
Abstract
Background
Alzheimer's disease (AD) is a chronic neurodegenerative disease and the most prevalent form of dementia. Fenugreek seeds possess anti-inflammatory and antioxidant effects, making them valuable therapeutic agents in managing neurodegenerative diseases.
Objective
This study aimed to investigate the primary biological pathways and specific mechanisms underlying the protective effects of fenugreek in preventing mice of AD by employing bioinformatics and experimental verification.
Methods
We administered fenugreek extract as an intervention in mice model of AD and then assessed their cognitive ability and histopathological changes. We predicted the key target genes associated with fenugreek action on AD and the main biological pathways using the bioinformatics method. Furthermore, we observed the different expression of target genes by western blot (WB).
Results
The bioinformatics analysis revealed a strong correlation between fenugreek and AD. The behavioral experiments confirmed that fenugreek improved the behavioral and cognitive dysfunction in mice with AD. The histopathology revealed significant changes that fenugreek can inhabit Nissl bodies. Western blot experiments confirmed that fenugreek exerted statistically significant modulatory effects on the levels of inflammatory proteins [interleukin-6 (IL-6), IL-10, and IL-1β] and oxidative stress-related proteins (amyloid-β protein precursor, apolipoprotein E, and presenilin 1).
Conclusions
This study suggested that fenugreek might be involved in the AD pathway and effectively prevented the progression of AD through significant anti-inflammatory and antioxidant effects.
Introduction
Alzheimer's disease (AD) is a debilitating neurodegenerative disease associated with aging and is the leading cause of dementia in older adults. 1 The clinical symptoms encompass cognitive impairments such as memory loss, impaired judgment, and reduced abstract thinking abilities. 2 GBD 2019 Dementia Forecasting Collaborators predict that there will be 152.8 (130.8–175.9) million individuals living with dementia in 2050. 3
AD is a complex, multifactorial condition with several main etiological hypotheses: the “amyloid hypothesis”, “the tau hypothesis”, and the “cholinergic defect hypothesis”.4,5 The primary pathological characteristics of AD include the deposition of amyloid-β (Aβ) and the hyperphosphorylation of tau proteins, which contribute to the accumulation of neurofibrillary tangles. 6 These changes lead to substantial synaptic disruption and neuronal degeneration. 7 It has been proposed that oxidative stress occurs early in AD,8,9 and this is supported by the detection of oxidative markers in brain regions affected by mild cognitive impairment.10,11 As oxidative stress is interrelated with Aβ aggregation, it plays a role in inducing neurological damage and memory loss. 12 In summary, the therapeutic approaches for AD focus on inhibiting acetylcholinesterase activity and reducing oxidative stress.13,14
The primary drugs used in clinical practice are cholinesterase inhibitors and N-methyl-D-aspartate antagonists, which alleviate the symptoms of AD without providing a cure or prevention for the disease. 4 In March 2021, after an 18-year gap, the Food and Drug Administration (FDA) approved aducanumab for treating AD. According to the prescribing information provided by the FDA, the drug should be administered during the mild dementia stage of individuals diagnosed with AD. 15 This marked the development of a disease-modifying class of compounds to target Aβ for treating AD. However, its therapeutic efficacy remains controversial. 16 Currently, no cure is available for AD, and initiating treatment at the onset of symptoms may prove to be ineffective and untimely. This study aimed to explore the pathogenesis of mice model of AD in its early stages and mitigate or postpone the disease progression through early intervention. Therefore, it is crucial to focus not only on AD treatment but also on primary AD prevention.
Fenugreek, scientifically known as Trigonella foenum-graecum or Hilbeh in the Middle East, contains more than 100 phytochemicals, primarily including polysaccharides, saponins, alkaloids, phenolic acids, and flavonoids. 17 Extensive preclinical and clinical studies have confirmed the antidiabetic, antihyperlipidemic, anti-obesity, anticancer, anti-inflammatory, antioxidant, and antibacterial properties of fenugreek such as.18–21 A randomized controlled trial demonstrated that fenugreek seed extract significantly improved the memory and quality of life of patients with AD. 22 Animal experiments demonstrated that fenugreek seed powder significantly reduced AlCl3-induced memory deficits, amyloid and tau pathology, oxidative stress, and inflammation in mice model of AD. 19 Therefore, fenugreek holds significant research and application values in preventing AD.
The network pharmacology in combination with amyloid precursor protein (APP)/PSEN (Presenilin) double-transgenic mice of C57BL/6 background was used to explore the potential mechanism of fenugreek against AD. This study used bioinformatics to screen for potentially effective monomers derived from fenugreek. The core cross-gene targets of fenugreek and AD were screened using the GeneCards database. Protein–protein interaction (PPI), Gene Ontology (GO), and Kyoto Encyclopedia of Genes and Genomes (KEGG) were used to analyze the potential associations among gene targets and investigate the mechanism of action and potential pathways. A standard network pharmacology approach was employed to predict the mechanism of the effects of fenugreek on AD supported with behavioral, morphological, and protein semi-quantitative analyses in mice model of AD. This study aimed to facilitate the clinical application of fenugreek by exploring the mechanism of action for AD prevention.
