
Abstract
Brain metastases of triple-negative breast cancer (TNBC) rapidly progress, causing severe neurological decline with a median survival of less than 6 months. This tragic disease is often difficult to identify with sufficient time for treatment and is exacerbated by a lack of effective pharmacological intervention. Replicating the biochemical and mechanical properties of the premetastatic niche in vitro is a critical step in expediting the development of new therapeutics. However, a high-fidelity and reproducible model system is needed. To quantify the influence of a brain-mimetic microenvironment on brain metastatic TNBC, we encapsulated and cultured the TNBC cell line, MDA-MB-231 (P231), and its braintropic subline, MDA-MB-231-BrM2a-831 (BrM2a), in three premetastatic niches: a highly cell-adhesive and highly cell-degradable permissive niche, a highly adhesive but less degradable niche, and a nonadhesive but highly degradable niche. To mimic brain extracellular matrix, we functionalized the adhesive formulations with a brain-mimetic peptide cocktail and compared the cell responses to a “generic” RGDS-functionalization. This suite of conditions allowed us to investigate the influences of integrin-mediated adhesion, cell-mediated degradation, and cell type on the fate of P231s and BrM2as. Our data demonstrate that brain-mimetic adhesion has little to no impact on P231 phenotype, but the BrM2as display reduced viable cell density, reduced proliferation, and a higher proportion of both spherical clusters and spherical individual cells compared with the “generic” RGDS-functionalized niches. This suggests that brain-mimetic adhesion signaling encourages a rounded, less invasive phenotype in BrM2as. Modeling pathological processes usingadvanced, biomimetic in vitro models that better replicate in vivo cell phenotype have the potentialtoenhanceimprove the outcome of preclinical therapeutic testing.
Impact Statement
This study implemented a brain-mimetic polyethylene glycol hydrogel to replicate cell adhesion signaling provided by the extracellular matrix in the early stages of brain metastasis of triple-negative breast cancer. The data indicate that braintropic BrM2as display a stronger response to the brain-mimetic formulation compared with the parental 231 line that they were derived from, in comparison to RGDS-only functionalization. This in vitro cancer model addresses a critical gap in physiologically relevant model systems for metastasis research. The platform’s tunability and relevance to preclinical research establishes it as a viable tool for mechanistic cancer studies and potentially for accelerating the development of new therapies for brain metastasis.
Introduction
In the United States, ∼317,000 new cases of invasive breast cancer will be diagnosed in 2025. 1 Nearly 48,000 (15%) will be triple-negative breast cancer (TNBC). 2 TNBC is defined by the absence of estrogen receptor, progesterone receptor, and low expression of human epidermal growth factor 2, 2 which complicates treatment by eliminating these pathways as targets. About 25% of patients with metastatic TNBC will develop lesions in the brain. 3 TNBC brain metastases are associated with rapid progression, neurological decline, and a median survival of <6 months. 4 Understanding the mechanisms in TNBC brain metastasis progression is particularly challenging due to the complex brain microenvironment.
The brain is a highly organized, soft extracellular matrix (ECM) composed of many proteins. 5 Efforts to characterize the adhesive and degradable components have identified the primary constituents.6,7 A recent publication implemented this data to generate a brain-mimetic hydrogel and demonstrated the ability to culture primary astrocytes in vitro in a quiescent state. 8 They also demonstrated that incubation of brain metastatic cancer cells in a solution containing brain ECM peptides resulted in reduced spreading when reseeded on a surface. 8 Brain ECM composition, combined with low stiffness, creates a unique biochemical and biomechanical premetastatic niche. The brain is protected by the blood–brain barrier, which limits extravasation of circulating tumor cells and impedes delivery of most chemotherapeutics, further complicating in vivo studies. 9 Upon successful entry, disseminated TNBC cells must adapt to the brain microenvironment, where they may die, become dormant, or form colonies that progress into overt metastases. 10
Despite the clinical burden of TNBC brain metastasis, a gap remains in experimental models that recapitulate the brain premetastatic niche. In vivo models of TNBC injection into mice have contributed to understanding the metastatic cascade and drug efficacy but are limited by host variability, low throughput, and poor resolution. 11 Two-dimensional (2D) in vitro models are higher throughput and easy to implement, but incapable of replicating three-dimensional (3D) tissue architecture, ECM complexity, and cell–cell interactions. 12 Most 3D in vitro models present a generic ECM, which fails to reproduce brain-specific features and therefore lack the ability to recapitulate in vivo-like cellular phenotypes. 13 Recent models demonstrate the role that laminin and collagen play in brain metastases dormancy.14–16 To address limitations of existing models and simulate brain tissue, bioengineered in vitro models that mimic the brain microenvironment are needed to expedite the study of TNBC brain metastasis. Poly(ethylene glycol) (PEG)-based hydrogels are a suitable material due to their mechanical tunability and control over incorporated ECM components17,18 and degradability. 19 Previous work has shown the capacity of PEG hydrogels to replicate in vivo chemosensitivity, metastatic growth, dormancy, and reactivation.19–24
In an effort to establish a reproducible and biomimetic in vitro model, this study seeks to characterize the influence of brain-mimetic, cell-adhesion signaling and matrix degradability on the progression of braintropic TNBC. We implemented three hydrogel formulations with two peptide functionalization schemes: a highly cell-adhesive and highly cell-degradable formulation (permissive), a highly adhesive but less degradable formulation (degradation-restricted), and a nonadhesive but highly degradable formulation (adhesion-restricted). Degradation-restricted and adhesion-restricted formulations were previously shown to induce dormancy in TNBC.23,24 The permissive and degradation-restricted formulations were functionalized with either a brain-mimetic ECM composed of an 8-peptide cocktail 8 or a “generic” ECM composed of only RGDS. Encapsulated cells were able to degrade each formulation due to the inclusion of the matrix metalloproteinase (MMP)-degradable peptide sequence, GGGPQGWGQGK (PQ). In the degradation-restricted formulation, the inclusion of the non-enzymatically degradable comonomer, N-vinylpyrrolidinone (NVP), which increases cross-link density, decreased hydrogel degradability. To characterize the cellular responses to these formulations, we cultured the highly metastatic, parental TNBC cell line, MDA-MB-231 (P231), and its braintropic subline, MDA-MB-231-BrM2a-831 (BrM2a), and quantified viability, viable cell density, proliferation, apoptosis, and morphology after 1 and 15 days in culture. The brain-mimetic functionalization scheme had little impact on the behavior of P231s, but BrM2as displayed lower viable cell density, lower proliferation, a recovering apoptotic population, and a higher proportion of both spherical clusters and spherical individual cells in comparison to the RGDS-only functionalization. This influence on BrM2a phenotype that was not observed in the P231s supports the implementation of this model as a brain-mimetic, in vitro cancer model.
