
Abstract
Mesenchymal stem cells (MSCs) are being tested in several biological systems and clinical settings with the aim of exploring their therapeutic potentials for a variety of diseases. MSCs are also known to be heterogeneous populations with variable functions. In the context of this multidimensional complexity, a recurrent question is what source or population of MSCs is suitable for specific clinical indications. Here, we reported that the biological features of MSCs varied with the individual donor, the tissue source, the culture condition and the subpopulations. Placental chorionic villi (CV) derived MSCs exhibited superior activities of immunomodulation and pro-angiogenesis compared to MSCs derived from bone marrow (BM), adipose and umbilical cord (UC). We identified a subpopulation of CD106(VCAM-1)+MSCs, which are present richly in placental CV, moderately in BM, and lowly in adipose and UC. The CD106+MSCs possess significantly increased immunomodutory and pro-angiogenic activities compared to CD106−MSCs. Analysis of gene expression and cytokine secretion revealed that CD106+MSCs highly expressed several immnumodulatory and pro-angiogenic cytokines. Our data offer new insights on the identification and selection of suitable source or population of MSCs for clinical applications. Further efforts should be concentrated on standardizing methods which will ultimately allow the validation of MSC products with defined biomarkers as predictive of potency in suitable pre-clinical models and clinical settings.
Introduction
The mesenchymal stem/stromal cells (MSCs) represent a kind of multipotent stem cells which are originated from mesoderm and able to differentiate into mesenchymal cell lineages such as osteocytes, chondricytes and adipocytes [1,2]. Friedenstein was the first to describe the isolation of clonogenic, proliferating fibroblastic MSCs from rat bone marrow (BM) [3]. Now, MSCs are known to reside in almost all tissues of the body, where they undergo self-renewal and multi-lineage differentiation. Besides BM, MSCs can be isolated mainly from adipose tissue and perinatal tissues (umbilical cord, cord blood, amniotic membrane and placenta) [4–7].
A number of studies have demonstrated that MSCs play important roles in immune and hematopoietic modulation, anti-inflammation, pro-angiogenesis, as well as tissue repair/restoration. These unique characteristics of MSCs make them ideal candidate for stem cell-based therapy, tissue engineering and regenerative medicine in many clinical settings. Increasing evidence indicates that MSCs exert their therapeutic effects mainly through inhibition of undesirable immune reactions and secretion of growth factors rather than the direct replacement of damaged cells [8,9]. Most important, it has been proven that MSCs are safe, do not form teratoma [10,11], and easy to be isolated, expanded, cryopreserved, and large-scale prepared as a drug for clinical uses in GMP-compliant facility.
To date, over 650 clinical trials involving the applications of MSC have been registered, among 45% trials used bone marrow-derived MSCs (BMSCs), while others used adipose-(AMSCs), umbilical cord-(UMSCs), placenta-(PMSCs), or cord blood-(CBMSCs) derived MSCs (see www.clinicaltrials.gov). Most clinical trials on BMSCs and AMSCs are personalized treatment using autologous MSCs, whereas the trials using perinatal MSCs are usually allogeneic transplantations. Interestingly, the MSCs obtained from different tissues have been used for the same indication but no solid evidence has been provided for showing which source or population of MSCs is clinically more effective. In fact, MSCs are heterogenous population of fibroblast-like multipotent cells, and their biological effectiveness varies with tissue source or subpopulation of MSCs [12,13]. The functional diversity of MSCs holds direct implications for basic research and therapeutic applications that require predictable function and efficacy. The use of homogeneous and effective populations of MSCs for therapeutic purpose remains an elusive goal. Thus, further clarify of biological features of MSCs will be helpful for selection of a suitable MSCs donor, tissues source or subpopulation, propagation procedures, cell doses, as well as clinical protocols for a given indication.
Materials and methods
Generation of human BMSCs, AMSCs, UMSCs and PMSCs
The study was approved by the Ethical Committee and the Institutional Review Board of the Chinese Academy of Medical Science & Peking Union Medical College. BM, adipose, placenta CV and UC samples were collected from healthy volunteers, and MSCs were isolated following the protocol described previously [11,14,15]. AMSCs, UMSCs, and PMSCs at passage 1 were supplied by the National Engineering Research Center of Cell Products (
BMSCs, AMSCs, UMSCs, and PMSCs were cultured in the same standard culture condition. The standard complete culture medium for MSCs was DMEM/F12 (DF12; Gibco, Grand Island, NY, USA), 10% fetal bovine serum (FBS; HyClone), 2 mM glutamine (Sigma, St. Louis, MO, USA), 100 U/ml penicillin-streptomycin (P/S; Invitrogen, Carlsbad, CA, USA), and 10 ng/ml epidermal growth factor (EGF; Peprotech).
