
Abstract
Introduction:
Nanoliposomes (NLs) are a rich source of phospholipids, including phosphatidylcholine, and can encapsulate and release hydrophilic and hydrophobic compounds, separately or simultaneously, to different cells.
Objectives:
This study aimed to assess the impact of NLs or nanomicelles (NMs) on characteristics of frozen–thawed bull sperm.
Methods:
Semen samples were collected with an artificial vagina twice weekly from five bulls. Ejaculates that met minimum characteristics (n = 36) were pooled, extended with 1, 2, or 3% NL or NM, and cryopreserved. Frozen–thawed sperm were assessed regarding motility (computer-assisted sperm analysis), morphology, and plasma membrane integrity (eosin-nigrosin), plasma membrane functionality (HOST), acrosome integrity (Pisum sativum agglutinin), and fertility (in vitro fertilization).
Results:
Extender with 2% NL had the highest (p < 0.001) total motility (69.85 ± 0.92%), progressive motility (58.90 ± 0.81%), sperm with integrated membrane (68.88 ± 0.49%), sperm with active membrane (65.02 ± 0.53%), sperm with intact acrosome (75.28 ± 1.1%), and in an in vitro fertilization system, the highest rates of cleavage (68.06 ± 3.98%) and blastocyst formation (40.10 ± 3.44%).
Discussion:
NL provided better cryoprotection than NM for bull sperm. Supplementing the extender with NL could enhance bull semen cryopreservation.
Introduction
Sperm cryopreservation is a critical tool for artificial insemination. 1 Sperm membrane integrity is affected when sperm are frozen. A consequence of sperm cryopreservation is damage to sperm membranes during cooling to 5°C. 2 These membranes contain high concentrations of unsaturated fatty acids and, therefore, are prone to oxidative stress, especially during freezing and thawing. 3 Furthermore, reducing the ratio of unsaturated fatty acids to saturated fatty acids increases damage to the sperm plasma membrane during cooling. 4 In addition, membrane fluidity decreases during sperm freezing, but increases during thawing, due to changes in membrane phospholipid composition. 5 Soy lecithin (SL) is used as an extracellular cryoprotectant to preserve phospholipids that protect sperm during cryopreservation. 6
The size of the SL particles in extenders is widely recognized as a critical determinant of the quality of frozen–thawed semen. 7 SL nanoparticles, characterized by their reduced particle size and enhanced solubilizing ability compared with untreated counterparts, facilitate improved interaction and coating of sperm. This enhancement contributes to a reduction in cold shock damage during the freezing and thawing processes. 8 Additionally, research indicates that a reduction in SL particle size correlates with an increase in the antioxidant capacity of semen following freezing and thawing. 7
The incorporation of lecithin, specifically in the form of intentionally synthesized micelles, may offer numerous benefits simply by including this cryoprotectant in the extender. 8 Lecithin micelles arise from the self-assembly of phospholipid molecules, occurring spontaneously, and can be fabricated in various sizes, functioning effectively as stable nanoparticles. 9 Micelles exhibit significant colloidal stability. 10 Consequently, given that nanomicelles (NMs) fall within the nanoscale range, it is likely that they possess a reduced particle size along with an enhanced capacity for solubilization. 9 There are studies that show adding lecithin 11 and NMs of lecithin8,12 to the semen extender improves the cryoprotection of goat semen.
A significant advancement in the formulation of effective freezing extenders is the incorporation of liposomes within the extender. The preparation of these liposomes involves determining the optimal composition and concentration of phospholipids, which are the active constituents of SL, thereby providing essential protection to cells during the cryopreservation process.13–15 There are also methods that can be used to make particles in the form of liposomes from lecithin. Liposomes are able to coat sperm.16,17 Unlike low-density lipoproteins, liposomes can be industrially produced and easily sterilized. 18 Nanoliposomes (NLs) are a rich source of phospholipids, including phosphatidylcholine, and they enclose and release hydrophilic and hydrophobic compounds, separately or simultaneously. 19 NLs share many physical and chemical similarities with conventional liposomes, but NLs have a greater surface area and, as a result, greater solubility and stability, greater bioavailability, and more precise access to target areas. 20 Therefore, the objective of this study was to compare characteristics of frozen–thawed bull sperm after adding three concentrations of either NL or NM to the extender.