Materials and Methods
See Supplementary Data.
Results
Hydrogel characterization
Both mechanical and biochemical hydrogel properties influence cancer cell behavior.19–29 Accordingly, we quantified the swelling (Fig. 1C and D), degradation (Fig. 1E), and compressive modulus (Fig. 1F) of the five hydrogel conditions used in these studies.

Hydrogel characterization.
Swelling
Both permissive formulations displayed high volumetric swelling (arginine, gylcine, apspartic acid, serine (RGDS): 263 ± 16%; Brain-Mimetic 239 ± 11%) (Fig. 1C and D). Significantly lower swelling was observed for both degradation-restricted formulations (RGDS: 19 ± 3%, p = 2.31 × 10−10; Brain-Mimetic: 30 ± 3%, p = 1.64 × 10−9) (Fig. 1C and D) compared with their respective permissive formulations. This is due to the presence of additional crosslinks from NVP.19,20,22,30,31 The adhesion-restricted formulation displayed modest swelling (71 ± 10%) and proved statistically significantly different from all other conditions (p < 0.004). This decreased swelling compared with the permissive formulations is likely due to increased PEG-PQ crosslinking from the absence of PEGylated peptides, which can compete with crosslinks in the hydrogel backbone. 31 Prior quantification of PEG-peptide incorporation was performed on swollen hydrogels, allowing the influence of swelling on ligand density to be accounted for.20,22
Degradation
Cell-mediated hydrogel degradation is important for cell spreading, migration, and cluster formation. Cells degrade these hydrogels via MMP-2 and -9 mediated cleavage of the PQ sequence. We quantified the degradation of the five hydrogel conditions via collagenase IV exposure (Fig. 1E). During degradation, the fluorescently labeled macromer, methacryloxyethyl thiocarbamoyl rhodamine B, which was covalently crosslinked into the hydrogels, was released into solution. Complete hydrogel degradation can be represented by a stable fluorescence intensity over time once the label has been fully released (Fig. 1E). The permissive and adhesion-restricted formulations degraded at a similar rate and were completely degraded within 75 min (Fig. 1E). Both degradation-restricted formulations displayed the slowest degradation and did not fully degrade in the 200-min duration (Fig. 1E) due to the presence of nondegradable, NVP-mediated crosslinks. Since fluorescence intensities were used for quantification, a control condition, not exposed to collagenase, was used to correct for photobleaching.
Compressive modulus
The compressive moduli of both acellular and cell-laden hydrogels were quantified after 1 and 15 days in culture for each condition. The acellular, permissive formulations had the lowest compressive moduli with an average of ∼2.6 kPa (RGDS: 2.85 ± 1.25 kPa, Brain-Mimetic: 2.38 ± 1.35 kPa) (Fig. 1F). The acellular adhesion-restricted formulation had a slightly higher modulus (4.83 ± 1.53 kPa) (Fig. 1F). The two degradation-restricted formulations had the highest moduli of ∼11.3 kPa (RGDS: 12.01 ± 2.10 kPa, Brain-Mimetic: 10.65 ± 1.03 kPa) (Fig. 1F). Both acellular, degradation-restricted formulations had statistically significantly higher moduli than the other three formulations (p < 0.015). This is due to the presence of additional crosslinks from the NVP.30,31 Based on prior studies, the increased modulus for the degradation-restricted formulations was assumed to have a less significant impact on cell fate than reduced degradability. 22 All hydrogel conditions demonstrated a slight increase in modulus after cell encapsulation and 1 day of culture (Fig. 1F), but no statistically significant differences were observed. Similarly, after 15 days in culture, no statistically significant changes in the moduli of any formulation were observed (Fig. 1F). While there is a noticeable decrease in the modulus of the RGDS-functionalized, degradation-restricted formulation by day 15 (Fig. 1F), it proved to be nonstatistically significant for both the BrM2as (p = 0.0695) and P231s (p = 0.0668).
These data indicate a nonsignificant difference between the brain-mimetic and RGDS-only functionalization schemes, with the differences observed in swelling, compressive modulus, and degradation rate being driven by the hydrogel formulation alone (permissive, degradation-restricted, adhesion-restricted).
Cell phenotype characterization
Post encapsulation, cells were cultured for 1 or 15 days in the five hydrogel conditions. To characterize the phenotype of P231s and BrM2as in response to the hydrogel properties, their cell viability (Fig. 2), viable cell density (Fig. 2), proliferation (Fig. 3), apoptosis (Fig. 4), and morphology (existence as individual cells or cell clusters, sphericity, and volume) (Fig. 5) were quantified.