Methods
Animals
The specific-pathogen-free (SPF) grade C57BL/6 mice used as wild-type controls were purchased from Chengdu Dashuo Laboratory Animal Co. The model animals used to examine the behavioral and neurological damage in AD were 8- to 10-week-old male APP/PSEN double-transgenic mice of C57BL/6 background, purchased from Jiangsu Ailingfei Biotechnology Co. After quarantine, all mice were housed at a room temperature of 22 °C–26 °C and a relative air humidity of 40%–60% in an SPF animal housing room. They had free access to food and water, with bedding made of poplar wood shavings that was changed regularly. They were placed under alternating 12-h light and 12-h dark cycles for 1 week to acclimatize.
The animal study was reviewed and approved by the Ethics Committee of Chengdu Medical college.
Experimental interventions
In this study, we used the traditional Chinese medicine fenugreek extract (a gift from Guo Feng, Anhui Agricultural University) made from fenugreek seeds. The seeds were refluxed at 70 °C with 50% ethanol and passed through a 50-nm inorganic ceramic composite membrane. The ethanol was removed, and the seeds were concentrated using a 20-kDa organic molecular sieve membrane, followed by lyophilization. The purity of fenugreek saponins in the extract was determined to be approximately 40.47%. Twenty 8- to 10-week-old male APP/PSEN1 transgenic model mice (25–30 g) were randomly categorized into the AD model group, AD + high-concentration fenugreek extract solution group, AD + medium-concentration fenugreek extract solution group, and AD + low-concentration fenugreek extract solution group, with five mice in each group. Five randomly selected 8- to 10-week-old male C57BL/6 mice (25–30 g) were used as the control group. The mice in the AD + high-concentration fenugreek extract group were given 5 mg/mL fenugreek extract solution (1 mL) in 0.9% sodium chloride via gavage daily for 60 days. The mice in the AD + medium-concentration fenugreek extract group were given 3.75 mg/mL fenugreek extract solution (1 mL) in 0.9% sodium chloride via gavage daily for 60 days. The mice in the AD + low-concentration fenugreek extract group were given 2.5 mg/mL fenugreek extract solution (1 mL) in 0.9% sodium chloride via gavage daily for 60 days. The mice in the control and AD model groups were given 0.9% sodium chloride solution (1 mL) via gavage daily for 60 days.
Morris water maze test
The Morris water maze test analyzed the learning and memory capability for spatial position and direction sense in mice. This test was conducted in two stages: the experimental training stage and the exploratory experiment stage. The water maze bucket was divided into four quadrants corresponding to four water entry points at the upper edge in the experimental training stage. The mice were put into the water from one of the entry points facing the pool wall. The escape latency, which was the time swimming to the platform and standing on the platform, was recorded for five consecutive days. The swimming speed was calculated simultaneously. After the training experiment, the exploratory experiment test was conducted. The platform was removed, and the mice were placed in water. The times of crossing the position of the initial platform and staying in the quadrant of the original platform within 60 s were recorded to test the memory of mice with the original platform. Morris water maze tests were performed in a silent environment from 9:00–12:00 a.m. on the second day of the open-field test. The Ganz FC62D video camera (HVS Image, UK) was used for recording the relevant parameters of the Morris water maze.
Hematoxylin–eosin staining
Half of the hippocampus was fixed with tissue fixation solution (P1110; Solarbio, China), embedded in paraffin, and sliced into 5-μm sections. After dewaxing, the sections were stained with hematoxylin and eosin (H&E), washed with water for 3–5 s, gradient dehydrated with alcohol (75%–85%–100%), made transparent with xylene, and sealed with neutral balsam mounting medium (E675007; Sangon, China). The images were recorded using the trinocular microscope system (BA200Digital; Mike Audi, China). Each section was observed at a lower magnification of 100×, and three fields were selected to collect 400× microscopic images to observe specific pathological conditions.
Nissl staining
We determined the number of Nissl bodies in the hippocampus and investigated the neuroprotective effect of the fenugreek extract solution on AD. Nissl staining was performed following the manufacturer's protocols. Then, 5-μm coronal sections were deparaffinized in xylene and rehydrated using ethanol. The slides were stained with Nissl staining solution (G1036; Servicebio; China) at 37 °C for 5 min. Five randomly selected fields in the ipsilateral cortex of each animal were analyzed in five nonadjacent sections. A Panoramic 250 digital section scanner manufactured by 3DHISTECH (Hungary) was used for image acquisition of the sections. Each group of sections was used to acquire 400× images of the CA1 region of the hippocampus to observe the general lesions in the hippocampal region.
Western blot analysis
The protein expression of apolipoprotein E (APOE), amyloid precursor protein-like (APPL), interleukin-1β (IL-1β), IL-6, IL10, and PSEN in the hippocampus was detected by western blot (WB) analysis. Tissues were homogenized in radioimmunoprecipitation assay buffer (Beyotime, China) and lysed on ice for 10 min. The lysate was centrifuged at 5000 g at 4 °C for 10 min to obtain the supernatant. The total protein content was confirmed using the bicinchoninic acid protein quantification kit (Beyotime). After quantification, the denatured protein was separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto polyvinylidene difluoride (PVDF) (Sigma–Aldrich, USA, #ISEQ07850). The following primary antibodies were obtained from Cell Signaling Technology (Zen-Bio, China) and used to incubate PVDF membranes at 4 °C overnight: β-actin (1:4000; Zen-Bio, #700068), APOE (1:1000; Zen-Bio, #R381129), APP (1:1000; Zen-Bio, #R23511), PEN1 (1:1000; Zen-Bio, #R25440), IL1β (1:1000; Zen-Bio, #350193), IL6 (1:1000; Zen-Bio, #500286), IL10 (1:1000; Zen-Bio, #502171). HRP-conjugated secondary antibodies (1:1000; Beyotime, China, #A0208, #A0216;) were incubated for an immune response at room temperature for 2 h. The proteins were analyzed using the enhanced chemiluminescence detection kit (Bio-Rad, USA) and Tanon 5200 (Shanghai, China).