Materials and Methods
SL (L-a-phosphatidylcholine; P3644) was purchased from Sigma Chemical Co. (St. Louis, MO, USA), whereas all other chemicals were purchased from Merck (Darmstadt, Germany).
Nanoliposome and extender preparation
NLs were prepared using a thin-film procedure, as described. 21 In brief, SL in the required percentages of 1–3% (w/v) was dissolved in 5 mL of chloroform, and the resulting solution was put in a rotary evaporator at 40°C under vacuum for 2 hours to evaporate the solvent, leaving a thin lipid film. Samples were placed overnight under a vacuum in a desiccator to remove residual chloroform. The lipid film was gradually hydrated with a Tris-based buffer (249.29 mM Tris, 69.38 mM fructose, and 88.48 mM citric acid) and the emulsion slow shaken for 1 hour at 40°C. To achieve particle size < 100 nm, the emulsion was transferred to a sonicator (Mechanical Probe Sonicator, Topsonics, Tehran, Iran) for 45 minutes and then stored at 4°C. 22
Mean diameter and characteristics of the nanoliposomal formulation were determined based on dynamic light scattering in a Zetasizer system (Zeta Plus, Brookhaven Instruments Corp., Holtsville, NY, USA). NL characteristics, including size distribution, shape, and particle aggregation, were assessed with transmission electron microscopy (TEM) using negative staining with 1% phosphotungstic acid. A 20-μL drop of the sample was placed on a Formvar Carbon film coated on a 300-mesh copper grid (EMS) for 2 minutes. Excess liquid was absorbed with filter paper, then the sample was negatively stained with a 20 μL drop of 2% uranyl acetate for 1–2 minutes; excess liquid was absorbed with filter paper, and the grid was allowed to air dry. Grids were examined on a Zeiss EM10C TEM (Zeiss Em10C, Oberkochen, Germany) operating at an accelerating voltage of 100 kV.
The extender contained 249.29 mM Tris, 69.38 mM fructose, and 88.48 mM citric acid (pH = 7.2). Glycerol 5% (v/v) was added to each extender. The NM base extender was made by adding SL in the required percentages of 1–3% (w/v) to the Tris buffer at 15°C. 23 In order to get a micelle emulsion of lecithin, the suspension was placed in slow shaking for 15 minutes. For preparing the micelle with nano range particle size (<100 nm), the emulsion was sonicated (mechanical probe sonicator, Ultrasonic technology development TOPSONICS, Tehran, Iran) for 45 minutes in an ice water bath. In order to remove the remaining titanium, the medium was filtered with a 0.22 µm pore syringe, then stored at 4°C until incorporated into semen extender.
Sperm preparation and evaluation
Semen collection
Semen was collected with an artificial vagina from five mature (2 to 4 years old) Holstein bulls twice weekly for 6 weeks. All experimental procedures were performed according to the Tehran University of Ethics Committee Rules (process number:
Semen extension and freezing
Semen was extended with each extender (final concentration, 20 × 106 sperm/mL) at room temperature (∼25°C) for 5 minutes, then cooled to 4°C over 4 hours and loaded into 0.5 mL French straws (Biovet, L’Agile, France). 22 Straws were sealed with polyvinyl alcohol powder, placed 5 cm above liquid nitrogen for 12 minutes, and then plunged into liquid nitrogen. After ≥ 8 weeks, straws were thawed individually at 37°C for 30 seconds in a water bath, and semen was expelled and evaluated. 22
Post-thaw sperm evaluation
Motion characteristics
Sperm motility was assessed as described. 24 Three straws from independent replicates were thawed in a water bath at 37°C for 30 seconds and pooled in a test tube. Semen was diluted 1:1 with Tris-buffered medium and incubated at 37°C for 10 minutes. An aliquot (5 µL) of the sample was placed on a pre-warmed Leja slide and a computer-assisted sperm analysis system (Version 12 IVOS, Hamilton-Thorne Biosciences, Beverly, MA, USA) was used to assess: total motility (TM), progressive motility (PM), linearity (LIN), straight linear velocity (VSL), curvilinear velocity (VCL), average path velocity (VAP), straightness (STR), average lateral head displacement (ALH), Wobble [WOB = (VAP/VCL) × 100], and beat-cross frequency (BCF, number of times the VCL path cross).