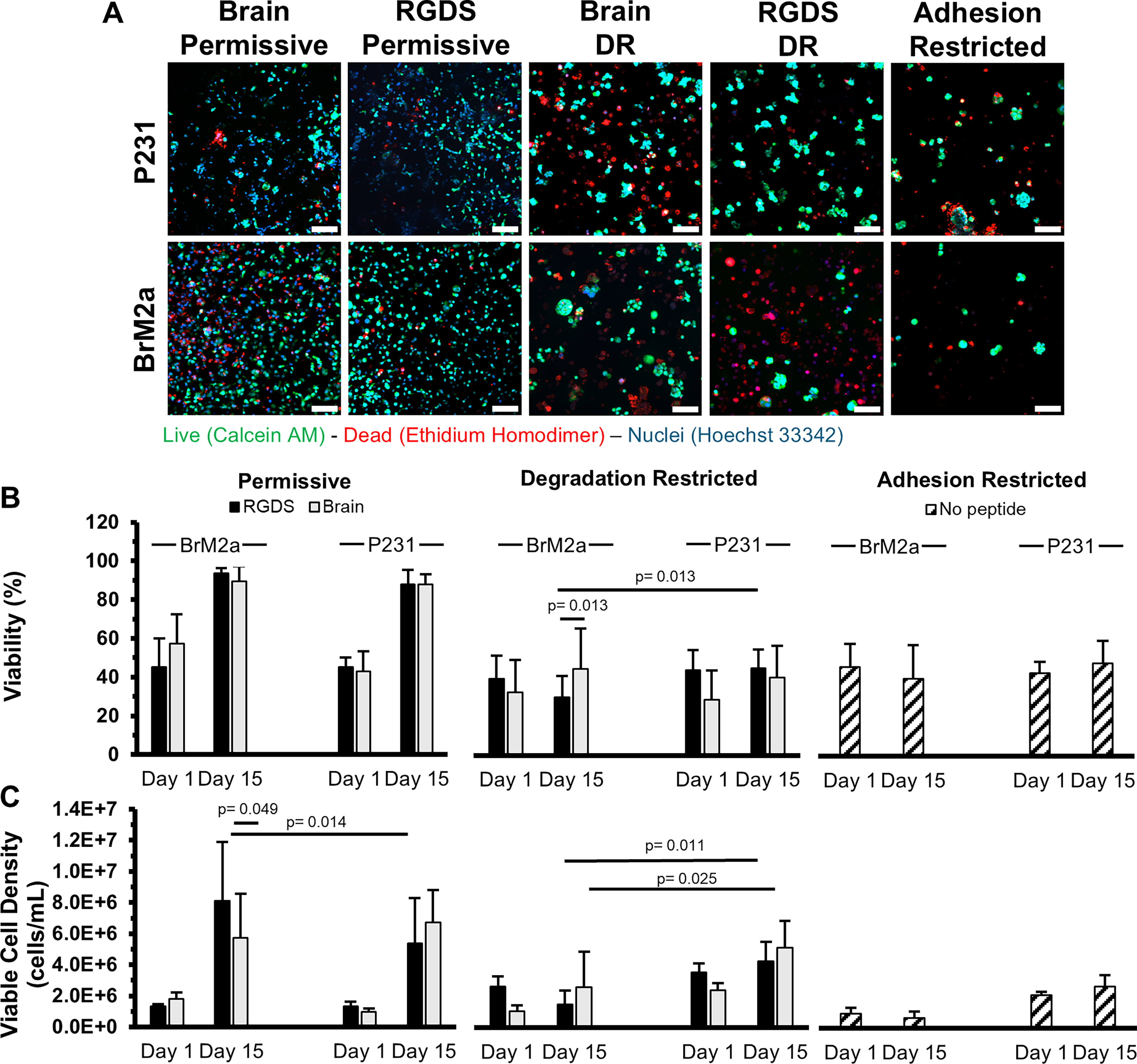
Viability and viable cell density.
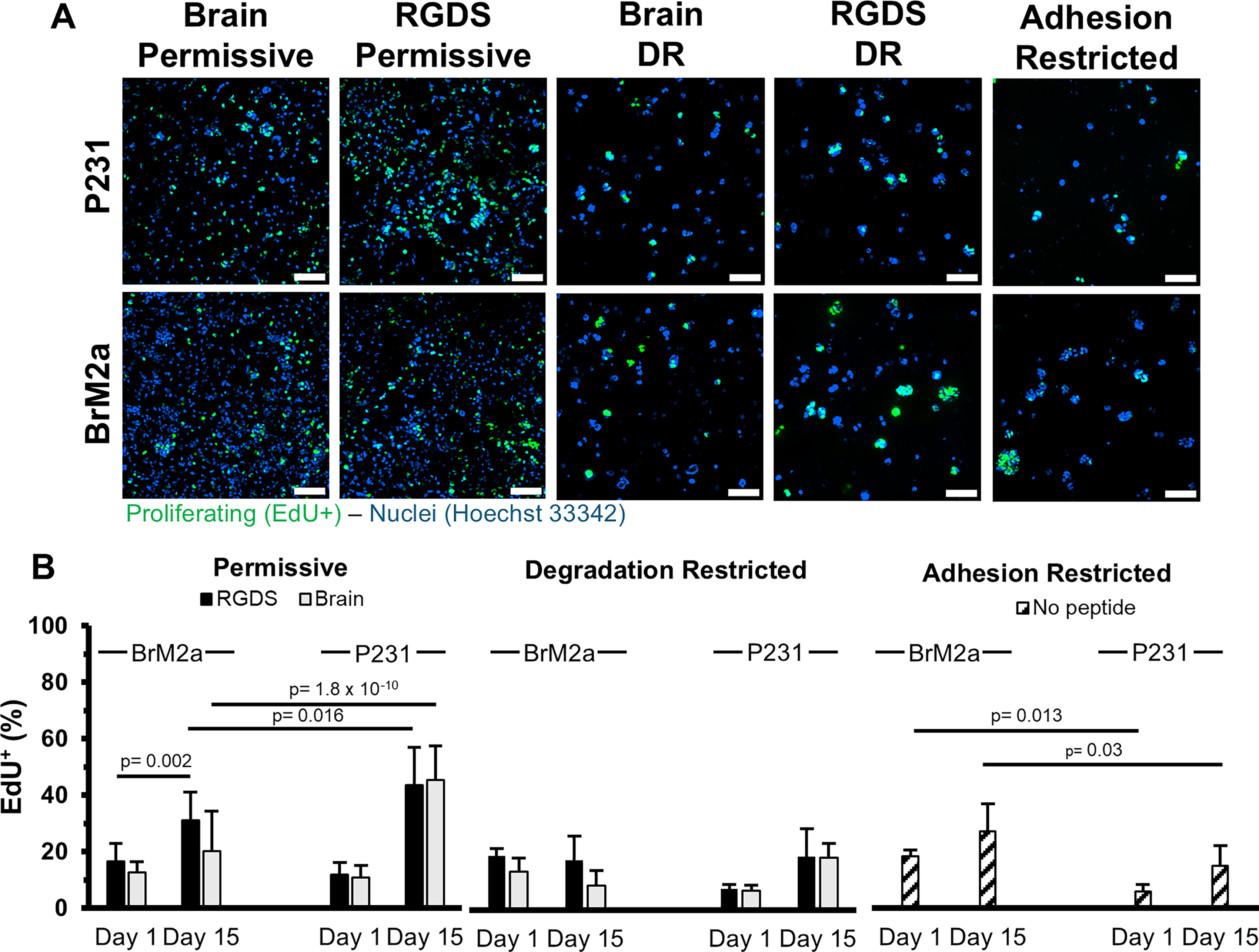
Proliferation.
