathway, leading to macrophage repolarization and increasing the M2/M1 ratio (Fig. 6e). 110
Targeting macrophages with antioxidant NPs appears to be a more precise design direction. Shihui Li et al. have designed a multifunctional supramolecular integrated nanomaterial based on mannose-modified azocalix[4] arene (ManAC4A) and ginsenoside Rb1. ManAC4A possesses the ability to clear ROS, reducing OS through its calixarene structure reacting with ROS. 79 Introducing reductive gases to clear ROS is also an emerging design concept. A study has designed an injectable magnesium-loaded hydrogen gel (Mg@PEG-poly(lactic-co-glycolic acid) [PLGA]) for the repair of osteoporotic bone defects. Magnesium microparticles in the material react with water to produce hydrogen gas, used to clear ROS. Hydrogen gas reacts with highly cytotoxic hydroxyl radicals (·OH), alleviating OS, thereby inhibiting inflammation and promoting tissue repair. 93 However, how to control the production of hydrogen gas on-demand may warrant further consideration. In addition, inhibiting ROS production through dysfunction appears to be another design concept beyond clearing ROS. A study has designed a self-amplified immune modulation strategy based on a cytokine-ZIFs system for the treatment of tissue regeneration disorders in diabetes. Zn2+ released in the wound area not only enhances the antioxidant capacity of macrophages, preventing dysfunction caused by ROS but also upregulates the expression of IL-33 receptor (ST2L) and triggers the polarization of M2-type macrophages. 111
For such ROS-scavenging materials, the antioxidant components integrated into their structure or delivery system are pivotal for mediating their immunomodulatory and osteogenic effects. However, aligning with the current trends toward targeted and refined material design, elucidating the precise molecular mechanisms of these antioxidant components appears essential. Materials design must simultaneously amplify beneficial antioxidant effects while stringently controlling or circumventing potential adverse reactions. Thus, the deeper integration of multiomics sequencing technologies in biomaterials development is warranted to facilitate clinical translation. Furthermore, in the context of infected osteolytic diseases, ROS within the cellular microenvironment exhibits a dual role, possessing inherent antimicrobial activity. Consequently, mitigating potential increases in infection susceptibility resulting from ROS scavenging becomes a critical consideration in material design. Finally, establishing the physiological ROS concentration range within the target bone tissue is fundamental. Designing materials whose antioxidant activity terminates upon reaching this target concentration is also critical for preserving immune homeostasis in the bone.