Plasma membrane integrity
Sperm membrane integrity was assessed by staining with eosin-nigrosin (1.67 g eosin-Y, 10 g nigrosin, and 2.9 g sodium citrate dissolved in 100 mL distilled water). Sperm smears were prepared by mixing one drop of semen with two drops of stain and spreading the mixture with another slide. Plasma membrane integrity was determined by assessing 200 sperm under phase-contrast microscopy (CKX41; Olympus, Tokyo, Japan) at ×400 magnification. Only sperm with strict exclusion of stain were considered to have an integrated membrane, whereas those with partial or complete pink staining were considered to lack an integrated membrane. 25
Plasma membrane functionality
The hypo-osmotic swelling test was used to assess plasma membrane functionality, based on the presence of curled and swollen tails of sperm exposed to a hypo-osmotic solution. The assay was performed by incubating 10 µL of semen with 1 mL of a 100 mOsm/L solution (57.6 mM fructose and 19.2 mM sodium citrate) at 37°C for 60 minutes at room temperature. 26 Mixtures were homogenized and evaluated under an inverted light microscope at ×400 magnification, with 200 sperm in at least five microscopic fields assessed. The percentage of sperm with swollen and curved tails was recorded.
Sperm abnormalities
An aliquot (10 µL) of semen was pipetted into 1.5 mL tubes containing 1 mL Hancock’s solution, prepared by mixing 62.5 mL formalin (37% formaldehyde), 150 mL sodium saline solution, 150 mL buffer solution, and 500 mL double-distilled water. Approximately 5 µL of semen was placed on a slide, a coverslip applied, and percentages of sperm with normal and abnormal morphology were determined by assessing 200 sperm under a phase-contrast microscope (CKX41; Olympus, Tokyo, Japan) at 1000 ×. Morphological abnormalities included head defects (large, small, small or no acrosome, detached head), neck and midpiece defects (asymmetrical, bent, thin, cytoplasmic droplet), or tail defects (coiled, short, broken, or duplicate, and terminal droplet).
Acrosome integrity
Acrosome integrity was assessed using Pisum sativum agglutinin (PSA). First, 5 µL of sperm suspension was added to 100 µL ethanol (purity, 96%), and 15 minutes later, 10 µL of the sperm suspension was mixed with 30 µL of PSA on a glass slide. 27 Thereafter, 200 sperm per slide were viewed at 400 × with a fluorescent microscope (BX51, Olympus) equipped with fluorescence illumination and a FITC filter (excitation at 455–500 nm and emission at 560–570 nm). Sperm heads that fluoresced green were considered to have an intact acrosome, whereas those with no fluorescence were recorded as having a damaged or disrupted acrosome.