Acquisition and screening of AD gene targets
In this study, we used “Alzheimer's disease” as a keyword to obtain disease gene targets from the GeneCards database. A correlation score of ≥5 was used as a threshold for screening AD-related gene targets from the GeneCards database, which is a comprehensive assessment of the association of a gene with the disease under study.
Identification and screening of active compounds and gene targets
All compounds in fenugreek were screened and analyzed using the Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP, http://tcmspw.com/). 23 Fenugreek were screened in the absorption, distribution, metabolism, and excretion assessment categories based on two key parameters: oral bioavailability (OB) and drug likeness (DL). OB was defined as the extent to which the body used the active ingredient. 24 OB largely determined the effect of compounds on diseases, whereas DL was used to screen and improve candidate compounds in an early stage of drug development. In this study, the active compounds in fenugreek were selected based on the criteria of OB ≥30% and DL ≥0.18. We imported the screened 15 herbal monomers into the GeneCards database to obtain pharmacogenetic targets.
Intersection of disease and drug–gene targets
The drug–gene targets were combined with disease–gene targets through the Venny website to obtain crossover gene targets.
Protein–protein interaction network construction
The Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database was used to analyze PPIs of fenugreek for AD and construct PPI networks. 25 We imported AD protein targets obtained from the GeneCards database into Cytoscape 3.7.1 and the STRING database for analysis. 26 Cytoscape 3.7.1 was used to analyze the network topology parameters and filter the core protein targets based on the criteria of node degree value and median centroid value greater than the mean.
Gene target enrichment analysis
We performed GO and KEGG enrichment analysis using relevant gene targets from the Database for Annotation, Visualization, and Integrated Discovery (DAVID) database. GO enrichment was used to obtain biological information about gene targets in biological processes (BPs), cellular components (CCs), and molecular functions (MFs). KEGG pathway enrichment was performed by enriching the signaling pathways involved in the relevant gene targets of the KEGG pathway. The OmicShare tool platform was used to visualize the results of GO and KEGG enrichment.
“Disease–core target gene–drug” network diagram
The disease–core target gene–drug network was constructed and topologically analyzed using Cytoscape 3.7.1 software. The top 10 KEGG pathways and their key genes, along with the top 15 active components of fenugreek, were imported into Cytoscape 3.7.1 software. Core gene targets were identified by screening for nodal degree values greater than twice the median.
Statistical analysis
SPSS software was used to calculate significance. Comparison of two groups was performed by t-test. Multiple comparisons were performed by One-Way Analysis of Variance (ANOVA) followed by Bonferroni post hoc test for normally distributed samples or ANOVA on RANKS followed by post hoc tests for non-normally distributed samples. p values < 0.05 were considered as significant in all tests. Statistical studies were performed using the GraphPad Prism statistical package (GraphPad Prism 5.0; GraphPad Software, Inc., San Diego, CA).
Results
Morris water maze test
The Morris water maze experiment was conducted to investigate the effects of fenugreek extract on the behavioral activity and memory learning ability of mice in each group (Figure 1). The experiment was repeated three times on days 1, 30, and 60.

Morris water maze test results. Experimental intervention on the first day: Results of escape latency during the platform concealment training period (A) and results of the platform latency and crossing times during the probe trial (D). Experimental intervention on the 30th day: Results of escape latency during the platform concealment training period (B) and results of the platform latency and crossing times during the probe trial (E). Experimental intervention on the 60th day: Results of escape latency during the platform concealment training period (C) and results of the platform latency and crossing times during the probe trial (F). nsp > 0.05; *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
The escape latency of mice in the platform concealment training phase was mainly used to assess the learning ability of mice. The results of the first experiment demonstrated no significant difference in the escape latency between the AD model and control groups on each day of the platform concealment training phase (p > 0.05). Similarly, no significant difference was found in the escape latency between each fenugreek extract solution intervention and the control group (p > 0.05). Further, no significant difference was observed in the escape latency between the intervention (p > 0.05) and model groups (p > 0.05). Similarly, the number of times crossing the original platform position within 60 s was not significantly different between the groups during the experimental phase of the no-platforming detection period (p > 0.05). The results of the second experiment revealed that, in the experimental phase of the platform concealment training period, the evasion latency of mice on the first day significantly increased in the AD model group compared with the control group (p < 0.05), the evasion latency of mice in the AD model group was also significantly different from that of mice in the AD + high-concentration fenugreek extract group (p < 0.05), and the evasion latency of mice in the AD + high-concentration fenugreek extract group was also significantly different from that of mice in the control group (p < 0.05). The aforementioned intergroup differences gradually increased until the fifth day of the experiment, but no significant difference was found between the AD model, medium-concentration fenugreek extract, and AD + low-concentration fenugreek extract groups (p > 0.05).
H&E staining
The pathomorphological results of the hippocampal tissues of the mice in each group were observed by H&E staining, followed by microscopy at 100× and 400× (Figure 2).
