
Abstract
Bone is a common site of breast cancer metastasis, which dramatically increases fracture risk. Recent in vivo bone metastasis studies show that mechanical loading is osteoprotective; however, little is known about how loading regulates breast cancer cell function in the unique bone mechanical environment, especially in combination with radiotherapy, one of the first-line treatments for advanced breast cancer patients. Here, we characterize the breast cancer cell response to a range of bone-mimicking fluid shears and determine how irradiation further modulates one candidate gene: SERPINE1. We found that irradiation, regardless of dosage, modulates SERPINE1 expression and is sensitive to the timing of administration. Additionally, protein expression of SERPINE1 accompanies a protumorigenic gene expression profile, which is elevated with higher-magnitude fluid shear stresses in a bone-mimicking 3D environment. Thus, we postulate that plasminogen activator inhibitor-1 (PAI-1) (encoded by SERPINE1) is a critical growth factor contributing to osteolytic lesion development in the bone metastatic vicious cycle of breast cancer.
Impact Statement
Research on the bone metastatic vicious cycle frequently investigates how osteocytes and other cells involved in bone remodeling impact breast cancer cells, but the downstream impacts of the breast cancer cell response to the bone mechanical environment are understudied. This work is one of the few studies investigating how breast cancer cells respond to a bone-like fluid mechanical environment and will facilitate our understanding of the breast cancer bone metastatic vicious cycle. Additionally, to our knowledge, this is the first study that has examined how the timing of ionizing irradiation administration influences the breast cancer cell mechanoresponse in a 3D model. Mechanosensation and the cell mechanoresponse have largely been studied in 2D models, and an understanding of the physiologically relevant 3D environment is crucial to elucidating complex cell–extracellular matrix interactions and responses to damaging stimuli such as ionizing radiation.
Introduction
Over 70% of advanced breast cancer patients are diagnosed with incurable skeletal metastasis.1,2 Breast cancer that metastasizes to bone is generally osteolytic, destroying bone in a feed-forward vicious cycle that concurrently promotes tumor growth and bone resorption. 3 Of all the breast cancer subtypes, triple-negative breast cancer (TNBC) has the lowest rate of bone metastases, but is the most aggressive subtype and has the lowest 5-year survival rate once bone metastasis occurs.1,4 Due to localized osteolytic lesions, bone metastases require a more targeted treatment approach. 5 Radiotherapy is one such frontline treatment modality, providing both local tumor control and palliation of skeletal pain. 6 For TNBC, radiation therapy is a critical intervention, since standard treatments targeting estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) are ineffective. 7 Radiation induces DNA double-strand breaks, leading to TNBC cell death.7,8 However, advanced TNBC is particularly resistant to most therapies, especially radiation, and requires treatment combinations to improve the effectiveness of local therapies.6,9,10
Exercise is one nonpharmacological intervention that has been rising to prominence as part of treatment strategies for cancer. 11 Physical activity is especially critical to maintaining bone strength. In healthy bone, mechanical stimuli are a well-documented anabolic signal for bone,12–15 but in bone metastasis, the skeleton’s response to these signals is disrupted. Recent in vivo studies have shown that increased mechanical loading (i.e., an analog for exercise) has antitumorigenic effects on bone metastasis models in mice.16–21 However, the skeleton’s anabolic response to loading is not completely protected in these models,16,18 suggesting that metastasis interferes with the skeleton’s ability to functionally adapt to mechanical loads.
The skeleton, as a mechanically active organ, maintains homeostasis via the osteocyte, the primary mechanosensitive cell in bone that senses and responds to mechanical stimuli.22–24 Osteocytes respond to force-induced interstitial fluid flow, the central mechanical signal leading to net bone formation. 24 Our previous in vitro studies revealed that osteocytes lost their mechanosensitivity when cultured with breast cancer cell conditioned media, pointing to a potential mechanism underlying the muted response to loading in vivo.25,26 Furthermore, when breast cancer cells were also loaded, their downstream effects on the mechanoresponse of mesenchymal stem cells 27 and osteocytes25,28 were modulated. Specifically, conditioned media from loaded breast cancer cells promoted a positive mechanoresponse from these osteoblast-lineage cells. However, none of these studies identified molecular mediators.
After metastatic colonization, cancer cells arrive in the bone marrow and often migrate to the endosteal surface. Fluid flow within the marrow and along the bone surfaces is highly variable, ranging from as low as 0.02 Pa to as high as 8 Pa based on computational estimates ex vivo.29–33 This, combined with the increased tissue stiffness of bone, anywhere between 1 and 10 GPa, compared to the stiffness of tumors in the breast tissue on the order of 1–10 kPa, makes the skeleton a far more mechanically complex and dynamic environment for cancer cells.34–37 The response of breast cancer cells to mechanical stimuli has been well documented in vitro, where a larger stimulus (e.g., substrate stiffness, increased fluid flow) correlates with increased migration and proliferation.38–41 However, most of these studies were not conducted in a bone-mimicking microenvironment, nor did they account for the unique micromechanics of that environment. In a previous study from our group, breast cancer cell viability was unaffected after undergoing mechanical loading in the bone microenvironment, but expression of bone remodeling genes was mechanically regulated. 16 We reasoned that the breast cancer cell response to mechanical stimuli may undermine the osteoprotective effects of increased loading. However, there are few studies characterizing the comprehensive breast cancer cell response to a bone-mimicking and mechanically similar microenvironment.
In this study, we investigated the response of TNBC cells to physiologically relevant, bone-mimicking fluid shear stresses. To address how mechanical loads may modulate TNBC resistance to common therapies like radiation, we also determined how irradiation alters the TNBC cell transcriptome. We identified several genes that may be responsible for abrogating the effects of loading, including SERPINE1. SERPINE1 encodes the serine protease inhibitor plasminogen activator inhibitor-1 (PAI-1), 42 and our particular interest in SERPINE1 was due to its overexpression in TNBC,43,44 its association with breast cancer cell therapeutic resistance,45,46 and its critical role in managing bone homeostasis via fibroblast growth factor 23 (FGF23) regulation and its transcriptional activation in the transforming growth factor beta (TGF-β) signaling pathway in osteocytes.42,47 Here, we show that irradiation, regardless of dose, shifts breast cancer cell gene expression in a timing-dependent manner, and that protein-level upregulation of many such genes accompanies a protumorigenic transcriptional profile under higher shear magnitudes in a 3D bone-mimetic culture system.