In vitro fertilization (IVF)
Sperm for IVF trials were obtained from the 1% NL, 2% NL, and 1% NM treatments (selected based on quality assessments, as described in Results). Bovine ovaries were recovered from a local abattoir and transported to the laboratory in Dulbecco’s phosphate-buffered saline solution (PBS) (at 25°C to 35°C). At the laboratory, follicles (2–8 mm in diameter) were aspirated with an 18-gauge, thin-wall needle connected to a 10-mL syringe. Cumulus-oocyte complexes (COCs) were isolated and cultured in Hepes-buffered TCM 199, penicillin-streptomycin 100 IU/mL, and 10% (v/v) fetal calf serum (FCS) for 24 hours in 5% CO2 at 39°C and 100% humidity. Thereafter, COCs with 4–10 layers of granulosa cells and oocytes with a uniform cytoplasm, homogenous distribution of lipid droplets in the cytoplasm, and an average outer diameter of ∼90 µm were selected and washed four times in fertilization medium (FertTALP) before being transferred in groups of 40–50 into 200-μL drops of fertilization medium. Three straws of frozen sperm representing one treatment were thawed (37°C, 40 seconds) and pooled. Motile sperm were obtained by centrifugation at 250 ×g on PureSperm (Nidacon; Gothenburg, Sweden) for 10 minutes at room temperature, with 2 × 106 sperm/mL transferred from the pellet to each droplet of fertilization medium containing matured COCs, and incubated for 24 hours in 5% CO2 in humidified air at 39°C. Cumulus cells were removed from presumptive zygotes by vortexing in 2 mL PBS for 1 minute. In vitro embryo culture was done in modified SOF medium containing FCS (10% v/v) under paraffin oil in a humidified atmosphere of 5% CO2 and 90% N2 at 39°C. Cleaved zygotes and embryos reaching the blastocyst stage were recorded at 3 and 7–8 days post-insemination, respectively.
Statistical analyses
The normality of data distributions was assessed with univariate procedures; all sperm endpoints were confirmed to have a normal distribution. To determine the effects of various concentrations of NL or NM on post-thaw sperm quality, all data were analyzed using Proc GLM of SAS 9.1 (SAS Institute, Version 9.1, 2002, Cary, NC, USA). All tests were performed in six replicates, and differences were located with Duncan’s multiple range tests. Results were expressed as mean ± SEM percentage. To predict the optimal concentration of NL or NM in extender, the first and second degree orthogonal methods for concentration were used, as follows:
Yij = μ + Ti + eij, where Yij: the value of each observation, μ: the mean effect, Ti: the effect of experimental treatments, and eij: the residual effects.
Results
Sperm in 2% NL had the highest (p < 0.05) TM (69.8%) and PM (58.9%; Table 1), with optimal concentrations of NL for these end points calculated as 1.88 and 1.89%, respectively. Sperm motility end points VCL, VAP, LIN, and VSL were all highest (p < 0.05) for 2% NL, whereas STR, WOB, and BCF were numerically highest in 2% NL (not significantly different from 1% NL but significantly higher than in all other groups).
Effects of Different Concentrations of Nanoliposomes or Nanomicelles on the Kinematic Parameters of Post-Thaw Bull Sperm
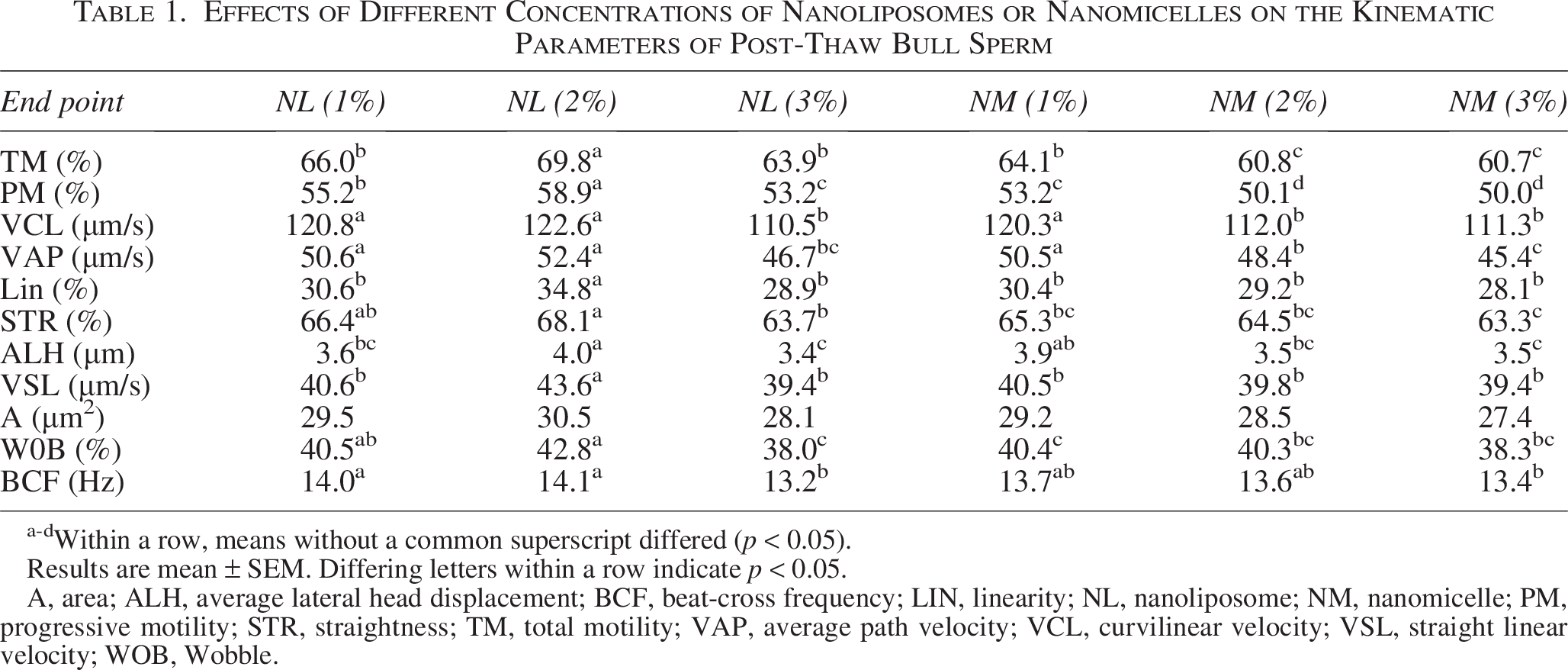
a-dWithin a row, means without a common superscript differed (p < 0.05).
Results are mean ± SEM. Differing letters within a row indicate p < 0.05.
A, area; ALH, average lateral head displacement; BCF, beat-cross frequency; LIN, linearity; NL, nanoliposome; NM, nanomicelle; PM, progressive motility; STR, straightness; TM, total motility; VAP, average path velocity; VCL, curvilinear velocity; VSL, straight linear velocity; WOB, Wobble.
The average size of NL particles (73.7 nm) was smaller (p < 0.05) than the average size of NM particles (75.2 nm), although the average zeta potential of both was −19.
Sperm cryopreserved in 2% NL had the highest (p < 0.05) cell membrane integrity and cell membrane functionality (Table 2), with calculated optimal concentrations of 1.89 and 1.88%, respectively, for NL. Furthermore, sperm cryopreserved in 2% NL had the highest (p < 0.05) intact acrosome and numerically the highest percentage of morphologically normal sperm (no significant difference among groups).
Effects of Different Concentrations of Nanoliposomes or Nanomicelles on CM Functionality (HOS-Test), and CM Integrity (Eosin-Nigrosin), Sperm with Intact Acrosome and Morphologically Normal Sperm of Post-Thaw Bull Sperm

a-dWithin a row, means without a common superscript differed (p < 0.05).
Results are mean ± SEM. Differing letters within a row indicate p < 0.05.
CM, cell membrane; HOS, hypo-osmotic swelling.
Mean Values for Development following in Vitro Fertilization for Bull Sperm Extended in Nanoliposome or Nanomicelles

a,bWithin a column, means without a common superscript differed (p < 0.05).