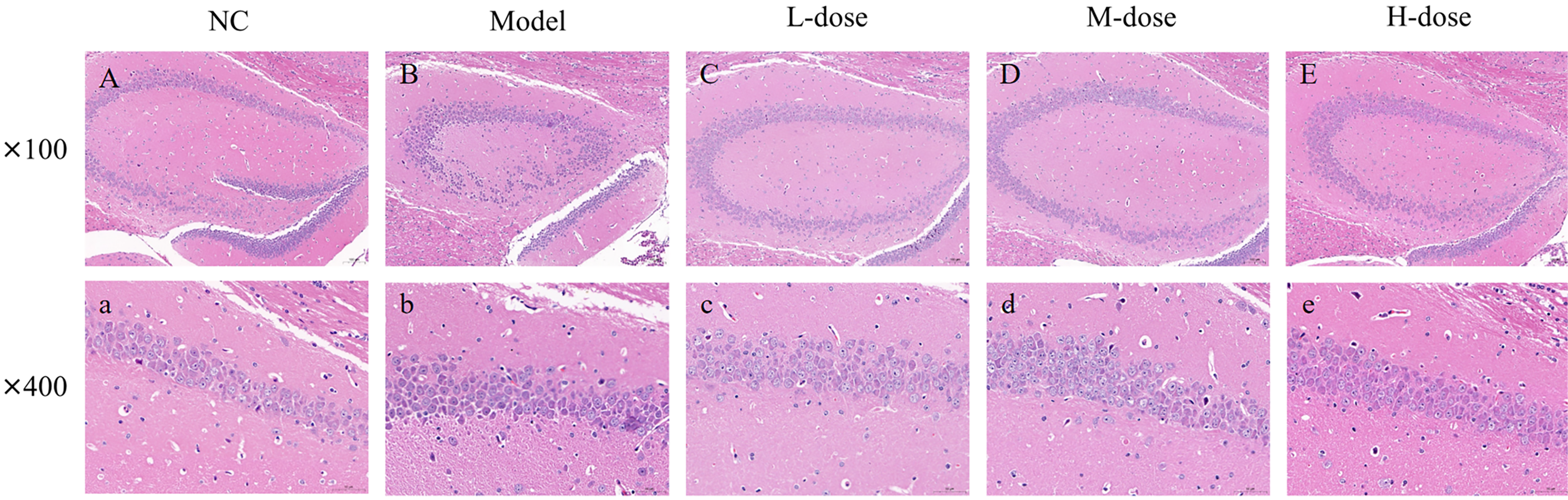
H&E staining of the mouse hippocampal tissue. (A) Blank control group (100×); (a) blank control group (400×). (B) AD model group (100×); (b) AD model group (400×). (C) AD + low-concentration fenugreek extract solution group (100×); (c) AD + low-concentration fenugreek extract solution group (400×). (D) AD + medium-concentration fenugreek extract solution group (100×); (d) AD + medium-concentration fenugreek extract solution group (400×). (E) AD + high-concentration fenugreek extract solution group (100×); and (e) AD + high-concentration fenugreek extract solution group (400×).
Following H&E staining, the pathomorphological findings of the hippocampus tissues of the mice in each group were examined using microscopy at 100× and 400×.
The regular groups are depicted in Figure 2A-(a). Each of the hippocampus's CA1, CA2, CA3, and dentate gyrus regions exhibited a unique molecular layer, a layer of pyramidal cells, and a stratification of polymorphic layers. A small number of neurons, dendrites from pyramidal cells, and other nerve fibers made up the molecular layer. Within the layer of pyramidal cells, the cone cells were closely spaced in bands, exhibiting distinct cell shape and readily visible nucleoli. No discernible glial cells were found. Also, no overt signs of inflammatory cell infiltration, glial cell growth, or other pathological alterations were noted. Figure 2B-(b) denotes the AD model groups. The color deepened, the volume of the cells was smaller, the internal structure of the cells was blurred, and the tail of the cytosol was visible with an obvious axon-like structure. Glial cell proliferation, inflammatory cell infiltration, and other pathological changes were not obvious. Figure 2C-(c) denotes the AD + high-concentration fenugreek extract solution groups. The pyramidal cell layer of the hippocampal area was neatly arranged, with cells densely packed. Occasionally, dark neurons were observed, characterized by reduced cell volume and a deepened color. Figure 2D-(d) denotes the AD + medium-concentration fenugreek extract solution groups. Figure 2E-(e) denotes the AD + low-concentration fenugreek extract solution groups, with no significant change compared with the model group.
Nissl staining
The pathological morphology of the hippocampal tissues of mice in each group was observed by Nissl staining followed by 400× microscopy (Figure 3).

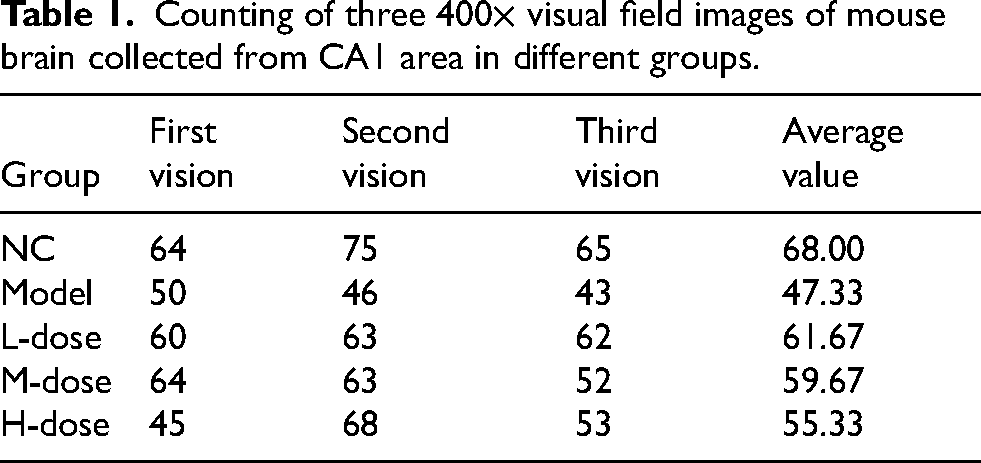
Nissl staining of hippocampal tissue of mice in different groups: (A) the blank control group (400×), (B) the AD model group (400×), (C) the AD + low-concentration fenugreek group (400×), (D) the AD + medium-concentration fenugreek group (400×), and (E) the AD + high-concentration fenugreek group (400×). The white arrow points to Nissl staining–positive neurons. Three 400× visual field images collected in the CA1 area in each group were counted (Table 1).