Methods
Bone-mimetic scaffold fabrication
Scaffolds were fabricated using a mixture of 152 mg of sieved NaCl, 8 mg of hydroxyapatite crystals, and 8 mg of poly(lactide-coglycolide) microspheres. 48 This mixture forms an 8mm diameter, 2 mm height structure after undergoing 2 metric tons of pressure delivered by a manual hydraulic press (Carver Press). A subsequent gas-foaming and salt-leaching procedure formed the final scaffold by revealing interconnected pores of 250–450 μm in diameter, resulting in an overall porosity of 70–90%. 49 Previous work on these scaffolds has confirmed that, while they do not mimic trabecular bone tissue microarchitecture, the scaffolds do recapitulate the bone microenvironment in important ways. Previous work shows that these scaffolds induce osteogenesis of mesenchymal stem cells 27 and reproduce key features of breast cancer cell behavior in the bone tumor microenvironment.48,50
3D cell culture
A bone-homing and GFP-expressing variant of the parental MD Anderson-Metastatic Breast-231 (MDA-MB-231) (generously provided by Dr. Julie Rhoades) cell line was cultured with complete Dulbecco's Modified Eagle Medium (DMEM, with 10% fetal bovine serum, or FBS, and a 1% penicillin–streptomycin solution). Cell passage numbers ranged between P20 and P35. Scaffolds were seeded drop-wise with 1.5 million bone-homing MDA-MB-231 cells (MDA-B) and allowed to incubate on an orbital shaker (0.04 g) for 48 h at 5% CO2 and 37°C prior to any treatments to aid adhesion and infiltration. During this time period, MDA-Bs typically distribute across all surfaces of the scaffold and infiltrate roughly 200 μm into the total scaffold depth.16,48,49
Mechanical loading via applied perfusion
MDA-B cells seeded in scaffolds were subjected to mechanical loading via a custom direct perfusion bioreactor (Fig. 1B). DMEM flowed through the bioreactor’s tubing system to deliver cyclic inlet velocities of 1 (Low) and 15 (High) mL/min at 1 Hz to scaffolds within the bioreactor chambers. Perfusion magnitudes were chosen based on our previous work in which we simulated the fluid mechanical environment that would be experienced in future experiments by “high,” “medium,” and “low” applied flow rates that our custom bioreactor system engendered during perfusion. 49 We selected the “low” and “high” flow rates for this article because they had the largest separation in median shear stresses. Also, these flow rates correspond to shear stresses of approximately 60 and 180 mPa, both of which encompass previous studies’ estimates of physiological shear stresses from fluid flow along the bone surface during loading.14,32 Finally, in early optimization studies, we confirmed that cell number, assessed via total DNA content, was insensitive to our range of flow rates (Supplementary Fig. S1), corroborating previous work with compressive loading of scaffolds. 16 This loading method was performed for 1 h every day for 3 days. Static scaffolds were placed similarly in bioreactor chambers but did not undergo perfusion. In between loading bouts, scaffolds were returned to the orbital shaker. At the end of the 3-day loading period, scaffolds were either harvested immediately for RNA extraction (n = 3, 4 technical replicates, i.e., scaffolds, per condition per experimental “n”) or allowed to incubate on an orbital shaker for 24 h in 1% FBS before this low-serum cell culture media was collected for cytokine array analysis (n = 5).
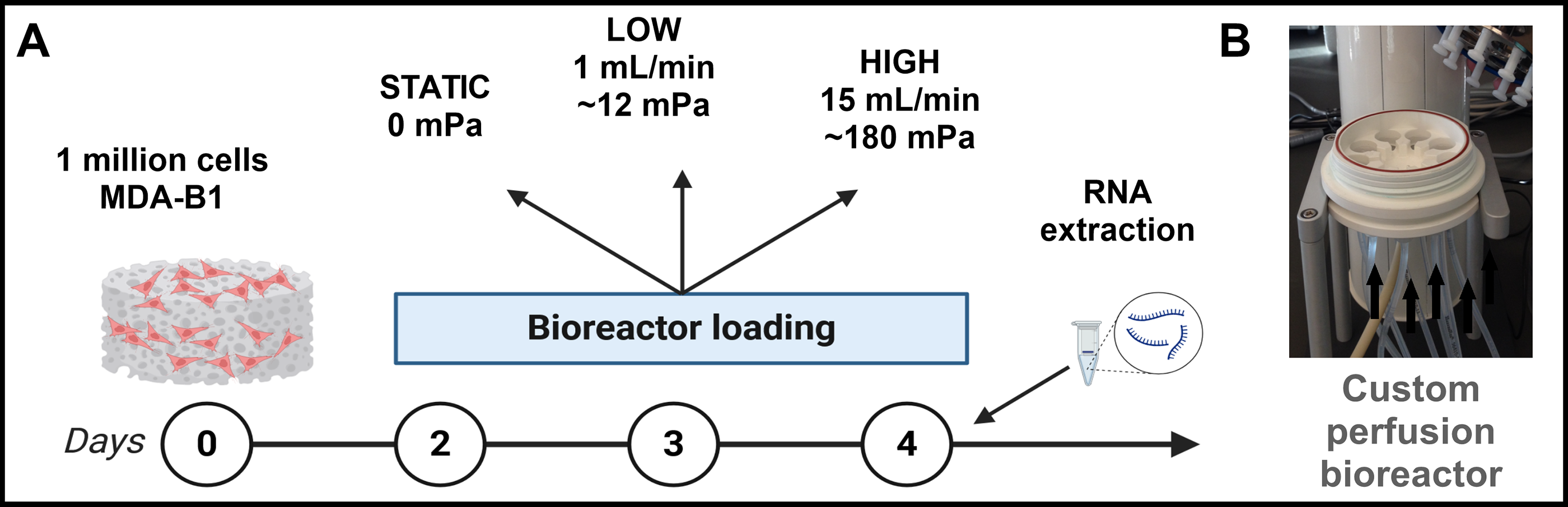
Perfusion via custom bioreactor.
Cell culture media was analyzed for a wide array of cytokines and chemokines (105 total) using a membrane-based immunoassay (Proteome Profiler Human XL Cytokine Array, R&D Systems, ARY022B). For sequencing, total RNA was extracted using an in-house protocol, from which poly(A)-selected RNA was converted into cDNA libraries with a size distribution of 250–350 bp using a KAPA Stranded mRNA-Seq kit (KAPA Biosystems, 07962193001). Dual-index adapters were attached at a concentration of 100 nM (KAPA Biosystems, 08861919702). Libraries were sequenced at 20 million reads per sample (paired-end) using an Illumina NovaSeq X instrument. Raw RNA-Seq data were processed via the BioFrontiers Fiji supercomputing cluster. Adapter sequences were trimmed, and low-quality bases (Phred score < 10) were filtered using BBDuk (BBMap v38.05). Read quality was evaluated after trimming using FastQC (v0.11.8). Reads were aligned to the hg38 reference genome assembly (UCSC Genome Browser) using STAR (v2.7.11a) with GENCODE v47 annotation. Gene-level counts were generated with featureCounts (v1.6.2), and differential expression analysis was performed in DESeq2 (v1.40.2). Genes with an adjusted p value < 0.05 and an absolute log2 of the fold change (FC) > 1 were considered significantly differentially expressed. We then used the open-source graphical gene enrichment tool ShinyGO 51 to identify significantly enriched biological processes and signaling pathways. Venn diagrams of differentially expressed genes were created using the open-source software “jvenn.” 52
Experimental design with irradiation and high-stimulus perfusion
Scaffolds seeded with MDA-B cells were divided into two groups. One group (Fig. 2, PRE) received irradiation at 0, 2, and 10 Gy 48 h after seeding and was allowed to incubate for 24 h prior to mechanical loading. After a 3-day loading period, scaffolds were then incubated for 24 h at 5% CO2 and 37°C, and RNA was extracted for quantitative real-time polymerase chain reaction (qPCR).