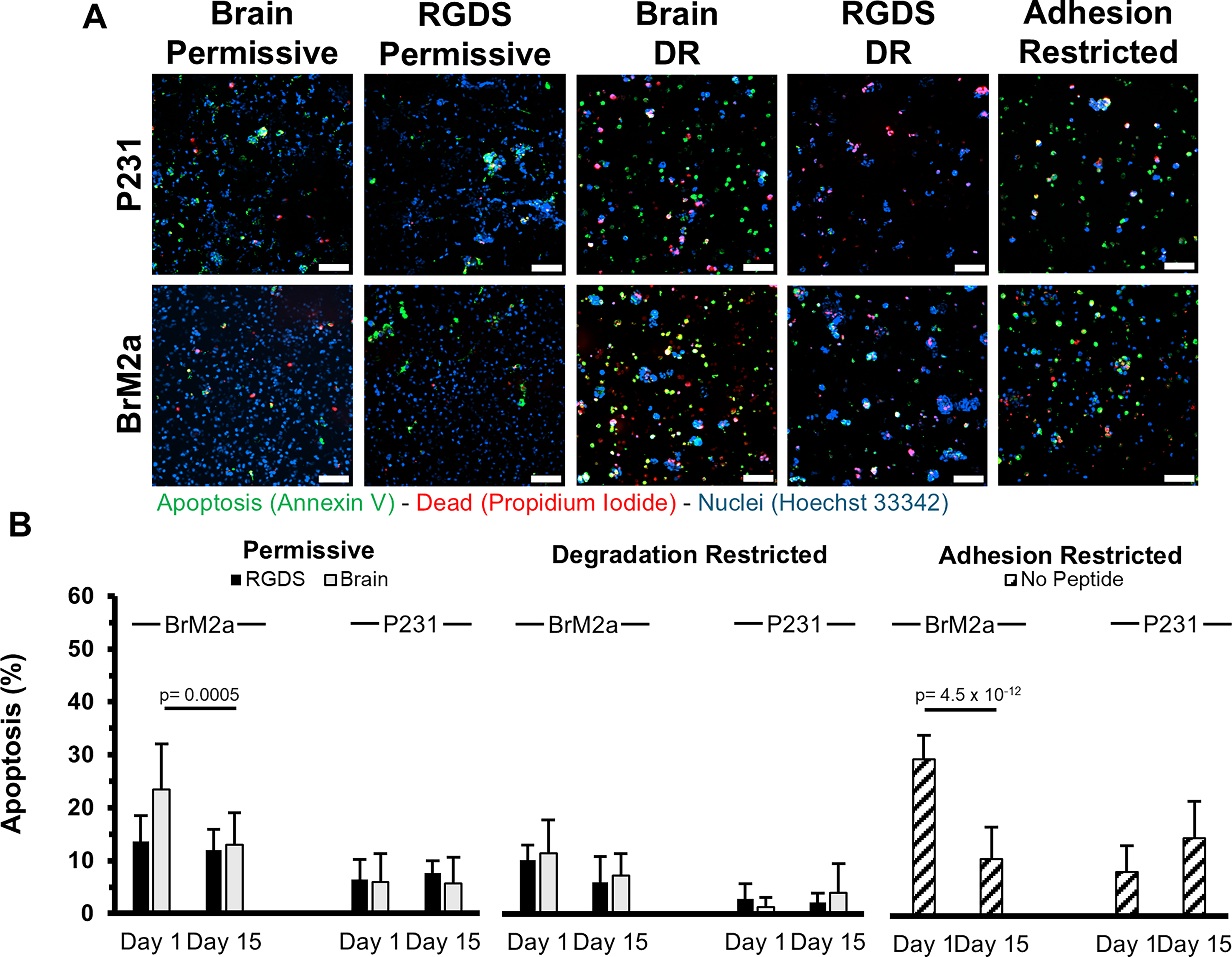
Early apoptosis.
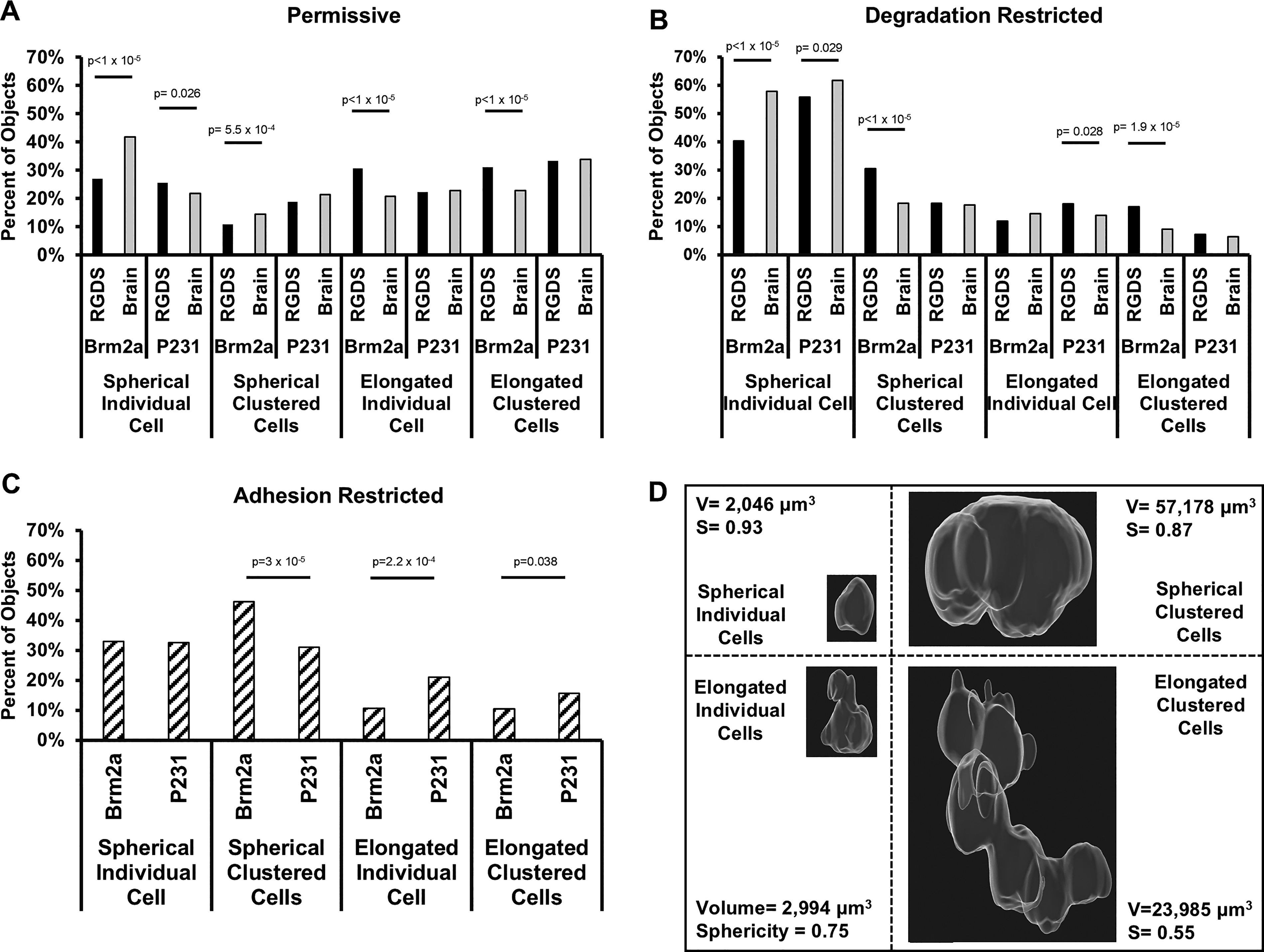
Morphology. Threshold gating of volume versus sphericity plots (Supplementary Fig. S1) was used to define four morphological states: (1) spherical individual cells, (2) spherical cell clusters, (3) elongated individual cells, and (4) elongated cell clusters. The percentage of the cell population residing in each morphology after 15 days in culture for the
Viability (P231s)
In both permissive formulations, the P231 viability significantly increased from ∼44%, 1 day post encapsulation, (RGDS: 45.2 ± 5%, Brain-Mimetic: 42.8 ± 10.4%) to ∼88% after 15 days in culture (RGDS: 87.9 ± 7.5%, Brain-Mimetic: 87.8 ± 5.4%) (Fig. 2B). P231 viability was significantly lower in all the degradation- and adhesion-restricted formulations compared with the permissive formulations. After 15 days in culture, P231s in the degradation-restricted formulations showed no statistically significant difference from day 1 with ∼36% viability (RGDS: 43.9 ± 10.3%, Brain-Mimetic: 28.6 ± 15%) to day 15 with ∼42% viability (RGDS: 44.9 ± 9.4%, Brain-Mimetic: 39.9 ± 16.3%) (Fig. 2B). The adhesion-restricted formulation also showed no statistically significant change from day 1 (43.0 ± 5.8%) to day 15 (48.2 ± 11.7%) (Fig. 2B).
Viability (BrM2as)
Similar to the P231s, the BrM2as cultured in the permissive formulations displayed a significant increase in viability from ∼51%, 1 day post encapsulation, (RGDS: 45.2 ± 14.8%, Brain-Mimetic: 57.2 ± 15.2%) to ∼92% after 15 days (RGDS: 93.7 ± 2.5%, Brain-Mimetic: 89.4 ± 7.8%) (Fig. 2B). BrM2as cultured for 15 days in