Counting of three 400× visual field images of mouse brain collected from CA1 area in different groups.
The experimental data of the obtained niche counts were statistically analyzed using SPSS 23.0 statistical software, and the results were expressed as mean ± SD. One-way analysis of variance was used to compare groups with the test level of α = 0.05.
Nissl staining was performed on the brain sections of mice in each group to observe the CA1 region of the hippocampus and count the Nissl microsomes. The Nissl microsome count in the hippocampal region significantly reduced in the APP/PSEN1 transgenic AD model group compared with the blank control, AD + high-concentration fenugreek extract solution, and AD + medium-concentration fenugreek extract solution groups, and the difference was statistically significant (p < 0.05). The differences between the AD + high-concentration fenugreek extract and AD + medium-concentration fenugreek extract groups and the blank control group were not statistically significant (p > 0.05). Also, the difference between the AD + low-concentration fenugreek extract and blank control groups was statistically significant (p < 0.05). The difference between the AD + low-concentration fenugreek extract and AD model groups was not statistically significant (p > 0.05). No statistically significant differences (p > 0.05) were found between the treatment groups (AD + high-concentration fenugreek extract, AD + medium-concentration fenugreek extract, and AD + low-concentration fenugreek extract groups).
Acquisition of disease-related gene targets
In this study, 12,825 AD gene targets were identified. AD-associated gene targets were retrieved from the GeneCards database based on the correlation scores. Targets with correlation scores ≥5 were considered AD-associated gene targets, resulting in the identification of 3843 AD-associated gene targets.
Identification of potentially active compounds in fenugreek
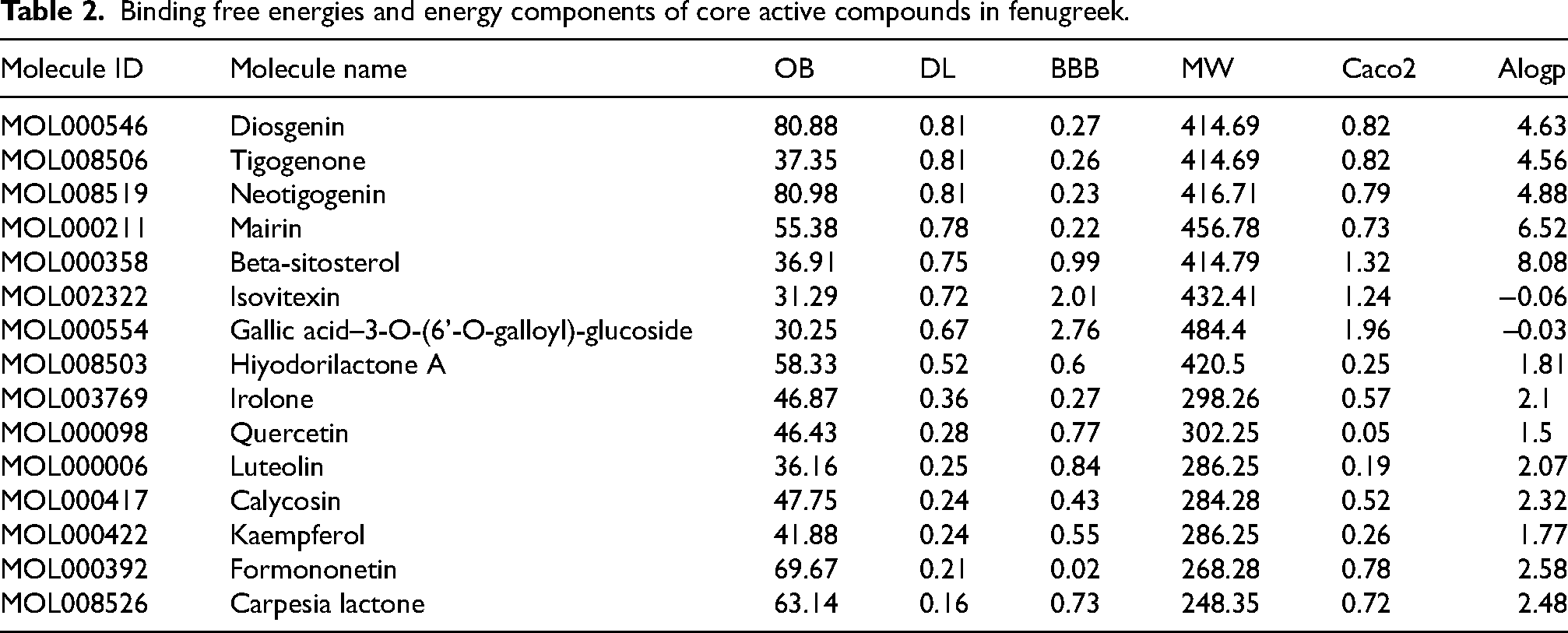
Fifty-nine potential compounds in fenugreek were retrieved from the TCMSP database, and 15 core active compounds in fenugreek were screened by further increasing the criteria to DL ≥0.18 (Table 2).