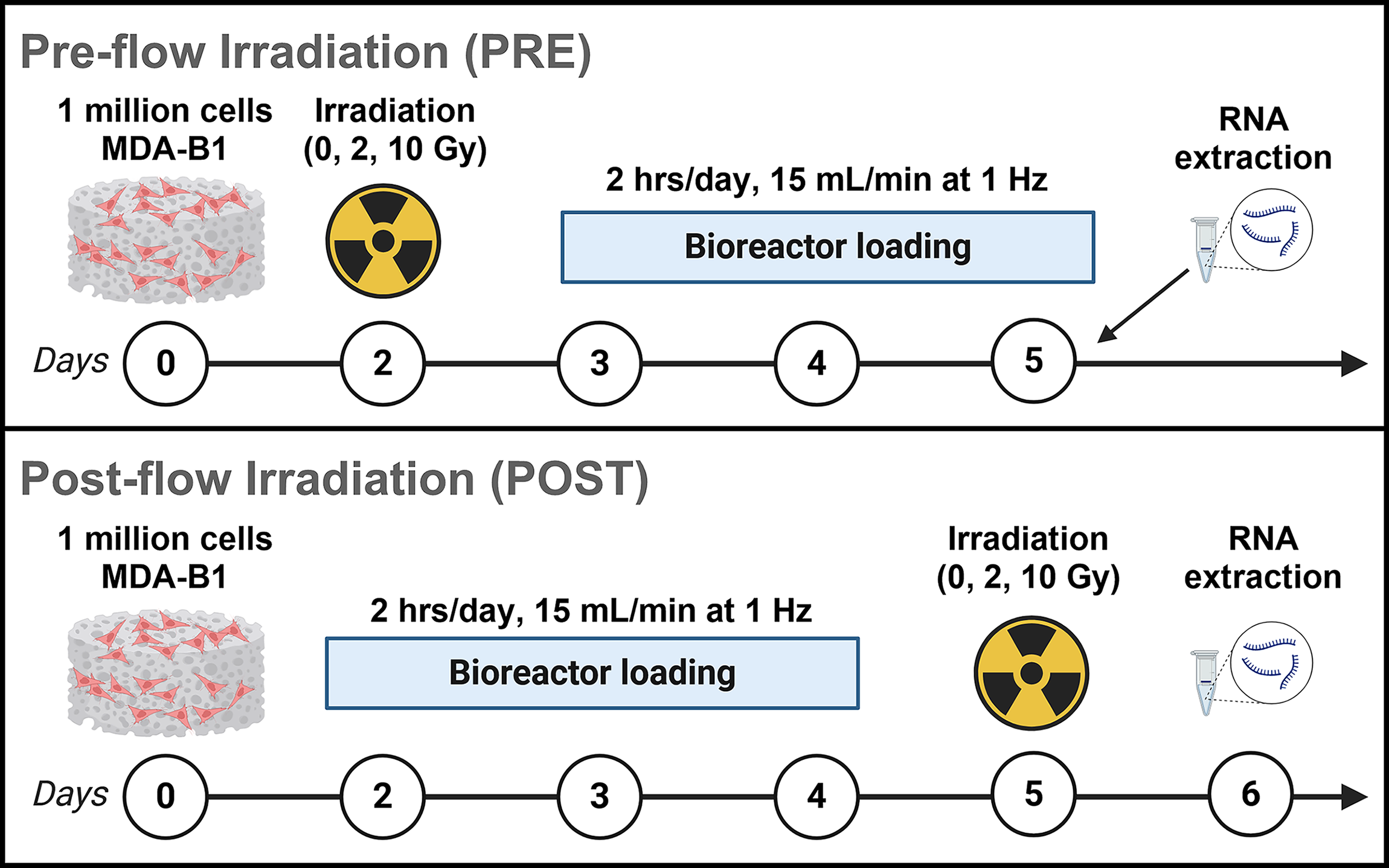
Time-sequence study with irradiation and high-stimulus perfusion.
The second group of scaffolds (Fig. 2, POST) was immediately subjected to loading after seeding. Once the 3-day loading period was complete, scaffolds were incubated on an orbital shaker for 24 h and subsequently received irradiation at 0, 2, and 10 Gy. These scaffolds were again incubated for 24 h at 5% CO2 and 37°C before RNA extraction for qPCR.
Samples were irradiated at room temperature in a Faxitron Cabinet X-ray System Model RX-650 (Lincolnshire, IL) at 115 kV and 5.33 rad/s. Loading bouts were administered at 2 h per day for 3 days, with 15 mL/min inlet flow rates for all loaded scaffolds. We applied a longer mechanical stimulation period to bolster a response to mechanical loading. Total RNA was extracted using an in-house protocol and converted into cDNA (Qiagen, 205311). cDNA was diluted to 25 ng/μL and 50 ng were quantified with qPCR. Real-time qPCR was run using a Rotor-Gene Q system (Qiagen, 9001862) and assays were assembled using primers designed in-house (Supplementary Table S1) and a QuantiNova SYBR Green qPCR kit (Qiagen, 208252).
Statistical analysis
For RNA-Seq, genes with an adjusted p value < 0.05 satisfying log2FC > 1 were considered upregulated, and those satisfying log2FC < −1 were considered downregulated. In our gene ontology enrichment analysis with ShinyGO, genes were sorted according to significance levels (p-adjusted < 0.05) set by Benjamini and Hochberg’s false discovery rate (FDR). 51 Treatment groups were normalized to nonloaded samples (reference group).
For time-sequence experiments, qPCR results were calculated using the efficiency-corrected 2-ΔΔCt method. We then conducted a three-way ANOVA with Tukey’s HSD post hoc testing (R 4.3.3 and GraphPad Prism 10.6.1) for main effect comparisons satisfying p ≤ 0.05. Treatment groups were normalized to nonloaded samples that did not undergo irradiation (reference group).