Flow cytometric analysis
Phenotype of MSCs was analyzed using the following antibodies: FITC-conjugated-CD19, CD31 and CD34; PE-conjugated-CD11b, CD44, CD45, CD54, CD73, CD90, CD105, CD106 and HLA-DR. Non-specific isotype-matched antibodies served as controls. For Tregs, CD4+T cells were labeled with PE-conjugated CD4, FITC-conjugated CD25, and then labeled PE-Cy5-conjugated Foxp3 by using Foxp3 Staining Buffer Set (eBioscience). All of the antibodies were purchased from BD Pharmingen. Cells were analyzed by flow cytometry in a FACS Calibur, using the CellQuest software (Becton Dickinson).
Osteogenic and adipogenic differentiation
Osteogenic and adipogenic differentiation was carried out as described previously [14,15]. Briefly, cells were plated in 24-well plates at a density of 3000 cells/cm2. The medium was changed with specific induction medium 24 hours later. For osteogenic induction, STEMPRO Osteogenesis Differentiation Kit (GIBCO) was used. For adipogenic induction, medium consisted of DMEM supplemented with 10% FBS, 1 µmol/l dexamethasone, 5 µg/ml insulin, 0.5 mmol/l isobutylmethylxanthine (IBMX), and 60 µmol/l indomethacin was used. Reagents for adipogenic induction were purchased from Sigma. After 3 weeks of induction, the cells were stained using alizarin red S or oil red O solution.
Isolation of human peripheral blood mononuclear cells (hPBMCs) and human cord blood mononuclear cells (hCBMCs) CD4+T cells
Human PBMCs and CBMCs were isolated by Ficoll-Paque (Axis-Shield) density gradient centrifugation from blood of health volunteer donors. CD4+T cells were purified using relevant magnetic MicroBead kits (Miltenyi Biotec) according to the manufacturer’s instructions. The purity of CD4+T cells was more than 95%.
Co-culture of immune cells and MSCs
Cells were cultured in complete 1640 medium (Gibco) containing 10% fetal bovine serum (FBS) (FBS-NZ ORIGIN), 2 mM glutamine, 100 U/ml penicillin and streptomycin, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate. MSCs (30 Gy irradiated) were plated in 96-well flat-bottom plate and allowed to adhere for 18 h at 37°C, and then 105 PBMCs or CD4+T cells were added in. Human PBMCs were stimulated by phytohemagglutinin (PHA, Sigma, 10 µg/ml) and CD4+T cells were stimulated by PHA (10 µg/ml) and IL-2 (peprotech, 10 ng/ml) for 72 hours. For the regulatory T cells (Tregs), CD4+T cells were stimulated with IL-2 (10 ng/ml) for 5 days. Cells were isolated and labeled with CD4, CD25 and Foxp3.
Separation of CD106+CV-MSCs and CD106−CV-MSCs
Subpopulations of MSCs were separated using EasySep® PE Positive Selection Kit (StemCell Technologies). MSCs were detached by 0.025% trypsin and labeled with PE-anti CD106 antibody, and CD106+ and CD106− cells were separated according to manufacturer’s instructions. In some cases, flow sorting was used. MSCs were labeled with PE-anti CD106 antibody, the positive and negative cells were separated using BD Influx (Becton Dickinson). For both of the methods, the purity of isolated cells was more than 90%.
RNA isolation, reverse transcription and real time PCR
2 × 106 of CD106+ and CD106−CV-MSCs were collected and resuspended in Trizol (Invitrogen). The samples were stored in −80°C for genechip assay. Total RNA was extracted by E.Z.N.A. Total RNA Kit I (OMEGA), and cDNA synthesis with MLV RT kit (Invitrogen) for 50 min at 37°C in the presence of oligo-dT primer. Real-time polymerase chain reaction analyses were performed by an Applied Biosystems 7300 Real-Time PCR System. Gene expression was tested on different types of MSCs, and the fold-change in gene expression was calculated as
Primers for real time reverse transcription-polymerase chain reaction
Primers for real time reverse transcription-polymerase chain reaction
MSCs were plated at 4 × 104 cells/cm2 in a T25 flask overnight. After washing twice with PBS, cells were cultured with 10 ml EBM2 (Lonza) for another 2 days. Their conditioned media (CMs) were then collected, centrifuged at
In-vitro Matrigel tube formation assay
To investigate their angio-vasculogenic capacities, BMSCs, AMSCs, UMSCs, and PMSCs were collected and seeded directly on a Matrigel (BD Bioscience) precoated 96-well plate at 2 × 104 cells/well in MSC complete medium. Photographs were taken using the microscope (Olympus, Melville, NY, USA) after 12 hours of incubation (scale bar = 500 µm). Tube numbers in each well were counted and each sample was performed in triplicate (BMSCs,
To better study the paracrine action, the CMs of BMSCs, AMSCs, UMSCs and PMSCs were used to incubate endothelial cells and further assessed their trophic effects on the angiogenic function of endothelial cells. CMs supplemented with 2% FBS, EBM2 supplemented with 2% FBS (served as the negative control), and EGM2-MV (contained plentiful cytokines and 2% FBS, served as the positive control) were used to culture endothelial cells for 9 hours, respectively. Endothelial cells were seeded on Matrigel at 2 × 104 cells/well in a 96-well plate. Photographs were taken by microscope (scale bar = 500 µm). Tube numbers in each well were counted, and the total tube length and total tube area in each well were measured using ImageJ software (NIH, USA). Three donor derived CMs were used and each sample was performed in duplicate.