Organelle-targeted materials
Organelle-targeted materials are designed to enter cells via specific pathways and target particular organelles, such as mitochondria or ER, to enhance antioxidant efficiency and minimize side effects on cells. Mitochondria are the primary source of ROS within cells. Ce6@M-Lip, through its mitochondria-targeting action (Fig. 7a), generates ROS upon laser irradiation, exacerbates mitochondrial dysfunction (Fig. 7b), induces apoptosis in pro-inflammatory macrophages (Fig. 7c), and thus serves in the treatment of RA. 112 In addition, a zinc-based metal-organic supercage, named Zn-NH-pyr, has been engineered. This material utilizes the -NH-pyr group in its structure as a proton donor and its electron-rich characteristics to provide active sites for ROS scavenging. Zn-NH-pyr captures abundant free radicals and brings them into close contact with the -NH active sites, preventing chain reactions initiated by highly reactive radicals by donating electrons. By activating the Nrf2 signaling pathway, it increases the production of antioxidants while inhibiting the NF-κB pathway, reducing the expression of inflammatory cytokines. 113 Another study designed a material termed “nanogluttons” (PAMAM-PEG-TPP [PPT]+BAPTAAM [PPTB] NPs), composed of polyamidoamine (PAMAM) and 1,2-Bis(2-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid tetrakis(acetoxymethyl ester) (BAPTA-AM), with a surface conjugated to polyethylene glycol (PEG) -TPP. PPTB NPs effectively control the sustained opening of mitochondrial permeability transition pores (mPTP) by “engulfing” or capturing Ca2+ around mitochondria, reducing the leakage of mitochondrial ROS into the cytoplasm. By regulating the sustained opening of mPTP, PPTB NPs decrease mitochondrial ROS production, block downstream inflammatory signaling pathways, and thus suppress the inflammatory activation of macrophages. In terms of biomedical effects, Zn-NH-pyr significantly improved joint inflammation, whereas PPTB NPs markedly reduced alveolar bone loss in a periodontitis model, demonstrating the potential for treating chronic inflammation-related diseases. 114
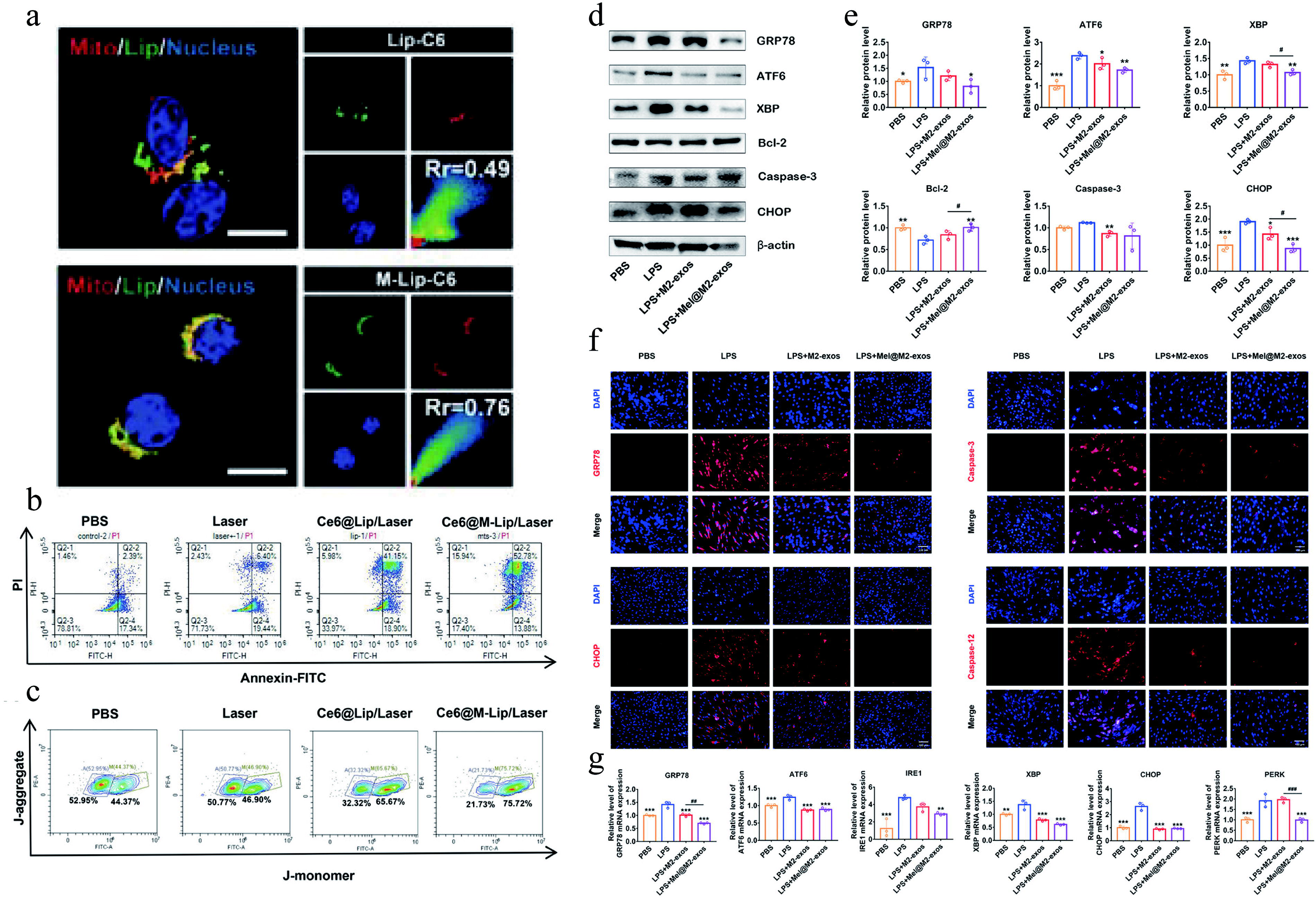
In vivo experiments of macrophage regulation by organelle-targeted antioxidant materials.