Results
High-magnitude fluid shear stresses amplified protumorigenic breast cancer cell gene expression
High-magnitude flow tripled the number of differentially expressed MDA-B genes in response to fluid shear (Fig. 3, 4). Notably, many of these genes are protumorigenic. Low-magnitude flow upregulated PDGFB (platelet derived growth factor subunit B), NEDD9 (neural precursor cell expressed, developmentally down-regulated 9), and LOXL4 (lysyl oxidase-like protein 4) (Table 1), all of which have been implicated in cancer cell migration and metastatic progression. PDGFB, in its PDGF-BB homodimer form, induces β-catenin expression by various breast cancer subtypes in vitro. 53 NEDD9 is a focal adhesion protein that plays significant roles in cell attachment, migration, and invasion,62,63 and LOXL4 is well known to be overexpressed in a number of cancers, interacting with cell-surface integrins to enhance cancer cell outgrowth. 56
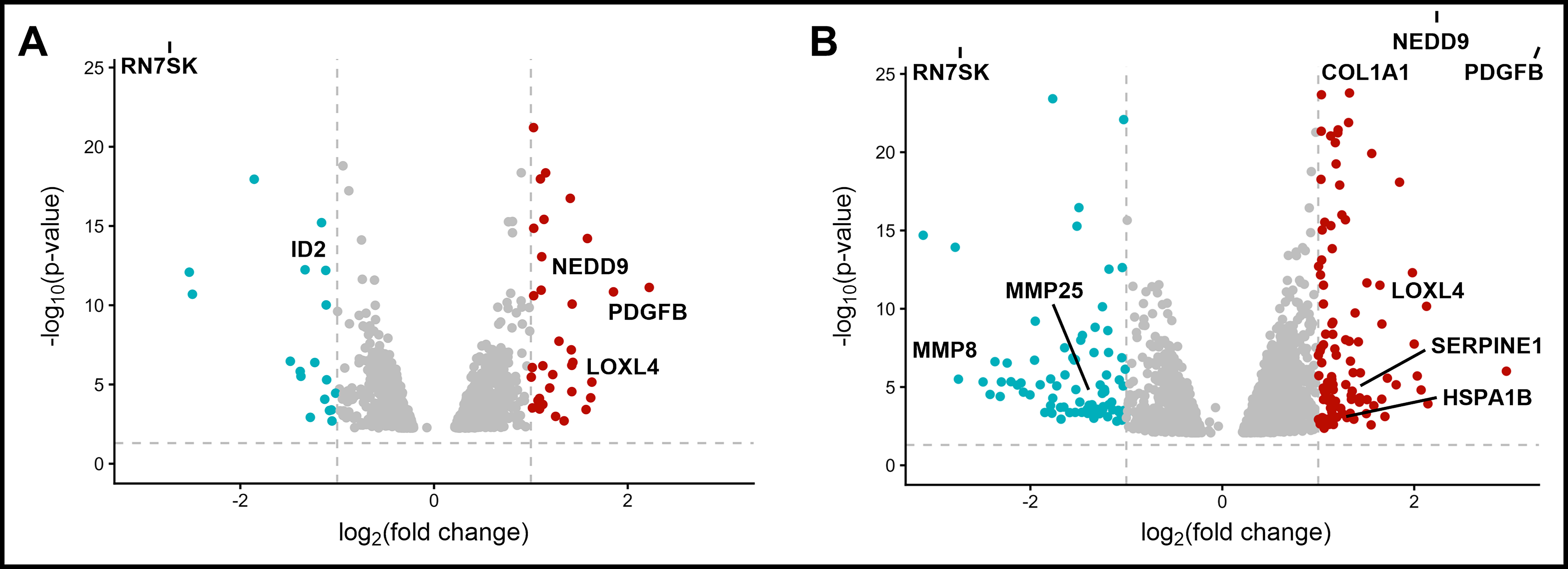
High flow results in more DEGs. Volcano plots of genes differentially expressed (threshold for up/down-regulation:
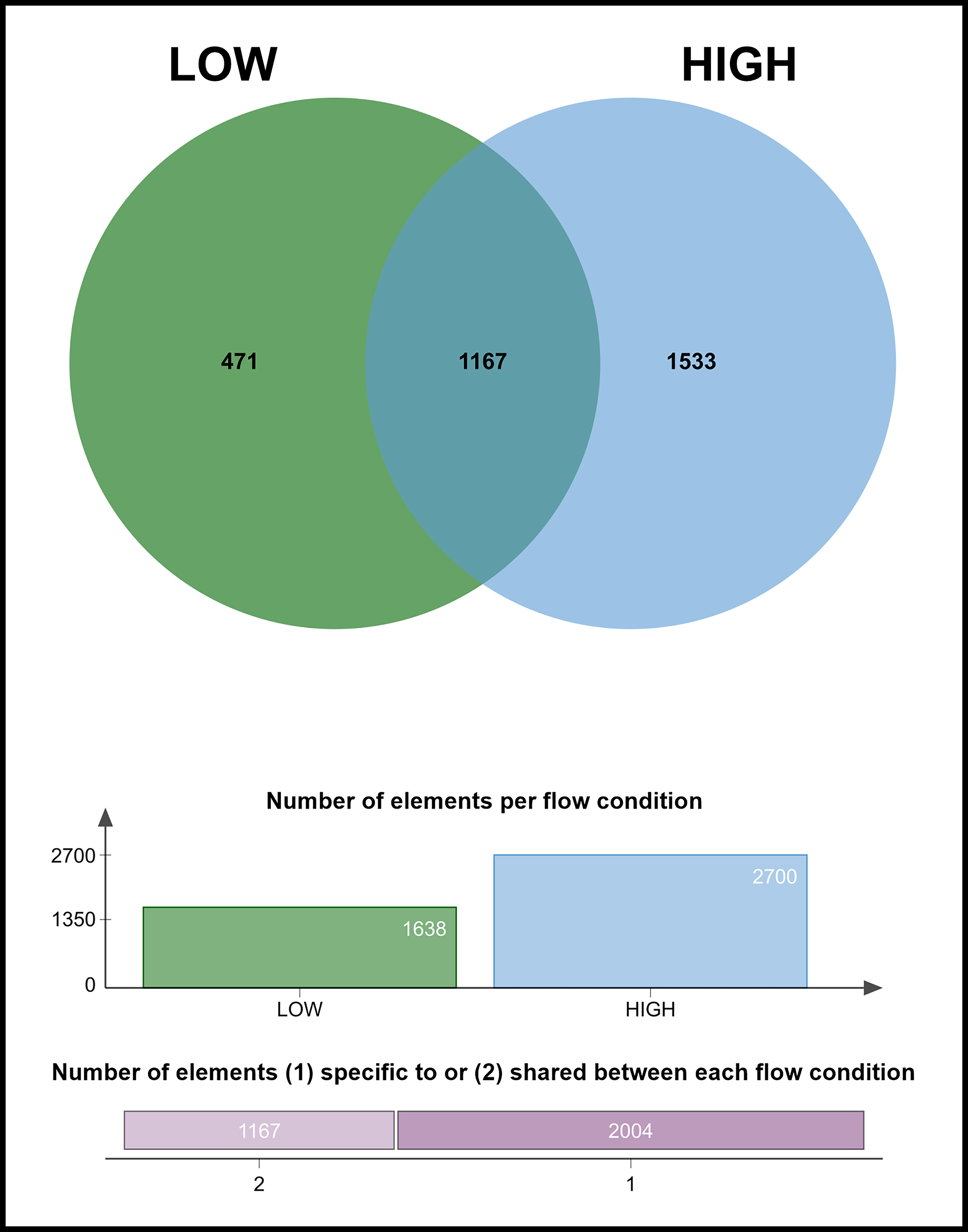
High flow produces a larger number of unique DEGs. While 1167 DEGs are shared between low and high flow conditions, high-magnitude flow expresses many more unique DEGs (1533) than low-magnitude flow (471).
Notable DEGs at the Low Flow Rate
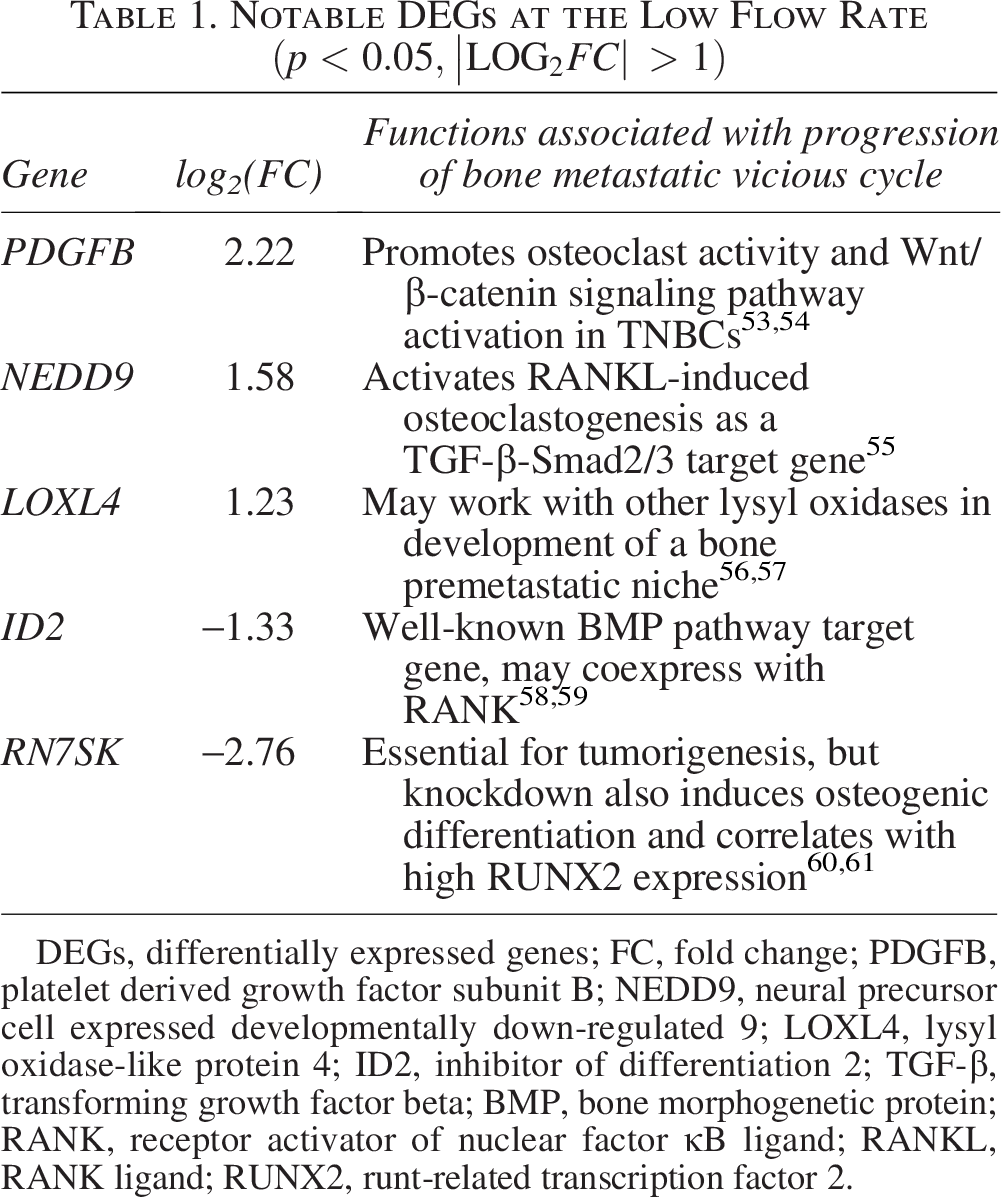
DEGs, differentially expressed genes; FC, fold change; PDGFB, platelet derived growth factor subunit B; NEDD9, neural precursor cell expressed developmentally down-regulated 9; LOXL4, lysyl oxidase-like protein 4; ID2, inhibitor of differentiation 2; TGF-β, transforming growth factor beta; BMP, bone morphogenetic protein; RANK, receptor activator of nuclear factor κB ligand; RANKL, RANK ligand; RUNX2, runt-related transcription factor 2.