the degradation-restricted formulations displayed a statistically significantly (p = 0.013) lower viability in the RGDS-functionalized hydrogels (29.8 ± 11%) compared with when cultured in the brain-mimetic hydrogels (44.5 ± 20.7%) (Fig. 2B), indicating a cell-specific response to the difference in biochemical functionalization. BrM2as cultured in the adhesion-restricted condition did not display a significant difference from day 1 (46 ± 12%) to day 15 (40 ± 17.5%) (Fig. 2B) but proved to be statistically significantly different from both permissive formulations (p < 2.63 × 10−13).
Viable cell density (P231s)
The P231 viable cell density significantly increased by ∼5.5-fold in both permissive formulations from ∼1.15 × 106 cells/mL (RGDS: 1.34 ± 0.29 × 106 cells/mL, Brain-Mimetic: 0.96 ± 0.24 × 106 cells/mL) to ∼6.05 × 106 cells/mL (RGDS: 5.39 ± 2.90 × 106 cells/mL, Brain-Mimetic: 6.71 ± 2.08 × 106 cells/mL) (Fig. 2C). P231s cultured in the degradation-restricted, RGDS-functionalized hydrogels displayed no significant change in viable cell density from day 1 (3.54 ± 0.59 × 106 cells/mL) to day 15 (4.27 ± 1.24 × 106 cells/mL) (Fig. 2C). However, P231s cultured in the degradation-restricted, brain-mimetic hydrogels displayed a statistically significant (p = 0.009) increase in viable cell density from 2.39 ± 0.46 × 106 cells/mL, 1 day post encapsulation, to 5.13 ± 1.73 × 106 cells/mL at day 15 (Fig. 2C). P231s cultured in the adhesion-restricted condition displayed no significant difference in viable cell density over 15 days in culture (Fig. 2C; day 1: 2.10 ± 0.24 × 106 cells/mL, day 15: 2.68 ± 0.73 × 106 cells/mL).