In the IVF studies, rates of blastocyst formation and cleavage were highest (p < 0.05) in 2% NL (40.1 and 68.0%, respectively), lowest (p < 0.05) in 1% NM (26.1 and 46.7%), and intermediate in 1% NL (28.7 and 58.1%) (Table 3).
Discussion
In this study, the effect of various concentrations of NL and NM with different thermophysical properties on the survival and fertility of bovine spermatozoa after freezing and thawing has been investigated. In the present study, NL extender resulted in significantly better motility endpoints (TM and PM) and sperm kinetics compared to NM extender. Specifically, 2% NL had the significantly highest total and progressive motility, sperm with an integrated membrane, sperm with an active membrane, sperm with an intact acrosome, and, in an IVF system, rates of blastocyst formation and cleavage. Therefore, NL provided better protection than NM for bull sperm during the freezing-thawing process, with 2% NL being the optimal concentration of the three concentrations tested.
The quality of frozen–thawed sperm can be affected by the kind and concentration of compounds in the extender. In the present study, 2% NL extender resulted in the highest TM and PM for frozen–thawed sperm. Similarly, SL-based extenders improved TM and PM, as well as the speed of sperm movement, 28 with the best sperm motility in extenders with 2% SL nanoparticles. 8
All sperm kinetics were significantly (or at least numerically) best in 2% NL. Similarly, a liposome-based extender improved sperm motility and kinetic parameters. 29 Post-thaw longevity and sperm kinetics (VAP, VSL, VCL, and ALH) improved during freezing and thawing in liposomes. Furthermore, sperm frozen with liposome travelled a significantly longer distance in the cervix compared to other treatments. 17 In yet another study, VAP, LIN, WOB were optimized in Triladyl®, whereas the highest VCL was in semen extended in Optixcell® or Andromed®30 However, there was no significant difference between lecithin and SL NL for VSL, VCL, VAP, ALH, STR, BCF, or LIN. 31 Cold damage to sperm mitochondria and axoneme during freezing and thawing decreases VCL values. 32 Both VSL (63.5) and VCL (115.1) were highest in liposome-based extender, whereas VAP (81.7) and BCF (24.1) were highest in soy lestin-2 treatment, and LIN was highest in soy lestin-1 and soy lestin-2. 33 Apparent differences among extenders were attributed to the quality and combinations of extenders.
In the present study, sperm extended in 2% NL had the highest cell membrane activity, membrane integrity, acrosome integrity, and percentage of morphologically normal sperm. Furthermore, NL increased both integrity and activity of the plasma membrane, as well as morphology compared with the NM extender. In previous studies with frozen-thawed caprine semen, the highest activity and integrity of the sperm membrane was in 2% NL (NM-based), whereas the best sperm morphology was in the 3% nanomicell. 8 Furthermore, membrane activity was highest in the 15 μM Quercetin-loaded NL group, whereas morphology was optimized in the 20 μM Quercetin-loaded NLs. 34 There was greater acrosomal and plasma membrane integrity with less lipid membrane disruption in liposome-based Optixcell extender compared with egg-based Triladyl extender. 30 Plasma membrane integrity was improved in liposome-based extender,12–14,29 and normal acrosomes were optimized with Zwitterionic liposome. 35 It has been reported that sperm with an intact acrosomal membrane were optimized in a SL-based extender. 36
Maintaining integrity of the cell plasma membrane through liposomal protection and transfer and replacement of liposome phospholipids in the sperm membrane improved sperm membrane stability and integrity during freezing and thawing.16,37–39 Phosphatidylcholine from plant sources (SL) contains unsaturated fatty acids that interact with the sperm membrane during freezing and thawing, increasing membrane fluidity and flexibility, which increase cell motility.40,41 However, during cold shock, the sperm plasma membrane releases phospholipids into the surrounding environment. 40 Cold shock-induced damage is related to the level of lipid transport, which causes sperm membrane perforation.8,13,24 The proposed mechanism to explain the protective effects of liposomes is reversible binding of liposome phospholipids to the sperm plasma membrane and coating of sperm with liposomes during freezing, 38 replacing some phospholipids of the sperm plasma membrane and thereby maintaining membrane function. 21 The nano-lecithin formulation could act via coating the sperm plasma membrane 37 or by the interaction with other extender components. In another study, the implementation of lecithin NM-based extender with 2.5 mM GSH reduced damage to bovine sperm during cryopreservation. 14