Binding free energies and energy components of core active compounds in fenugreek.
Common gene targets obtained by intersection
We obtained 12,824 AD gene targets and 1155 fenugreek gene targets. Venny processed 935 crossover gene targets (Figure 4).

Targets of the intersection of fenugreek and AD.
Construction of protein–protein interaction network
We obtained protein scores from the GeneCards database, and a relevance score ≥30 was considered a key protein target. Seventy-three key protein targets were screened using the relevance score. The PPI network of fenugreek and AD was constructed using the STRING database (Figure 5A). Fifty-eight core protein targets (e.g., AKT1, APP, STAT3, etc.) were obtained by increasing the confidence score to ≥0.95. The bioinformatics analysis indicated that the inflammatory factors occupied most of the core PPI network. Additionally, the factors involved in regulating lipid metabolism were closely associated with other targets, indicating that starch deposition and oxidative stress played a major role in AD.

(A) Protein–protein interaction (PPI) network of protein target (confidence interval >0.4) and (B) core protein target (confidence interval >0.95).
GO and KEGG enrichment analyses
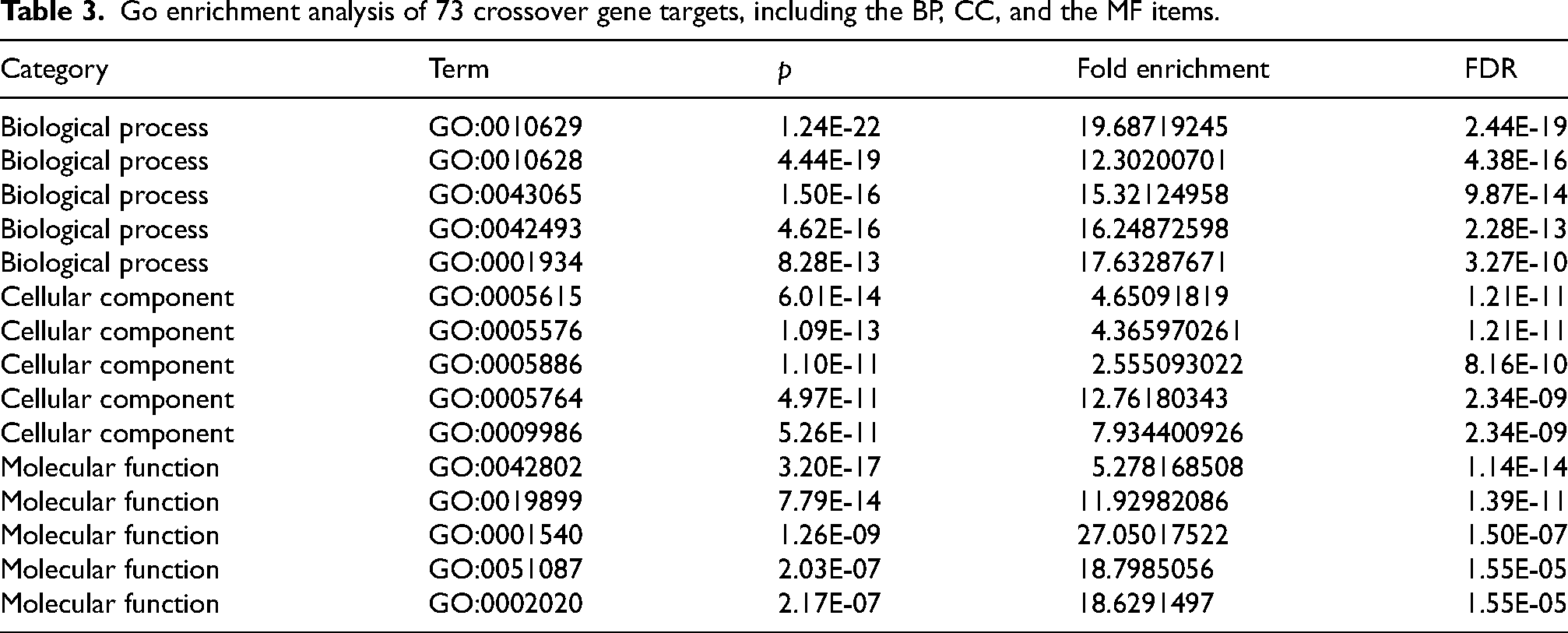
Seventy-three crossover gene targets were imported into the DAVID database for enrichment analysis. Under the p < 0.05 condition, the GO enrichment analysis yielded
273 GO entries, including 97 BP, 94 CC, and 82 MF entries. The top 10 BP, CC, and MF entries were filtered based on the number of included targets. The results demonstrated that the BPs were highly associated with inflammation and oxidative stress, mainly involving cytokine-mediated signaling pathways, inflammatory responses, and immune responses. The extracellular space, extracellular regions, and cell surfaces accounted for a relatively large proportion of the CCs. Cytokine activity, protein binding, and receptor binding were the major components of molecular function (Table 3).
Go enrichment analysis of 73 crossover gene targets, including the BP, CC, and the MF items.