Interestingly, high-magnitude fluid shear stress also downregulated several extracellular matrix (ECM) turnover markers, such as matrix metalloproteinases MMP25 and MMP8 (Table 2). This downregulation accompanied an upregulation of genes with important roles in ECM protein production, such as COL1A1 (pro-alpha1 chain of Type 1 collagen) and HSPA1B (heat shock protein family A member 1B). This suggests that higher-magnitude fluid flow spurs desmoplasia in the tumor microenvironment, as significant changes in MMP markers were absent in the low flow groups (Table 1).
Notable DEGs at the High Flow Rate

DEGs, differentially expressed genes; FC, fold change; PDGFB, platelet derived growth factor subunit B; TNBC, triple-negative breast cancer; NEDD9, neural precursor cell expressed developmentally down-regulated 9; RANK, receptor activator of nuclear factor κB ligand; RANKL, RANK ligand; TGF-β, transforming growth factor beta; LOXL4, lysyl oxidase-like protein 4; COL1A1, pro-alpha1 chain of Type 1 collagen; HSPA1B, heat shock protein family A member 1B; MMP, matrix metalloproteinase; RUNX2, runt-related transcription factor 2.
Gene ontology enrichment analysis revealed that both low- and high-magnitude flow enriched pathways associated with transcriptional misregulation in cancer (Fig. 5), designated by such genes as GADD45G (growth arrest and DNA-damage inducible gamma), ID2 (inhibitor of differentiation), and BAIAP3 (BAI-associated protein 3). These results also suggest that dynamic fluid shear may induce cancer stemness in MDA-B cells. In low-flow groups, MDA-B cells demonstrated enrichment of signaling pathways regulating stem cell pluripotency (Fig. 5A), while in high-flow groups, MDA-B differential gene expression was associated more specifically with the Hippo signaling pathway. However, there was a relatively high false discovery rate (log10FDR ∼1.8–2.8) associated with these stemness pathways (likely due to the low number of genes identified to be associated with these pathways), requiring more targeted investigation to confirm whether fluid shear does indeed stimulate cancer stemness. Many genes differentially expressed in the high-flow groups are also involved in pathways regulating cell motility, migration, and invasion.
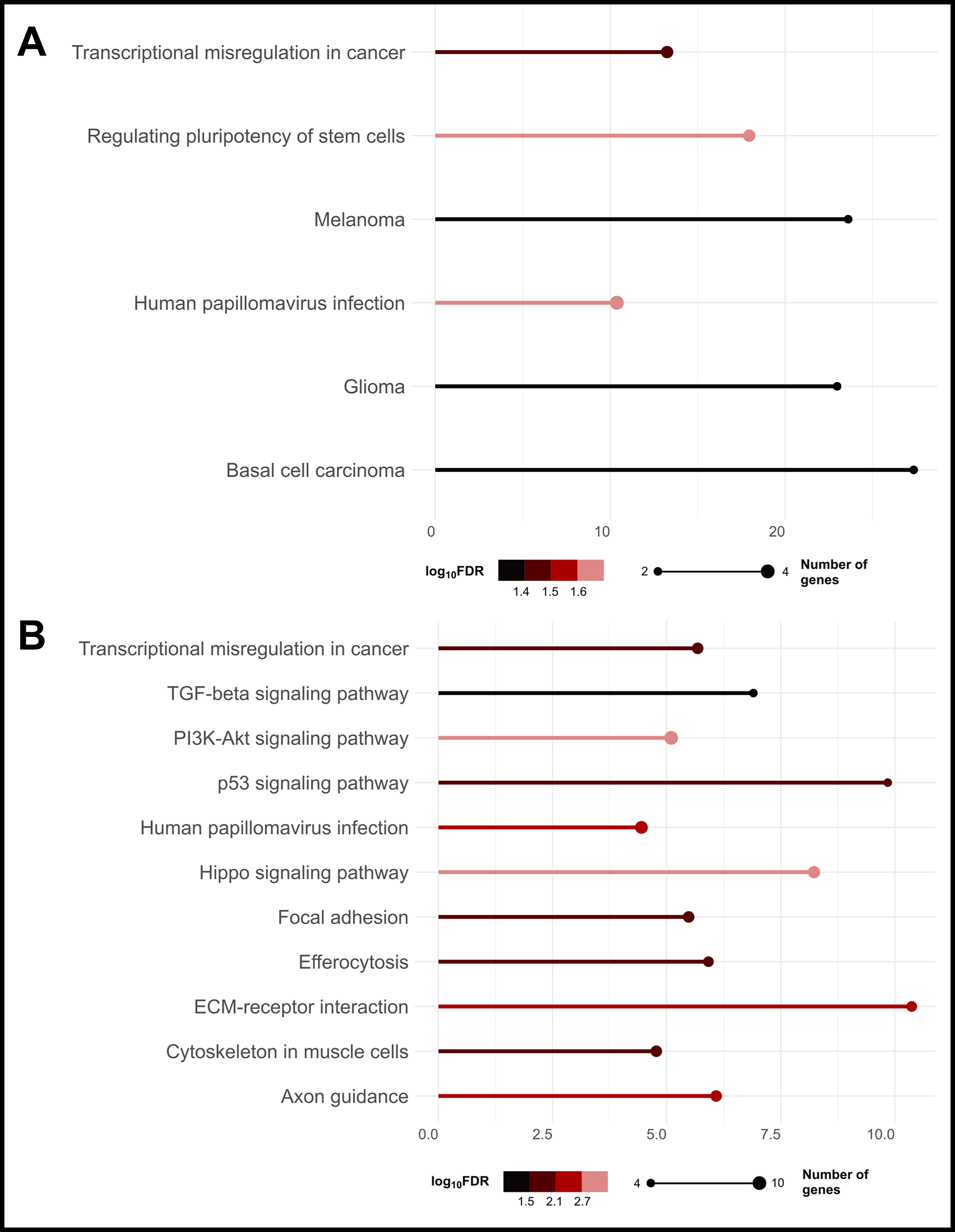
Higher magnitude fluid shear is increasingly protumorigenic.