ER stress can lead to the unfolded protein response (UPR) and the generation of ROS. Mel@M2-exos, designed by Ya Cui et al., clears ROS by reducing excessive ER stress and UPR, promoting the transition of macrophages from the M1 to M2 phenotype for the treatment of periodontitis (Fig. 7d–g). 115
These materials, by targeting specific organelles, can more effectively scavenge ROS and modulate the intracellular environment, thereby promoting bone regeneration. With precise targeting, these materials can reduce interference with normal cellular functions and enhance therapeutic efficacy. However, current research primarily focuses on subcellular organelle targeting. Incorporating macrophage-specific targeting strategies based on their surface markers could potentially enable a more precise elucidation of the specific contributions of antioxidant activity to immune-osteogenic cross talk. In summary, ROS-scavenging materials and organelle-targeted materials show great potential in the field of bone regeneration biomaterials. These materials clear ROS and modulate macrophage polarization through different mechanisms, providing new therapeutic strategies for bone regeneration. Osteogenesis is a protracted process, consequently demanding sustained material functionality. This requirement inherently challenges both biocompatibility and controlled biodegradation kinetics over extended periods. Furthermore, facilitating the clinical translation of such materials necessitates the establishment of standardized regulatory frameworks and strategic road maps.
Enzyme-based materials
Nanozyme-based materials
Nanozymes, a class of nanomaterials with enzyme-like catalytic activity, are being widely developed due to their highly adjustable catalytic activity and low production costs. Certainly, they have also been applied to osteogenic regulation associated with macrophage polarization, serving as ROS scavengers.
Ce-based nanozymes, owing to their abundance of oxygen vacancies, have a reversible redox transformation between Ce4+ and Ce3+, thus enabling the elimination of ROS. However, the agglomeration of NPs and their susceptibility to degradation in vivo are challenges that need to be conquered in the development of Ce-based nanozymes. Combining Ce NPs with other biomaterials may be a viable solution. CeO2 and TGF-β3 modified mesoporous silica NPs have been incorporated into gelatin methacrylate (GelMA) and hyaluronic acid methacrylate (HAMA) to synthesize composite hydrogels. This hydrogel can solidify to repair the defective areas of the annulus fibrosus, and the embedded NPs can induce M2 macrophage polarization by scavenging ROS. 116 CeOX NPs and Rhein (RH) are conjugated to the hyaluronic acid (HA) backbone to form a ROS-responsive micelle (HA@RH-CeOX). It can be transported into cells safely, and excessive ROS within cells cleave the thioketal linker, thereby releasing RH and CeOX. CeOX reduces OS in M1 macrophages, whereas RH inhibits TLR4 signaling. The combined effort of both agents drives the M2 polarization of macrophages, which is conducive to cartilage repair. 8 Furthermore, doping with other metals has also been considered as a solution. Doping Zn into CeOX gives the material more oxygen vacancies and enhanced electron transfer capability. This endows the material with an enhanced ability to alleviate OS and an effective immunomodulatory in the OA microenvironment. 7
A recent study has developed a CeO2 nanozyme that targets the process of efferocytosis. The CeO2 nanozymes are coated with the membranes of apoptotic neutrophils (AM/CeO2), which is a biomimetic signal that initiates macrophage efferocytosis. Then CeO2 nanozymes can facilitate continuous efferocytosis by reducing the ROS levels within macrophages. Efferocytosis, in turn, promotes M2 polarization of macrophages. AM/CeO2 is loaded by a short fiber matrix, which enhances the biocompatibility of the material and provides more binding sites for macrophages (Fig. 8). 117 In addition, Ce-based nanozymes can be used as coatings on the surface of implants.118,119 For example, the chain armor structure composed of Ce and tannic acid (Ce-TA) can be applied to any implant surface and is capable of reducing mitoROS to inhibit HIF-1α. This shifts the macrophage phenotype to M2, which is beneficial for subsequent implant osseointegration. 119

Antioxidant nanozyme (short fiber sponges [PFS]@apoptosis-mimetic/cerium dioxide [AM/CeO2]) reprogram of the bone immune microenvironment to accelerate regeneration.