KEGG pathway analysis resulted in 134 pathways with p < 0.05. The top 15 pathways were screened based on the number of targets included. The results revealed that the enriched pathways involved multiple pathways related to lipid metabolism, mainly lipid and atherosclerosis, AD, fluid shear stress and atherosclerosis, and other signaling pathways (Figure 6A). The picture on the right depicted the possible targets involved in each pathway.

(A) Top 15 KEGG pathways. (B) Subnetwork depicts top 15 KEGG pathways and related genes.
Screening and analysis of key protein targets and core monomers
The 55 screened by PPI were imported into Cytoscape software. The top 20 genes (CCL2, IL-6, TNF, IL-1β, IL-10, IFN-γ, VEGFA, STAT3, EGFR, PIK3CA, AKT1, PTEN, TP53, CTNNB1, APP, MAPT, APOE, APOA1, APOB, and PSEN1) were selected using the cytoHubba plugin, and the main biological pathways involved in the hub genes were predicted using ClueGo software (Figure 7). 27

(A) Top 20 key genes obtained by the degree value method. Each node represents a protein, and each edge represents the relationship between two proteins. The larger the node in the PPI, the darker the color, the more the node protein will interact with the surrounding proteins. (B) Six hub genes and the key biological processes they are involved in.
WB experimental verification
The effect of fenugreek on the protein expression levels of APOE, APPL, IL-1β, IL-6,
IL-10, and PSEN1 in the hippocampal tissues of APP/PSEN1 mice with AD was observed. The expression levels of APOE, APPL, IL-1β, IL-6, IL-10, and PSEN1 were significantly higher in the model group compared with the blank group (p < 0.001). The expression levels of APPL were not significantly different in the low-dose fenugreek extract group. The expression levels of IL-1β and IL-10 reduced in the fenugreek low-dose group, and the expression levels of PSEN1 increased in the low-dose fenugreek extract group and significantly decreased in the high-dose fenugreek extract group compared with the model group (p < 0.001) (Figure 8).

Protein expression of APOE, APPL, IL-1β, IL-6, IL-10, and PSEN1 in the hippocampus was detected by WB. Protein level quantification of APO (A), APPL (B), IL-1β (C), IL-6 (D), IL-10 (E), and PSEN1 (F) among NC, Model, L-Dose, M-Dose, and H-Dose groups. Data were presented as mean ± SD (*p < 0.05, ***p < 0.01, ****p < 0.001).
Discussion
Currently, the pathogenesis of AD remains complex and poorly understood. This study focused on integrating network pharmacological analysis with experimental validation to investigate the molecular mechanisms underlying the potential of fenugreek for preventing AD. The findings indicated that fenugreek might decelerate AD progression by exerting antioxidant and anti-inflammatory effects and preserving neuronal integrity in mice model. Therefore, we further hypothesize that fenugreek has a role in delaying dementia development in individuals at risk for AD or in patients diagnosed with AD, a hypothesis that needs to be further tested in clinical trials.
To investigate the underlying mechanism of fenugreek prevent in mice model of AD, network-based pharmacology was applied to predict the potential targets of fenugreek and AD in this study. And 935 potential targets of fenugreek for AD were obtained after intersection. The common genes were screened based on their relevance values, resulting in the identification of 73 key genes. GO enrichment and KEGG pathway analyses of the 73 key genes revealed their predominant localization within the extracellular matrix. Among which 20 core targets (CCL2, IL-6, TNF, IL-1β, IL-10, IFNG, VEGFA, STAT3, EGFR, PIK3CA, AKT1, PTEN, TP53, CTNNB1, APP, MAPT, APOE, APOA1, APOB, and PSEN1) stood out after more precise filtering. WB experimental validation of 6 core targets revealed that APOE, APPL, IL-1β, IL-6, IL-10, and PSEN1 were modulated in the process of fenugreek prevent in mice model of AD, suggesting that fenugreek can be used a potential drug to prevent AD through multi targets. The excessive buildup of inflammatory cytokines (IL-6 and IL-1β) and Aβ peptides (APP and APOE) in the extracellular space contributes to pathological damage, including neuroinflammation, oxidative stress, and neurotoxicity. 28 The main BP entails both negative and positive regulation of gene expression. Previous studies have demonstrated that fenugreek restores BDNF levels by inhibiting the expression of proinflammatory cytokines, 29 inhibits acetylcholinesterase activity, promotes the elevated expression of neuroacetylcholine, 30 and exerts neuroprotective effects in mice with AD, thereby improving cognitive impairment. The primary MFs were “identical protein binding” and “enzyme binding.” The interaction between related proteins and enzymes altered the structure and function of proteins, resulting in pathological phenomena. One example was ERAB, which was encoded by the human short-chain L-3-hydroxyacyl coenzyme A dehydrogenase gene and led to neuronal dysfunction. 31 The BioSign data revealed that the “Alzheimer's disease signaling pathway” was the most significant pathway involving GSK3β, APP, PSEN1, IL-6, IL-1β, APOE, SNCA, and others. APOE and APP were the key determinants of AD risk.32,33 PSEN1 plays a crucial role in memory and neuronal maintenance. 34 We performed Hubba enrichment analysis on the factors involved in the top 10 KEGG pathways using p values, identifying the top 20 key factors closely associated with neuroinflammation and Aβ protein deposition.