One gene playing roles in many of these motility and invasion pathways, as well as cancer stemness-associated pathways, is SERPINE1. This gene encodes PAI-1, a serine protease inhibitor implicated in ECM remodeling, angiogenesis, and cancer progression.44,71 In our enrichment analysis, we found SERPINE1 to be involved in multiple enriched pathways, including Hippo, PI3K–Akt, TGF-β, and p53. While these pathways have quite far-reaching functions, they are known to be highly activated in metastatic cancers, especially TNBC.43,44,72
Additionally, many pathways such as TGF-β and PI3K-Akt have been implicated in their roles in mediating bone homeostasis in bone metastatic breast cancers.47,73 Elevated PAI-1 levels, particularly, are associated with poor prognosis in breast cancer, and emerging evidence points to a potential role in modulating osteocyte signaling and impairing bone quality.42,47,74 To confirm that flow altered PAI-1 secretion from MDA-B cells, we found that PAI-1 secretion followed SERPINE1 upregulation with high-magnitude fluid flow via an antibody array screening (Fig. 6B,C). Furthermore, SERPINE1 was one candidate gene that supported our interest in the combined effects of radiotherapy and mechanical loading on MDA-B expression. Ionizing radiation is known to induce SERPINE1 overexpression in TNBC cells in vivo, and this overexpression is generally protumorigenic, but whether this response is altered by mechanical loading is unknown. As such, we selected SERPINE1 as a target gene for subsequent experiments involving ionizing radiation.
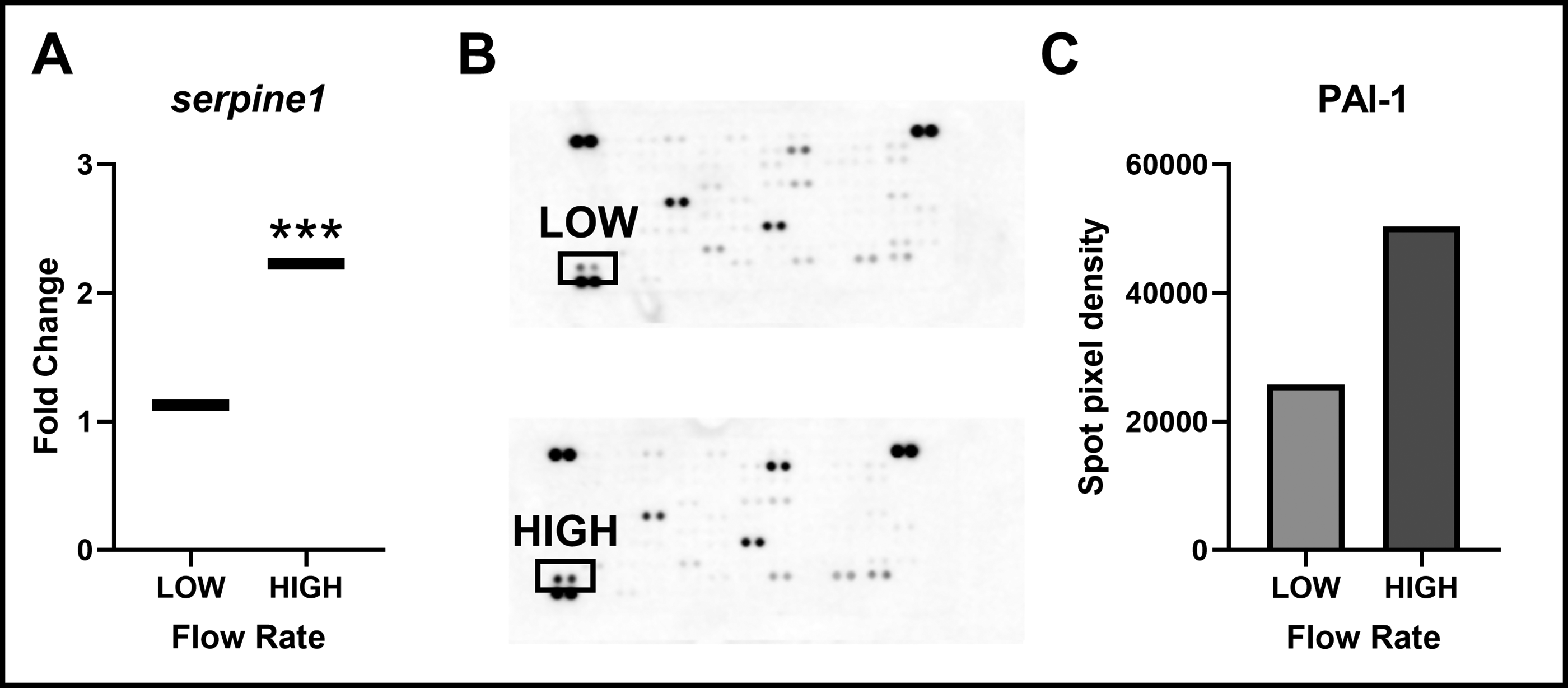
SERPINE1 is significantly upregulated with high-stimulus fluid shear stress.
Irradiation prior to flow enabled apoptosis evasion, but postflow irradiation may be more osteoprotective
Unexpectedly, we found that MDA-B gene expression of SERPINE1, Ki67 (MKI67, Marker of proliferation Ki-67), and CASP3 (caspase-3) was insensitive to irradiation dose irrespective of treatment sequence (Fig. 7, Supplementary Figures S2, S3, and S4). While previous studies show that parental MDA-MB-231 cells cultured in 2D begin undergoing apoptosis at 2 Gy of ionizing radiation, with marked apoptosis at 10 Gy, we found in our 3D model that 10 Gy did not significantly increase MDA-B CASP3 expression in nonloaded or loaded groups (Fig. 7B). There was a nonsignificant trend (p = 0.83) of decreased proliferation (downregulation of KI67) in loaded groups when loading preceded irradiation (Fig. 7C). Similarly, SERPINE1 was downregulated (ns, p = 0.89) as irradiation dose increased, but only when loading followed irradiation.
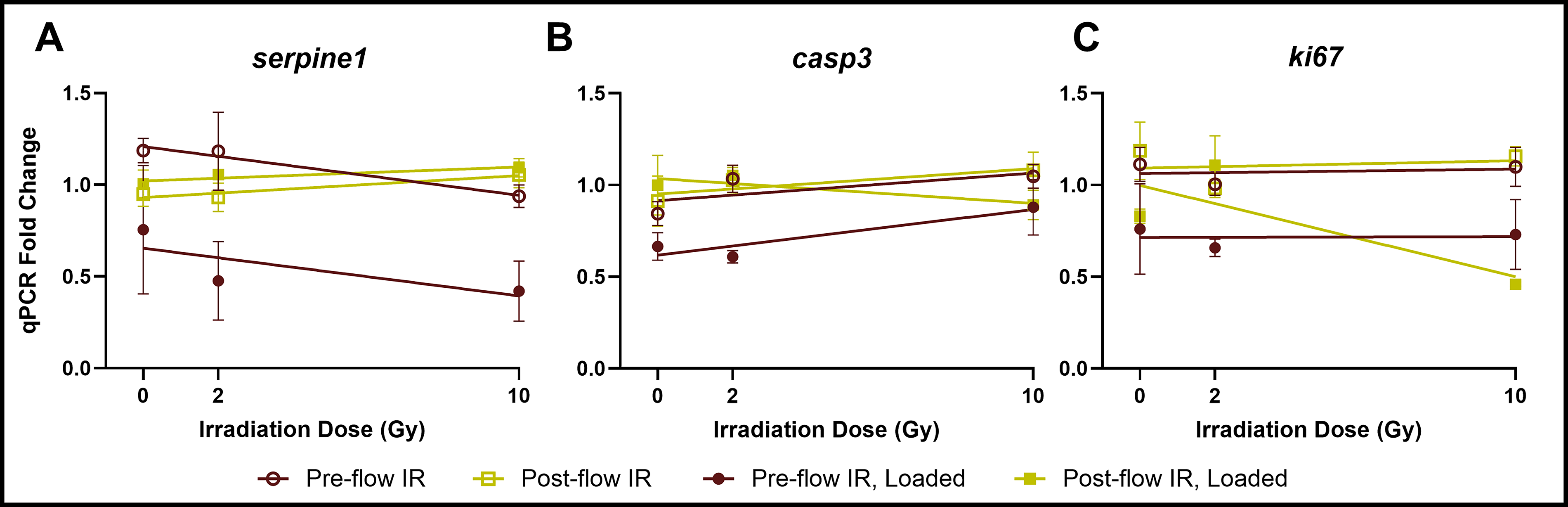
MDA-B gene expression of
Though MDA-B gene expression was insensitive to irradiation dose, with or without perfusion, we observed that the timing of irradiation administration significantly altered this expression. When we combined high-magnitude mechanical loading with irradiation, loading did not upregulate SERPINE1 expression, regardless of the sequence of experiments (Fig. 8A), in contrast to our dosed loading studies (Table 2, Fig. 6). In fact, when irradiation preceded loading (PRE), fluid shear downregulated SERPINE1 relative to nonloaded groups. When irradiation followed loading (POST), however, loading neutralized this effect. Regardless of the sequence of events, based on SERPINE1 expression alone, the inclusion of irradiation in these experiments made loading antitumorigenic and osteoprotective.