Viable cell density (BrM2as)
BrM2as cultured in both permissive formulations displayed a significant increase in viable cell density from ∼1.57 × 106 cells/mL, 1 day postencapsulation, (RGDS: 1.31 ± 0.17 × 106 cells/mL, Brain-Mimetic: 1.82 ± 0.41 × 106 cells/mL) to ∼6.92 × 106 cells/mL after 15 days in culture (RGDS: 8.10 ± 3.8 × 106 cells/mL, Brain-Mimetic: 5.74 ± 2.81 × 106 cells/mL) (Fig. 2C). The viable cell density of BrM2as cultured in the brain-mimetic, permissive hydrogel formulation was statistically significantly (p = 0.049) lower compared with the RGDS-functionalized, permissive hydrogel formulation at day 15 (Fig. 2C), again indicating a response to the functionalization scheme. The BrM2as displayed a statistically significantly (p = 0.014) higher viable cell density (8.10 ± 3.80 × 106 cells/mL), compared with the P231s (5.39 ± 2.90 × 106 cells/mL), after 15 days in the RGDS-functionalized, permissive hydrogels, indicating a cell-specific response. The viable cell density of BrM2as cultured in both degradation-restricted formulations did not significantly change (day 1: RGDS: 2.64 ± 0.66 × 106 cells/mL, Brain-Mimetic: 1.05 ± 0.40 × 106 cells/mL; day 15: RGDS: 1.51 ± 0.88 × 106 cells/mL, Brain-Mimetic: 2.62 ± 2.28 × 106 cells/mL) (Fig. 2C). The adhesion-restricted formulation induced lower viable cell density compared with the permissive formulations and did not significantly change (day 1: 0.95 ± 0.36 × 106 cells/mL; day 15: 0.65 ± 0.73 × 106 cells/mL) (Fig. 2C). These data illustrate that these unique formulations can impact cell fate with the permissive formulations promoting growth, the degradation-restricted formulations impeding growth, and the adhesion-restricted formulations severely impeding growth.
Proliferation (P231s)
In both permissive formulations, P231s displayed a statistically significant increase (RGDS: p = 7.77 × 10−16, Brain-Mimetic: p = 6.66 × 10−16) in the percentage of the cell population undergoing proliferation from ∼11.5%, 1 day post encapsulation (RGDS: 11.9 ± 4.3%, Brain-Mimetic: 11 ± 4.2%) to ∼44.6% after 15 days (RGDS: 43.7 ± 13.2%, Brain-Mimetic: 45.5 ± 11.9%) (Fig. 3B). The percentage of the cell population undergoing proliferation also statistically significantly increased (RGDS: p = 0.050, Brain-Mimetic: p = 0.036) in the degradation-restricted formulations from ∼7.2% (RGDS: 7.5 ± 1.3%, Brain-Mimetic: 6.8 ± 1.8%) to ∼18.5% (RGDS: 18.6 ± 9.9%, Brain-Mimetic: 18.3 ± 4.9%) (Fig. 3B). Contrary to the other hydrogel conditions, no significant difference in proliferation was observed in the adhesion-restricted formulation (day 1: 5.8 ± 2.6%; day 15: 15.1 ± 7.1%) (Fig. 3B).
Proliferation (BrM2as)
Unlike the P231s, the BrM2as displayed sensitivity to the functionalization scheme (RGDS vs. brain-mimetic). BrM2as cultured in the brain-mimetic, permissive formulation had 12.8 ± 3.7% of the cell population proliferating on day 1, which displayed a nonstatistically significant (p = 0.52) increase to 20.3 ± 14.1% on day 15. Contrastingly, BrM2as cultured in the RGDS, permissive formulation displayed a statistically significant (p = 0.0022) increase from 16.5 ± 3.7% to 31.1 ± 14.1%. Also, in both permissive formulations, the BrM2a proliferation was statistically significantly lower (RDGS: p = 0.016, Brain-Mimetic: p = 1.80 × 10−10) than the P231s. In both degradation-restricted formulations, proliferation decreased from ∼16.2% (RGDS: 18.9 ± 2.7%, Brain-Mimetic: 13.4 ± 4.7%) to ∼13.1% (RGDS: 17.5 ± 10.1%, Brain-Mimetic: 8.6 ± 5.3%) (Fig. 3B). In the adhesion-restricted formulation there was a nonsignificant increase in proliferation from day 1 (18.4 ± 2.2%) to day 15 (27.1 ± 9.7%) (Fig. 3B), but both values were significantly higher (day 1: p = 0.013; day 15: p = 0.030) than the P231s.
Apoptosis (P231s)
Annexin V labeling was used to identify cells undergoing early apoptosis. P231s cultured in the permissive formulations displayed a small percentage of the population in early apoptosis at ∼6.2% (RGDS: 6.4 ± 3.8%, Brain-Mimetic: 6 ± 5.3%) after 1 day in culture and ∼6.7% (RGDS: 7.6 ± 2.3%, Brain-Mimetic: 5.7 ± 5%) on day 15 (Fig. 4B). The apoptotic percentage of P231s cultured in the degradation-restricted formulations was also low with ∼2% (RGDS: 2.8 ± 2.8%, Brain-Mimetic: 1.2 ± 1.8%) on day 1 and ∼3.1% (RGDS: 2.2 ± 1.7%, Brain-Mimetic: 3.9 ± 5.5%) (Fig. 4B) on day 15. In the adhesion-restricted formulation, there was a nonsignificant increase from day 1 (∼8.3 ± 4.8%) to day 15 (14.5 ± 6.9%) (Fig. 4B). On day 15, the adhesion-restricted formulation was statistically significantly higher (p < 0.006) than all other gel types, excluding the RGDS-functionalized permissive formulation (p = 0.088).
Apoptosis (BrM2as)
The percentage of the cell population undergoing early apoptosis was consistent over the culture period in the RGDS-functionalized, permissive formulation with 13.6 ± 4.9% on day 1 and 12 ± 3.9% on day 15. There was a significant (p = 0.0005) decrease in apoptosis for BrM2as cultured in the brain-mimetic, permissive formulation from 23.5 ± 8.6% to 13 ± 6%. In the degradation-restricted formulations, there was a nonsignificant decrease in both functionalization schemes with ∼10.9% (RGDS: 10.2 ± 2.9%, Brain-Mimetic: 11.5 ± 6.3%) on day 1 and ∼6.7% (RGDS: 6 ± 4.8%, Brain-Mimetic: 7.3 ± 4.1%) on day 15 (Fig. 4B). A high level of early apoptosis was observed in BrM2as cultured in the adhesion-restricted formulation (29.3 ± 4.5%) which significantly decreased (p = 4.55 × 10−12) to 10.6 ± 5.9% (Fig. 4B). BrM2as showed a significantly higher percentage of early apoptotic cells compared with the P231s on day 1 in the adhesion-restricted formulation (p = 2.38 × 10−14) and in both brain-functionalized formulations (permissive: p = 9.44 × 10−11; degradation-restricted: p = 7.12 × 10−4). Also on day 1, there were nonsignificant differences between cell types in both RGDS-functionalized formulations (permissive p = 0.067, degradation-restricted p = 0.158) (Fig. 4B).