The onset of AD is typically preceded by a transitional period known as mild cognitive impairment (MCI), during which patients with an amnestic MCI (aMCI) can perform activities of daily living compared with those with AD. Patients with AD are characterized by learning and memory deficits, decreased higher executive functions, and reduced reasoning ability, whereas memory deficits are the hallmarks of aMCI. In both cases, the altered functions primarily result from synaptic dysfunction associated with the changes in the synaptic proteome.35,36 An estimated 15% of patients with aMCI progress to AD each year. 37 Considering that patients with MCI had a high risk of progressing to early AD and extensive oxidative damage in MCI might precede evident AD neuropathological changes, these findings strongly indicated that oxidative imbalance emerged in the early stages of AD and might play a central role in AD pathogenesis. 8
The fenugreek extract comprises numerous phytochemical compounds, including flavonoids and saponins, which are recognized as antioxidants capable of reducing oxidative stress. 20
The analysis of the raw data from this study revealed that most of the top 15 fenugreek extract solutions, which played crucial pharmacological roles in AD, belonged to the flavonoid and saponin classes. KEGG data confirmed the close interaction between APP, PSEN1, and APOE genes in the AD pathway. The mutations in the APP gene result in the accumulation of Aβ and the formation of neurofibrillary tangles, thereby contributing to the early onset of AD.33,38,39 PSEN1 is the core protein that activates the γ-secretase complex and plays a vital role in Aβ production by APP. The knockout studies of PSEN1 in mice demonstrated synaptic dysfunction and memory impairment, suggesting its critical role in memory and neuronal maintenance. 34 Sun et al. demonstrated that the mutations in the PSEN1 gene led to an elevation in the Aβ42/Aβ40 ratio through a reduction in Aβ40 levels. 40 Therefore, the therapies aimed at lowering Aβ levels or inhibiting Aβ protofibrillogenesis may mitigate the risk or prevent the onset of AD. APOE protein is a glycoprotein with high expression in the liver, brain astrocytes, and certain microglia. It functions as a receptor-mediated ligand involved in the endocytosis of lipoprotein particles, including cholesterol, which is crucial for myelin production and normal brain function. 4 The APOE gene consists of three isoforms: ApoE2, ApoE3, and ApoE4. The APOE polymorphic alleles play a significant role in determining the risk of AD. APOE influences the clearance, aggregation, and deposition of Aβ in an isoform-specific manner. 32
Previous studies have demonstrated the genetic contribution of AD to inflammation, highlighting inflammation as a potential early indicator of AD risk or pathogenesis. 41 Additionally, a characteristic of AD is the development of neuritic plaques composed of Aβ peptides (Aβ42) derived from the proteolytic processing of APP by secretases (α, β, and γ). 42 Among these, γ secretase, which mainly includes progerin 1 (PSEN1) or progerin 2 (PSEN2), further cleaves the membrane-bound fragments of amino acid residues cleaved by β secretase, resulting in Aβ1−40 and Aβ1−42 isoforms. Aβ plaques are deposited in various regions of the brain. 43
Subsequently, microglia become activated and initiate a proinflammatory cascade response, 28 involving the production of IL-1β and IL-6 by brain endothelial cells to counteract Aβ plaques.44,45 Furthermore, Culjak et al. discovered that the changes in the concentrations of IL-1α, IL-10, and TNF-α in AD partially supported the association of the neuroinflammatory response with AD. Additionally, astrocytes and microglia synthesize APOE in the central nervous system. 46 APOE, in turn, functions as an anti-inflammatory agent, regulating the inflammatory response of astrocytes and microglia. 47
Limitation
This study will select clinical AD treatment drugs as the control group and compare with fenugreek experimental group to further explore the efficacy of fenugreek.
Conclusions
This study revealed that fenugreek could prevent the pathogenic process of AD. Additionally, the administration of fenugreek delayed the onset of inflammatory and oxidative pathological changes in patients with AD prior to the manifestation of early symptoms such as recent amnesia. Surprisingly, the flavonoid and saponin components of fenugreek exhibited significant anti-inflammatory and antioxidant effects. Thus, individuals at high risk of developing AD can incorporate fenugreek into their daily diet in an early stage, consuming it in appropriate amounts or concentrations as a preventive measure.
Footnotes
Acknowledgments
We gratefully acknowledge the assistance of Chengdu Medical College and the First Affiliated Hospital of Chengdu Medical College, Chengdu, China, for the experimental platform. We also thank Professor Guo Feng of Anhui Agricultural University for providing fenugreek extract.
Author contributions
Li Xiong (Supervision; Visualization; Writing – original draft; Writing – review & editing); Qinxuan Li (Conceptualization; Data curation; Funding acquisition; Investigation; Methodology); Xuhui Zhou (Data curation; Formal analysis; Methodology; Resources); Jiujia Xiao (Data curation; Formal analysis; Funding acquisition; Investigation); Xingyu Yang (Writing – original draft; Writing – review & editing); Hengxiang Xu (Data curation; Methodology); Chuan Guo (Conceptualization; Data curation; Formal analysis; Investigation; Methodology; Project administration; Resources; Software; Supervision).
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Project of Study on the mechanism of CAPE action on WNT/β-catenin pathway in U251 human glioma in vivo, Chengdu Municipal Health Commission, 2021-2024, Subject No. 2021063; Study on the mechanism of action of caffeic acid phenethyl ester in glioma in vivo, Research Starting Fund for High-level Talents of the First Affiliated Hospital of Chengdu Medical College, 2019, Subject No. CYFY-GQ27.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
All relevant data are included within the manuscript and its Supporting Information files.