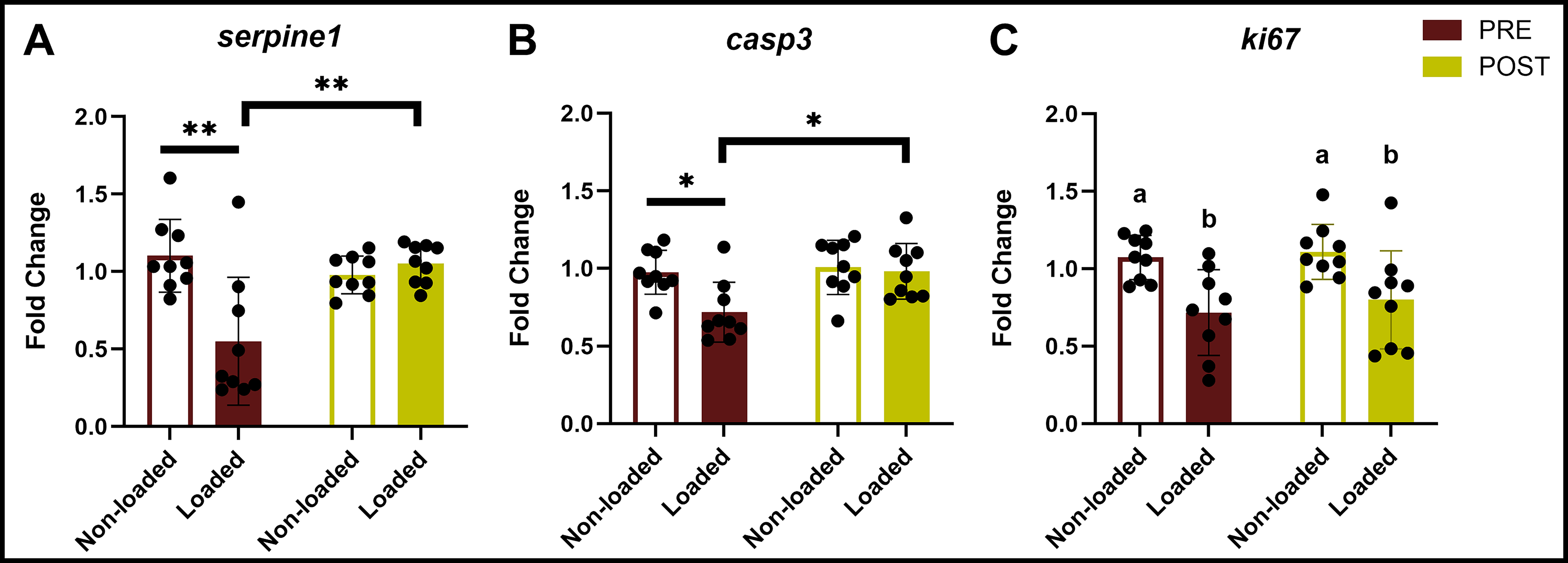
Postflow irradiation may be more osteoprotective.
Similar to SERPINE1, when irradiation preceded loading (PRE), MDA-B gene expression of CASP3 was downregulated (Fig. 8B), suggesting that preflow irradiation enabled cancer cell evasion of apoptosis. When irradiation followed loading (POST), CASP3 expression was unchanged with loading. Interestingly, KI67-mediated cell proliferation was consistently downregulated with loading compared to nonloaded groups, irrespective of irradiation timing (Fig. 8C).
Discussion
The objective of this study was to characterize the transcriptional mechanoresponse of breast cancer cells within a 3D bone-mimicking platform. 48 We showed that the protumorigenic response to fluid shear is sustained during bone metastasis using our 3D dynamic flow model (Fig. 5), as shown in previous studies with steady flow models. 41 Our data provides evidence that high-magnitude flow may stimulate fibroblastic gene expression in the tumor-bone microenvironment due to the concurrent downregulation of MMPs and upregulation of genes coding for ECM modeling functions, e.g., collagen production and chaperone protein-mediated regulation of ECM protein folding (Table 2). We identified one candidate gene highly expressed with high-magnitude flow: SERPINE1. We also showed that MDA-B cell secretion of PAI-1 (encoded by SERPINE1) increased with high flow as well.
While we have focused on one gene in this article, SERPINE1, our initial sequencing results demonstrate that dynamic, bone-mimicking fluid flow activated several key pathways in TNBC cells that require further investigation, especially in the context of the bone metastatic vicious cycle. An important observation from our sequencing data is that high-magnitude fluid flow promoted a persistent fibroblastic gene expression pattern in our MDA-B cells. Specifically, we showed that MDA-B cells upregulate ECM production genes while downregulating MMPs. Typically, elevated interstitial flows are known to promote ECM production in TNBC cell via a fibroblast-like phenotype, while increasing expression of ECM degradation markers such as MMPs.64,75,76 MMP upregulation is a key marker of protumorigenesis, contributing to increased invasion in TNBC cells. 77 However, our loading model for breast cancer bone metastasis downregulated MMPs at high magnitudes of flow, suggesting that breast cancer cells differentially respond to fluid shear once they have colonized the skeleton.
Our timing sequence study with irradiation and perfusion yielded surprising results. Several studies have shown that ionizing radiation (at both 2 and 10 Gy) upregulated both SERPINE1 and CASP3 in TNBC cells and induced significant amounts of apoptosis.45,78–80 Yet, we did not observe upregulation of SERPINE1 or CASP3 expression, even in our nonloaded groups (Fig. 8B). When irradiation was paired with flow, we in fact observed a suppression of SERPINE1 upregulation for both post- and preflow irradiation treatment. Moreover, previous studies on the DNA damage response of parental MDA-MB-231 cells to radiation (8–10 Gy) show that CASP3 is regularly upregulated, a response we did not observe in our nonloaded groups. We speculate that this shift could be due to the 3D culture environment, combined with the introduction of a loading stimulus that interferes with the typical TNBC cell response to radiation. We also speculate that the more unique results we observe in this study may be due to the use of an aggressive, bone-homing variant of MDA-MB-231 cells that is likely particularly resistant to ionizing radiation. In other bone-homing cell lines generated by the same technique (repeated in vivo passaging of MDA-MB-231 cells), bone-homing cells overexpressed genes such as SMAD2 (SMAD family member 2), HRAS (Hras proto-oncogene, GTPase), and TIP60, 81 all of which are associated with radioresistance in cancer cells.82–84 In our initial dosed loading experiments, we observed that flow enriched pathways associated with cancer stemness, consistent with literature on flow inducing cancer stemness. As a more stem-like profile is also associated with radioresistance, we anticipate that flow further magnified the radioresistance of this bone-homing variant. Further mechanistic studies are required to confirm this theory.