Prussian blue nanozymes, iron-based nanozymes, have also been extensively investigated in the treatment of diseases, including Alzheimer’s disease, inflammatory bowel disease, Parkinson’s disease, OA, and OP. Hollow Prussian blue nanozymes can improve the microenvironment of OP and inhibit bone resorption. This effect may be related to the reduction of intracellular ROS and the inhibition of NF-κB signaling pathway. 120 The biodegradable hollow-structured Mn Prussian blue nanozyme (HMPBzyme) eliminates ROS through its variable valence states, thereby inhibiting HIF-1α and inducing M2 polarization of synovial macrophages for the treatment of OA. Furthermore, the biodegradability of HMPBzyme enhances its biosafety, which makes it superior to other materials. 6 Similar to Ce-based nanozymes, the doping of other metals has also been used to enhance the performance of Prussian blue nanozymes. Due to the doping of Mn, the Prussian blue NPs have not only acquired the ability to eliminate various ROS and modulate immunity but also gained the capacity to directly promote osteogenesis, ultimately resulting a reduced bone loss mediated by periodontitis. 5
For Pt-based nanozymes, the development of Pt and selenium (Se) hybrid nanozymes has been proven to have enhanced ROS and reactive nitrogen species (RNS) scavenging capabilities, effectively promoting the polarization of synovial macrophages toward the M2 phenotype. 121 In addition, selecting a carrier with an appropriate structure for Pt-based nanozymes is crucial for enhancing the performance of the material. ZIF-8, known for its high specific surface area and porosity, allows for the introduction of lanthanum (La) and Pt to form a bimetallic metal-organic framework (MOF) (Pt@ZIF-8@La). While Pt scavenges ROS to modulate macrophage polarization and osteogenesis, both La and Zn (a product of the degradation of ZIF-8) promote osteogenic mineralization. 122 Graphitic carbon nitride (g-C3N4) possesses a stable N/C coordination network arising from its triazine ring structure, which facilitates the introduction of Pt. The lone-pair electrons of nitrogen of g-C3N4 can further enhance the catalytic activity of Pt. Therefore, Pt SA/C3N4 nanozyme can scavenge ROS and RNS, thereby normalizing the OA microenvironment. 123
In addition, Mn-based nanozymes have also been applied to macrophage polarization-related osteogenic regulation. Manganese dioxide-ferritin nanozyme (MF@S nanozyme) is susceptible to lysosomal internalization, so researchers modified the MF@S nanozyme with SS31, a cationic peptide that targets mitochondria. This modification allows the nanozyme to specifically deliver to the mitochondria of cells in the microenvironment of diabetic bone defect, thereby scavenging mitoROS, promoting M2 polarization of macrophages, and facilitating bone regeneration. 124 Mn porphyrin-based two-dimensional MOF nanosheets promote the polarization of M2 macrophages by facilitating the dismutation of O2− and H2O2, thereby inhibiting the apoptosis of BMSCs. Furthermore, the ultrathin two-dimensional structure allows for the exposure of as many active sites as possible, which enhances the catalytic activity. 125