Morphology
Morphological analysis was implemented to quantify the percentage of the cell populations that persisted as individual cells or cell clusters along with sphericity and volume measurements of these individual cells or clusters after 15 days in culture (Fig. 5). The volume and sphericity of each object was quantified and plotted (Supplementary Fig. S1). Gates were placed vertically at a volume (V) of 4,000 μm 3 and horizontally at a sphericity (S) of 0.8. The volume gate was chosen because it was the mean measured volume of individual cells and provides a way to separate individual cells from cell clusters. The sphericity gate was chosen to differentiate round (>0.8) cells and cell clusters from more elongated (<0.8) cells and clusters. The four quadrants created by these gates provide a means to define four groups: (1: V ≤4,000 μm3, S ≥0.8) spherical single cells, (2: V >4,000 μm3, S ≥0.8) spherical clusters, (3: V ≤4,000 μm3, S < 0.8) elongated single cells, and (4: V >4,000 μm3, S < 0.8) elongated clusters. Differential integrin ligation/activation between the RGDS and brain-mimetic functionalizations may induce unique morphological phenotypes and will be evident in the proportion of objects found in each of these four classifications.
Morphology (P231s)
P231s cultured in both permissive formulations displayed a similar percentage of cells residing in spherical clusters (RGDS: 18.8%, Brain-Mimetic: 21.4%), elongated clusters (RGDS: 33.3%, Brain-Mimetic: 33.89%), or as elongated individual cells (RGDS: 22.3%, Brain-Mimetic: 22.9%). However, a significant increase (p = 0.026) in spherical single cells was observed in the RGDS-functionalized permissive formulation (25.6%) compared with the brain-mimetic permissive formulation (21.9%) (Fig. 5A). P231s cultured in both degradation-restricted formulations displayed a similar percentage of cells residing in spherical clusters (RGDS: 18.4%, Brain-Mimetic: 17.7%) and elongated clusters (RGDS: 7.3%, Brain-Mimetic: 6.6%). However, in the degradation-restricted conditions, the two functionalization schemes induced significant differences in spherical individual cells (RGDS: 56%, Brain-Mimetic: 61.8%, p = 0.029) and elongated individual cells (RGDS: 18.2%, Brain-Mimetic: 14%, p = 0.028) (Fig. 5B). The adhesion-restricted formulation induced the cell population to exist mostly as spherical individual cells or clusters (63.5%), which was higher than the permissive conditions (∼44%) (Fig. 5A), and lower than the degradation-restricted conditions (∼77%) (Fig. 5C).
Morphology (BrM2as)
In both permissive formulations, BrM2as displayed a significant, functionalization-induced difference in the percentage of cells residing as spherical clusters (RGDS: 10.9%, Brain-Mimetic: 14.5%, p = 0.00055), elongated clusters (RGDS: 31.2%, Brain-Mimetic: 22.8%, p < 0.00001), elongated individual cells (RGDS: 30.8%, Brain-Mimetic: 20.9%, p < 0.00001), or spherical individual cells (RGDS: 27.1%, Brain-Mimetic: 41.8%, p < 0.00001) (Fig. 5A). It is worth noting that the percentage of elongated clusters in the RGDS-functionalized, permissive formulation (31.2%) was similar to the P231s (RGDS: 33.3%, Brain-Mimetic: 33.8%), yet BrM2as cultured in the brain-mimetic, permissive formulation was much lower (22.8%). In both degradation-restricted formulations, there were also significant differences induced by the functionalization schemes in three of the four classifications: spherical clusters (RGDS: 30.5%, Brain-Mimetic: 18.3%, p < 0.00001), elongated clusters (RGDS: 17.1%, Brain-Mimetic: 9.1%, p = 0.00002), and spherical individual cells (RGDS: 40.4%, Brain-Mimetic: 57.9%, p < 0.00001) (Fig. 5B). The only nonsignificant difference occurred for elongated individual cells (RGDS: 12%, Brain-Mimetic: 14.7%, p = 0.14). In the adhesion-restricted condition, similar to the P231s, a majority of the BrM2a population existed as spherical cells or spherical clusters (79%), which was higher than both permissive conditions (∼47%) but only slightly higher than both degradation-restricted conditions (∼74%) (Fig. 5C). In the adhesion-restricted formulation, the BrM2as (46%) formed more spherical clusters than the P231s (31%). The P231s showed a larger portion of both elongated individual cells and elongated clusters than the BrM2as.
Discussion
Development and validation of tissue-mimetic matrices for in vitro culture is a critical step in the advancement of model systems. The differences in biochemical composition and mechanical properties of different organs further complicate the modeling of specific tissues. Models have been developed to mimic bone,32–35 liver,36,37 lymph,38,39 and brain.9,40,41 The highest quality reproduction of tissue metrics is currently achieved with ex vivo decellularized scaffolds, 42 but these scaffolds are heterogeneous and require complex processing. Bottom-up hydrogel design provides precise control of desired metrics while ensuring homogeneity and reproducibility between samples and experiments, but it requires in-depth validation. This bottom-up approach was utilized to develop a brain-mimetic hydrogel that was implemented to culture primary human astrocytes in a quiescent state, to demonstrate that astrocyte reactivity is sensitive to biochemical composition and that BrM2as display diminished spreading post incubation with the brain-mimetic peptide cocktail. 8 This inspired us to investigate if a similar brain-like hydrogel could serve as a brain-mimetic model to quantify the response of braintropic TNBC cells.