We measured CASP3 expression in this study as our marker of radiation-induced apoptosis and only observed detectable changes (downregulation) when irradiation preceded loading (PRE). However, irradiation also causes cell death via necrosis, indirect DNA damage, and inhibited cell division. 8 Thus, CASP3 expression alone may not fully capture MDA-B cell death due to radiation. It is possible that other mechanisms of cell death are instead leading to marked MDA-B cell death at 10 Gy, as we would expect. In our sequencing data, we observed no flow-based differential expression of markers of cell death via either apoptotic or necrotic processes, such as CASP3 and PPIF (peptidyl-prolyl isomerase F). 8 Accordingly, the significant downregulation of CASP3 with loading in preflow irradiation groups appears to be primarily a consequence of the timing of irradiation. In future studies, we aim to determine whether necrotic and autophagy-dependent cell death markers (also activated by ionizing radiation 8 ) respond similarly to CASP3. We also aim to quantify cell death via histology at various timepoints within these experiments to confirm whether evasion of cell death truly occurs in response to perfusion when the mechanical loading application follows irradiation.
Our interest in the MDA-B response to both radiation and bone-mimicking fluid shear stresses was due to the common use of radiation in bone metastasis treatment and its adverse effects on the skeleton. Clinical and in vivo studies show that high-dose therapeutic radiation results in decreased bone trabecular volume and increased bone marrow adiposity.85,86 Though some studies show positive effects on bone cells (mainly osteoblasts) with low doses of irradiation (∼1–2 Gy), low doses over the course of several weeks still lead to increased bone loss and adiposity in vivo.87,88 In the case of bone metastasis, ionizing radiation is known to alter cellular signaling pathways in both cancer cells and bone cells, with consequences that may intersect with or oppose mechanobiological regulation.85,86,89–91 As many pathways involved in the irradiation response are classically mechanical signaling pathways, we surmise that radiation sensitized MDA-B cells to fluid shear (PRE, Fig. 8) but suppressed the MDA-B response to fluid shear when administered postflow (POST, Fig. 8). Cell adhesion-mediated radioresistance, Wnt/β-catenin signaling, and regulation of nitric oxide expression and mechanosensitive ion channels are just some examples of classical mechanotransduction pathways that also participate in regulating the response to radiation.91–94 Other studies have shed light on the direct effects that mechanical forces have on the cellular response to radiation. For example, shear stress applied to endothelial and colon cancer cells through a parallel-plate flow chamber increased radiation-induced nitric oxide expression and increased radiosensitivity by modulating integrin/FAK signaling pathways. 94 Clearly, the ECM micromechanical environment has a significant role in mediating the response to radiation. Nonetheless, the mechanisms explaining how mechanical stress affects the cellular radiation response, and vice versa, are still unclear.
Here, we applied a single dose of radiation to mimic the fractionated doses patients would receive in a single session, allowing us to create a model for immediate effects arising from a single dose of radiation coupled with an applied load.7,9 When we applied ionizing radiation in this study, gene expression continued to be loading-dependent, and though MDA-B expression was irradiation dose-independent, the timing of irradiation administration significantly influenced our results. We found that preflow irradiation enabled cancer cell evasion of apoptosis and SERPINE1 downregulation, while postflow irradiation left markers of apoptosis unchanged alongside SERPINE1, reversing the MDA-B response to loading we observed in our sequencing data. Irradiation dose did not affect the breast cancer cell response to high-magnitude fluid shear, but timing did, and irradiation, regardless of sequence, may return an osteoprotective effect to loading in the bone-tumor microenvironment. Clinically, we suspect that, with multiple doses of radiation across several sessions, these effects could combine to result in sustained downregulation of protumorigenic genes like SERPINE1 by TNBC cells. Of the clinical studies that have examined the effects of radiotherapy combined with an exercise regimen, trials involving patients with spinal bone metastases found that this strategy generally improved bone density and did not further increase fracture risk.95–97 The results reported from our in vitro study, combined with future work further interrogating these mechanisms, will be useful in providing context to these clinical data.
While this study identifies key mechanosensitive targets in bone-homing breast cancer cells, there are limitations to the current approach. First, our 3D model is unicellular, which precludes the investigation of reciprocal, bidirectional signaling between cancer cells and the bone microenvironment. In vivo, the cancer cell mechanoresponse is undoubtedly modulated by other cell types. For instance, studies using osteocyte-conditioned media have yielded varied results, reporting both reduced metastatic potential and increased expression of protumorigenic factors.98,99 Notably, Verbruggen et al. demonstrated that these outcomes can differ significantly depending on whether cancer cells receive conditioned media or are in direct coculture, highlighting the critical importance of reciprocal signaling. 100 Furthermore, advanced multicellular platforms, such as the novel microfluidic chip developed by Mei et al. that integrates breast cancer cells, osteocytes, and an endothelialized lumen, are beginning to reveal how such complex environments regulate processes like intravasation. Second, our current scope was limited to characterizing the cancer cell’s transcriptional and secretory response. 101 Thus, we did not investigate the downstream functional effects of these signals (e.g., PAI-1) on osteocyte activity or bone remodeling. Future studies utilizing transwell coculture or integrated multicellular scaffolds will be necessary to determine how these load-induced signals ultimately impact the “vicious cycle” of metastasis. Nevertheless, this work provides a foundational transcriptome-wide map of how radiotherapy and mechanical loading intersect to regulate the breast cancer cell phenotype.
Conclusions
This study provides additional evidence of the direct effects of mechanical loading on breast cancer cell radiosensitivity and demonstrates that the timing of radiation administration is crucial to modulating the response to those mechanical signals. A key implication of our study is that integrating radiotherapy with loading may restore the osteoprotective effects of loading, which are lost in bone metastasis by suppressing flow-induced upregulation of a protumorigenic and/or proresorptive profile. More involved mechanistic studies are necessary to improve our understanding of the cell machinery that is directly involved in these responses. Nevertheless, we present this work as an assertion that physiologically relevant models applying fluid shear stress on the cells involved in the bone metastatic vicious cycle are necessary to investigate how common therapies interact with the tumor micromechanical environment to enable or hinder metastatic progression.
Authors’ Contributions
S.V.: Conceptualization, data curation, formal analysis, investigation, validation, visualization, writing—original draft, writing—review and editing. A.O.: Data curation, formal analysis, writing—review and editing. W.R.T.: Conceptualization, writing—review and editing. E.B.C.: Conceptualization, project administration, resources, supervision, writing—review and editing. M.E.L.: Conceptualization, funding acquisition, project administration, resources, supervision, writing—original draft, writing—review and editing.
Footnotes
Acknowledgments
The authors acknowledge the Cancer Center Support Grant P30CA046934. Sequencing was performed through the University of Colorado Anschutz’s Genomics Shared Resource (RRID: SCR_021984). ![]() on DNA content as a function of loading condition were gathered by Dr. Boyuan Liu. Bone-homing and GFP-expressing MDA-MB-231 breast cancer cells in this study were kindly provided by Dr. Julie Rhoades at Vanderbilt University, Nashville, TN, USA. The authors would also like to thank Dr. Tin Tin Su for allowing them to use her irradiator to complete the experiments in this study.
on DNA content as a function of loading condition were gathered by Dr. Boyuan Liu. Bone-homing and GFP-expressing MDA-MB-231 breast cancer cells in this study were kindly provided by Dr. Julie Rhoades at Vanderbilt University, Nashville, TN, USA. The authors would also like to thank Dr. Tin Tin Su for allowing them to use her irradiator to complete the experiments in this study.
Declaration of Generative AI in the Writing Process
During the revision of this article, the corresponding author used Gemini (Google) for language editing and to improve the clarity and flow of the Discussion section and responses to reviewer comments. All AI-generated suggestions were critically reviewed and verified by the authors. The AI was not used for data collection, analysis, or generation of original scientific conclusions, for which the human authors maintain full responsibility.
Data Availability Statement
Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This study was supported by NSF
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.