Given that an appropriate amount of ROS is essential for maintaining some normal physiological activities and some type of ROS may specifically exist in disease microenvironments, some researchers have begun to develop specific ROS scavengers. Cu-N4ClG SAzymes possess SOD and CAT-like activities, which enable them to scavenge the superoxide anions commonly existing in the OA microenvironment. The MIL-47(V)-F (MVF) nanozyme, owing to its exposed vanadium sites with high reactive activity, exhibits glutathione peroxidase-like activity to specifically eliminate H2O2, which is the major ROS. This allows this material to eliminate ROS moderately. 126
Bioenzyme-based materials
Although metal nanozymes have achieved a significant position in this field, their application is still limited by their potential toxicity concerns. 127 Fortunately, bioenzyme-based materials show better advantages and have been developed (Fig. 9). A hydrogel incorporating zippered G4-Hemin DNAzyme has been fabricated through the rolling circle amplification technique. The DNAzyme in the hydrogel emulates the catalytic structure of horseradish peroxidase, thereby exhibiting efficient activity of scavenging ROS within the microenvironment of OP. 10
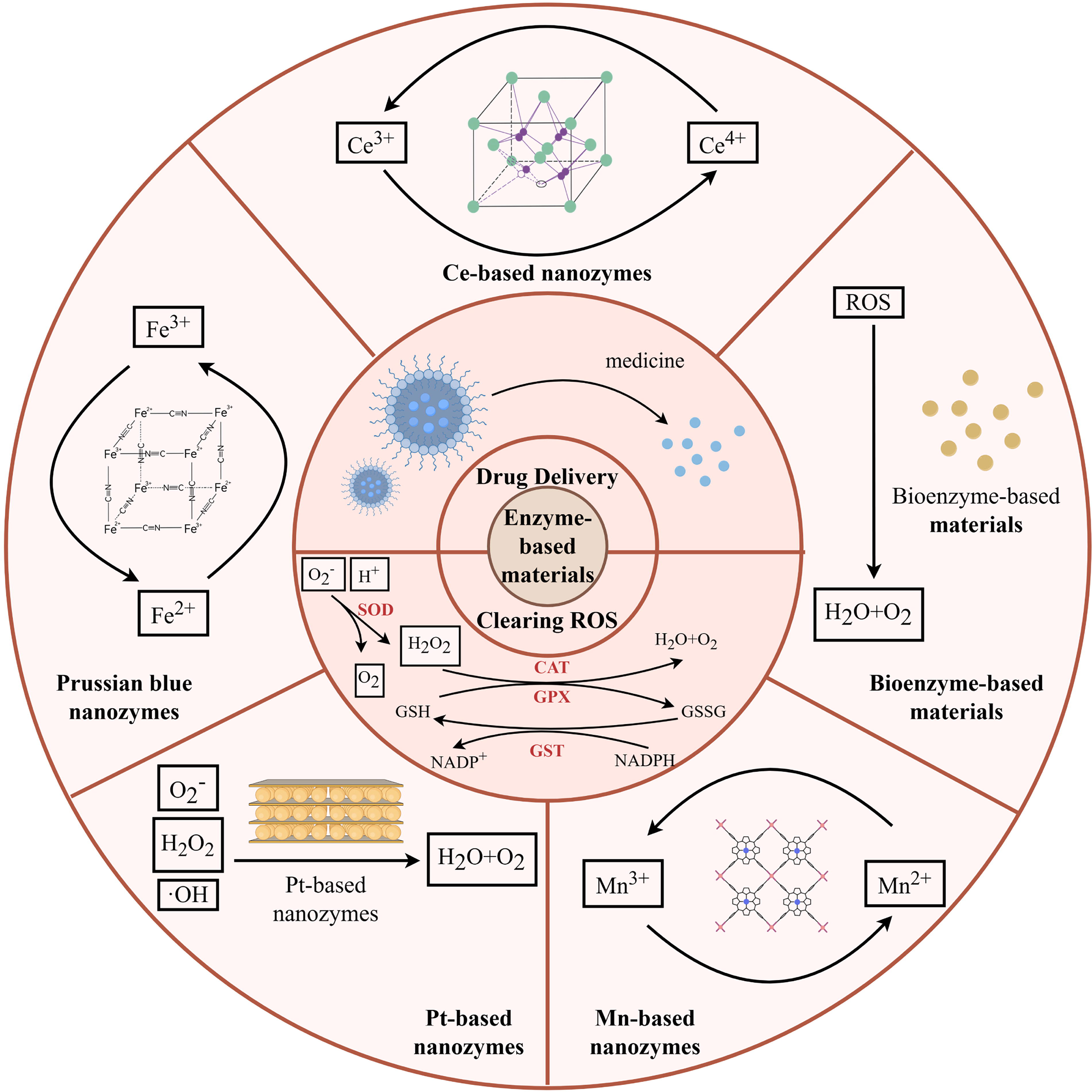
The role of enzyme-based materials in modulating macrophage polarization via oxidative stress regulation for enhanced osteogenesis. A variety of enzyme-based materials have been used in osteogenesis enhancement via oxidative stress-mediated macrophage polarization, such as Ce-based nanozymes, Prussian blue nanozymes, Pt-based nanozymes, Mn-based nanozymes, and bioenzyme-based materials. Their different compositions result in diverse mechanisms of action and material characteristics. But the primary modes of action of these materials are drug delivery and direct ROS scavenging. Created by FigDraw. Mn, manganese; Pt, platinum.
However, how to ensure that these bioenzymes are safely delivered to the target site and exert effective action is regarded as a problem that bioenzyme-based materials must overcome. The ultrathin ZIF-8-coated SOD nanoformulation (utZIF-SOD) achieves better cellular uptake and escapes from being degraded through the small size effect. Furthermore, the ultrathin ZIF-8 coating gives the material enhanced stability. Consequently, utZIF-SOD shows a greater ROS scavenging capacity compared with native SOD, thereby effectively reversing age-related OP. 9 In addition, the use of TA-functionalized polymer capsules to deliver CAT has been proven to reduce OS in the nucleus pulposus. 128
A key advantage of metal-containing nanozymes for treating osteo-inflammatory diseases lies in the potential of their constituent metal ions, following antioxidant activity, to further potentiate osteogenic differentiation of MSCs. Consequently, selecting metal ions necessitates a dual emphasis on high osteoinductive potency and minimal cytotoxicity. This rationale provides a compelling basis for designing dual-phase intelligent antioxidant nanomaterials capable of sequentially exerting antioxidant effects followed by robust osteogenic promotion. Furthermore, a critical design imperative for both metallic nanozymes and biological enzymes is ensuring their intact structural and functional delivery to the target pathological site. Beyond MOF structures and graphene-based carriers investigated in this section, the membrane-derived carriers discussed earlier, which offer targeting capabilities toward immune cells and organelles, represent another promising class of delivery vehicles.