Liposomes have been employed as cryoprotective agents in various animal species, including horses, 42 buffalo, 17 rams, 43 pigs, 44 and cattle, 16 with observed enhancements in fertility following artificial insemination. 45 It is suggested that liposomes, due to their composition of phospholipids (such as phosphatidylserine, dioleoylphosphatidylcholine, phosphatidylcholine, dipalmitoylphosphatidylcholine, and dimyristoylphosphocholine) along with both saturated and unsaturated fatty acids, have the ability to merge with the sperm plasma membrane, thereby mitigating the damage inflicted on sperm during the freeze–thaw cycle. 46
It seems a possible explanation for the improved sperm kinetics by NL, but not NM is that, structurally, the bilayer architecture of liposomes allows them to act as “membrane patches” that can fuse with and repair the sperm membrane, while also providing a versatile platform for the sustained delivery of both hydrophilic and hydrophobic protectants. In contrast, the monolayer structure of micelles limits them to simple hydrophobic cargo delivery without the capacity for membrane integration or comprehensive protection. This fundamental structural difference explains the superior efficacy of liposomes in preserving sperm kinetics during cryopreservation. In an IVF system, the highest rate of blastocyst formation and cleavage was with 2% NL. In a previous study, sperm in a liposome-based expander had a fertility rate of 65%. 29 The acrosome reaction is essential for fertilization and must occur in close proximity to the oocyte. Sperm fertility is directly related to the percentage of sperm with intact acrosomes, 47 and a normal acrosome is required for an acrosome reaction, facilitating fertilization. 48 An IVF system is reliable for predicting fertility, as it evaluates interactions of sperm and egg that largely mimic in vivo conditions. 49 Liposomal delivery of phospholipids to the sperm membrane can enhance sperm motility and oocyte penetration. 50 Furthermore, greater sperm membrane fluidity accelerated activity of key enzymes such as plasma membrane ATPases and facilitated membrane binding at fertilization. 51 The mannose ligand receptor is a protein with a major role in identifying and binding sperm to the transparent layer; lecithin altered the expression of these receptors on human sperm and increased the ability of sperm to bind to the transparent layer. 41
Due to limits in ovary availability, we could not use IVF to assess all six experimental groups. However, as the optimal concentration of NL was calculated as 1.62–1.92%, it is suggested that future research should continue with 1–2% NL and fertility tested in vivo.
Conclusions
The use of an extender with 2% NL optimized protection of the sperm membrane against damage caused by freezing and thawing, and it also optimized motility end points, sperm speed, and sperm morphological characteristics.
Authors’ Contributions
M.A.A. performed the investigation and formal analysis and prepared the original draft of the manuscript. A.T. contributed to conceptualization, supervision, project administration, validation, and review and editing of the manuscript. T.N. participated in the investigation and formal analysis and contributed to review and editing. M.Z. provided supervision and validation. A.M.S. contributed to supervision and review and editing. J.P.K. contributed to review and editing. All authors reviewed and approved the final version of the manuscript prior to submission.
Footnotes
Acknowledgments
We thank the NDJ Company for providing bull semen.
Author Disclosure Statement
None of the authors have any conflict of interest to disclose that will affect the work reported in this article.
Funding Information
All the authors confirm that their research is supported by an institution that is primarily involved in education or research. This work was supported by the University of Tehran (7108017/6/43).