We utilized brain-inspired formulations to characterize the response of the parental P231, and its braintropic subline, BrM2a, to different premetastatic niches. To contrast the brain-mimetic functionalization, we utilized a “generic” RGDS-only functionalization with the same total peptide concentration. Utilizing both functionalizations, we implemented a permissive formulation to approximate the elasticity and degradability of native tissue, a degradation-restricted formulation with the same peptide concentration but significantly lower degradability to investigate the role of MMP-mediated degradation independent of ECM adhesion signaling, and an adhesion-restricted formulation as a control. The brain-mimetic and RGDS functionalization schemes did not induce differences in the hydrogel mechanical properties but differences were observed between formulations that were independent of peptide functionalization. While all hydrogel formulations displayed volumetric swelling following polymerization, the most substantial was in the permissive formulations, with modest swelling in the adhesion-restricted formulation, and relatively low swelling in the degradation-restricted formulation. The adhesion-restricted and permissive formulations showed similar degradation rates and compressive moduli, with the degradation-restricted formulations displaying a reduced degradation rate and significantly higher modulus. These differences in formulation and functionalization result in five microenvironments that allow systematic investigation into which hydrogel properties influence cell fate.
Evaluation of cell viability and viable cell density provides a snapshot of the cellular response to their microenvironment. These metrics revealed a cell type-dependent response to the degradation-restricted formulations, as the BrM2as displayed a significantly lower viable cell density in both brain-mimetic and RGDS functionalization schemes compared with the P231s. This and other observed cell type differences are likely a result of expression changes in the 179 genes identified to be influenced by the repeated injection and recovery used to isolate the brain-tropic BrM2a cell line. 43 The BrM2as also displayed a functionalization-dependent response with significantly higher viable cell density in the RGDS-functionalized, permissive formulation compared with the brain-mimetic, permissive formulation. The BrM2a viable cell density in the RGDS-functionalized, permissive formulation was also significantly higher than the P231s. Interestingly, BrM2a viability was significantly lower in the RGDS-functionalized, degradation-restricted formulation compared with the brain-mimetic functionalization, indicating that the brain-mimetic cocktail may aid BrM2a survival. BrM2a sensitivity to RGDS in the permissive formulation could be facilitated by upregulation of proteins such as leupaxin and RhoGDI2, which have roles in downstream signaling of β1 integrin. Leupaxin and RhoGDI2 were identified as upregulated >1.5-fold in the BrM2a line compared with the parental line. 43 It should be noted in previous publications,21,23,24 viability at the first time point was higher than reported here. This is likely due to previous studies quantifying viability 4 h post encapsulation rather than 24 h used here. The exposure to free radicals produced during photopolymerization can injure cells but may not kill them immediately, 44 and this impact may not have been captured previously.
Microenvironmental properties are also known to influence cell fate by regulating proliferation and apoptosis. EdU incorporation data indicate cell type-dependent proliferation differences in each of the three formulations, with lower BrM2a proliferation in both permissive functionalizations, stagnant BrM2a proliferation contrasted by increasing P231 proliferation in the degradation-restricted formulations, and higher BrM2a proliferation than P231 in the adhesion-restricted formulation. These cell type-dependent differences are also likely caused by differences in gene expression between P231s and the BrM2as. Only the BrM2as displayed a functionalization-based difference in the permissive formulation with increased proliferation in the RGDS-functionalization and a nonsignificant change in the brain-mimetic scheme. This functionalization-based difference is consistent with increased mitogenic signaling through elevated β1 integrin signaling. For both cell types, in nearly all conditions, there was a nonsignificant change in the proportion of apoptotic cells from day 1 to 15, but in the brain-mimetic, permissive, and adhesion-restricted formulations, the BrM2as, not the 231s, showed a significant decrease in the percentage of the population undergoing early apoptosis. For both conditions, the annexin V positive cell population was comparatively high on day 1 and may be a result of upregulated cell death mediators caspase1 and CRADD, but the BrM2a recovery may be a result of upregulated expression of DUSP1, serpins, and interleukins, which act to suppress automated cell death. 43
Microenvironmental cues can also influence morphology and invasivity. Elongated cells and clusters may represent an invasive, more mesenchymal phenotype, while spherical cells and clusters are often less invasive. The P231 morphology demonstrated a modest impact of the functionalization scheme on the distribution of individual cells, with more spherical individual cells in the RGDS, permissive formulation compared with the brain-mimetic, permissive formulation. Alternatively, P231s in the RGDS degradation-restricted formulation produced more elongated individual cells than the brain-mimetic, degradation-restricted formulation. The BrM2as displayed a much stronger reaction to the functionalization schemes, with the brain-mimetic functionalization inducing a larger percentage of spherical individual cells and lower percentage of elongated clusters in both the permissive and degradation-restricted formulations. This similarity in distribution in both the brain-mimetic permissive and degradation-restricted formulations suggests a strong influence of adhesion signaling in the determination of this tendency toward spherical single cells.
Taking all these data into consideration, it can be concluded that sensitivity to the functionalization scheme is largely restricted to the BrM2as with little to no influence on the behavior of the P231s. BrM2a sensitivity to the brain-mimetic functionalization is evident in the lower viable cell density, lower proliferation, recovering apoptotic population, and higher proportion of both spherical clusters and spherical individual cells in the brain-mimetic, permissive condition compared with the RGDS permissive condition. These results suggest that for BrM2as, brain-mimetic adhesion signaling promotes, but does not enforce, a more rounded, less aggressive phenotype, which may aid their persistence in brain tissue. The ability of the brain-mimetic, permissive formulation to elicit this BrM2a behavior supports the capability of this formulation as a tissue-mimetic matrix for further in vitro studies. Future implementation of this hydrogel formulation has the potential to contribute to the identification and optimization of critically needed therapeutics to overcome the unique challenges presented in treating brain metastatic TNBC.
Authors’ Contributions
S.F. performed the experiments, analyzed the data, performed statistical analysis, drafted the original article, and prepared the figures. J.S. provided mentorship, critical review, and editing of the article and figures and edited the final documents.
Footnotes
Funding Information
Research reported in this publication was supported by an Institutional Development Award (IDeA) from the National Institute of General Medical Sciences of the National Institutes of Health under award number P20GM113125, by the Institute for Engineering Driven Health at the University of Delaware (UD), and the State of Delaware through the Delaware Biotechnology Institute’s Bioscience Center for Advanced Technology. Microscopy access was supported by grants from NIH-NIGMS (P20GM139760), the State of Delaware, and an Institutional Development Award (IDeA) from the National Institute of Health’s National Institute of General Medical Sciences under grant number P20GM103446. S.F. was partially supported through a UD Graduate Scholar Award.
Disclosure Statement
No competing financial interests exist.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.