Conclusion
Bone tissue repair and regeneration is a complex process involving the participation of various cells and is regulated by multiple factors. The immune system plays a crucial role in maintaining the balance between bone formation and resorption, and OS can disrupt this delicate equilibrium, leading to impaired bone healing and regeneration. Therefore, interventions aimed at modulating OS to enhance osteogenesis represent a promising therapeutic strategy for promoting functional bone tissue regeneration.
Although we have previously discussed the involvement of OS in the development of bone disease and its potential to promote osteogenesis through various mechanisms, there are still many potential mechanisms to explore. For instance, M2 macrophages are widely recognized for their role in tissue repair and regeneration. However, during the inflammatory phase, excessive infiltration of M2 macrophages can impair the tissue’s ability to resist foreign pathogens, potentially compromising tissue healing. The mechanisms underlying this dual function of M2 macrophages have not been well characterized. Thus, controlling the appropriate extent of M2 polarization remains a critical area of focus.
Despite the advancements, existing research faces challenges such as the specificity of macrophage polarization, the biocompatibility of materials, and the long-term stability of the redox state in the microenvironment. Potential solutions may lie in the development of smart biomaterials that can respond to environmental cues, advanced fabrication techniques for better control over material properties, and a deeper understanding of the cross talk between macrophages and stem cells.
The future of research in this field is likely to involve the convergence of nanotechnology, biomaterials, and cellular biology. The development of smart biomaterials that can dynamically respond to the local environment and effectively modulate macrophage polarization represents an exciting avenue. Integrating antioxidant capabilities into electromagnetic biomaterials represents an innovative strategy to enable real-time quantification of ROS within tissues, based on which these materials could autonomously modulate antioxidant activity in response to local ROS concentration. In addition, the exploration of novel pathways and targets for modulating OS, such as the role of microRNAs and long noncoding RNAs, could offer new therapeutic opportunities. Furthermore, the integration of systems biology approaches to understand the future of research in this field may involve the integration of nanotechnology for targeted delivery, the use of systems biology approaches to understand the complex interactions between macrophages, stem cells, and the bone microenvironment, and the incorporation of machine learning for the prediction of material performance. This includes the use of computational models to predict the behavior of these cellular interactions under various conditions and the development of high-throughput screening methods to identify effective modulators of macrophage polarization. Otherwise, the exploration of the role of the microbiome in bone regeneration and the development of biomaterials that can harness this interaction present exciting avenues for future research. This review also has inherent limitations. Our analysis focused comprehensively on macrophages; however, neutrophils represent a significant cellular source of ROS and exhibit persistent infiltration in diverse osteo-inflammatory diseases. A systematic investigation into how antioxidant materials modulate the interplay between neutrophils and stem cells remains an important area for future research.
Finally, the translation of these findings into clinical practice will require rigorous testing and validation. This includes the development of standardized protocols for the assessment of macrophage polarization in clinical samples and the evaluation of the safety and efficacy of novel biomaterials and compounds in preclinical and clinical trials. The modulation of the redox state in the extracellular microenvironment and the regulation of intracellular oxidative metabolism signaling pathways are critical for macrophage M2 polarization and stem cell interactions, essential for bone regeneration in various diseases. The development of biomaterials that can effectively regulate these processes offers a promising therapeutic strategy for bone repair and regeneration.
Authors’ Contributions
Z.M.: Conceptualization (lead), investigation (lead), and writing—original draft (lead). Y.Z.: Investigation (lead) and writing—original draft (lead). Y.M.: Visualization (lead). X.L.: Writing—review and editing (supporting) and funding acquisition (supporting). H.L.: Writing—original draft(supporting). H.Liu.: Conceptualization (lead), supervision (lead), and funding acquisition (lead). J.L.: Writing—review and editing (lead) and funding acquisition (lead).
Footnotes
Author Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This article was supported by grants from the National Natural Science Foundation of China 82401072 and 82170934, Natural Science Foundation of Sichuan Province 2023NSFSC1512 and 24NSFSC0536, and Sichuan Science and Technology Program (China) (2024YFFK0201).
Declaration of Generative Artificial Intelligence (AI) in Scientific Writing
During the preparation of this work, the ChatGPT-4o was used in the preparation of this article to improve language and readability. The author(s) have composed, reviewed, and edited the content manually, taking full responsibility for the content of the publication.
Data Availability
No data were used for the research described in the article.