Endothelial cell proliferation assay
To investigate the proliferative effects of the secretion of BMSCs, AMSCs, UMSCs, and PMSCs, endothelial cells were incubated with different MSC population-derived CMs. The endothelial cell proliferation was measured using the Cell Counting Kit-8 (Dojindo, Rockville, MD, USA). Endothelial cells were first seeded at 1.0 × 104 cells/well in a 96-well plate overnight. After gently removing the medium, cells were washed twice with PBS, and then CMs supplemented with 2% FBS, EBM2 supplemented with 2% FBS (served as the negative control), and EGM2-MV (served as the positive control) were added to the cells for another 48 hours. Each sample was performed in triplicate. ΔOD450 indicated the final data after subtracting the background.
Enzyme-linked immunosorbent assay
To determine the concentration of VEGF, HGF, bFGF, and PGE2 in supernatants of MSCs, enzyme-linked immunosorbent assays (ELISAs) were performed. Cell-free supernatants were collected and kept in refrigerator at −80°C. The 48-hour supernatants of MSCs with an initial seeding density of 20,000 cells/well in a 96-well plate were collected for IFN-γ, VEGF, HGF, and bFGF measurements, while the 72-hour supernatants of MSCs with an initial density of 105 cells/well in a six-well plate were collected for PGE2 measurements. All of the supernatants were centrifuged at
Statistical analysis
Statistical analysis was performed by GraphPad Prism 6.0 software (Graph Pad Software, Inc., San Diego, CA, USA). All data are shown as the mean ± SEM. One-way ANOVA followed by Bonferroni multiple comparisons was employed to determine the statistical significance. Paired t test was used to analyze the endothelial gene modification after endothelial differentiation. The result was considered statistically significant if
Results
Phenotypic similarity and diversity of MSCs
The phenotypes of different types of MSCs were analyzed. Each type of MSCs was tested for 3 donors. A series of cell markers were examined at passage 3 of in vitro cultivation, including CD11b, CD19, CD29, CD31, CD34, CD44, CD45, CD54, CD73, CD90, CD105, CD106, HLA-DR, Nestin and SOX2. All the four types of MSCs highly expressed embryonic stem cell markers Nestin and Sox2, as well as CD29, CD44, CD73, CD90 and CD105. The most significant difference of their phenotypes was the CD106, which highly expressed on the PMSCs (68%), moderately on BMSCs (13%), lightly on UMSCs (4%) and absent on AMSCs (Table 2).
FACS analysis of phenotype of BMSCs, AMSCs, UMSCs and PMSCs. Data were presented as Mean ± S.E.M.
FACS analysis of phenotype of BMSCs, AMSCs, UMSCs and PMSCs. Data were presented as Mean ± S.E.M.
Typical MSC markers including CD105, CD90, CD73, CD29 and CD166. Immunogenic markers: HLA-ABC and HLA-DR. Hematopoietic and endothelial cells related markers: CD45, CD144, CD133 and CD31. Mesenchymal/Endothelial related marker: CD106. The data was presented as the mean percentage of positive cells mean ± SEM (
To determine most significant differences among MSCs of different origins, we first compared the proliferative ability of MSCs isolated from human term placental CV, UC, adult BM and adipose using identical methods. The MSCs isolated from 22 UC samples were cultured using identical methods and propagated until proliferative arrest. It was found that the passage number of UMSCs varied among individual donors, from 23 to 39 passages, with a mean of 30.68 passages (Fig. 1(A)), whereas adult BMSCs and AMSCs had only a mean 7 and 14 passage number, respectively (
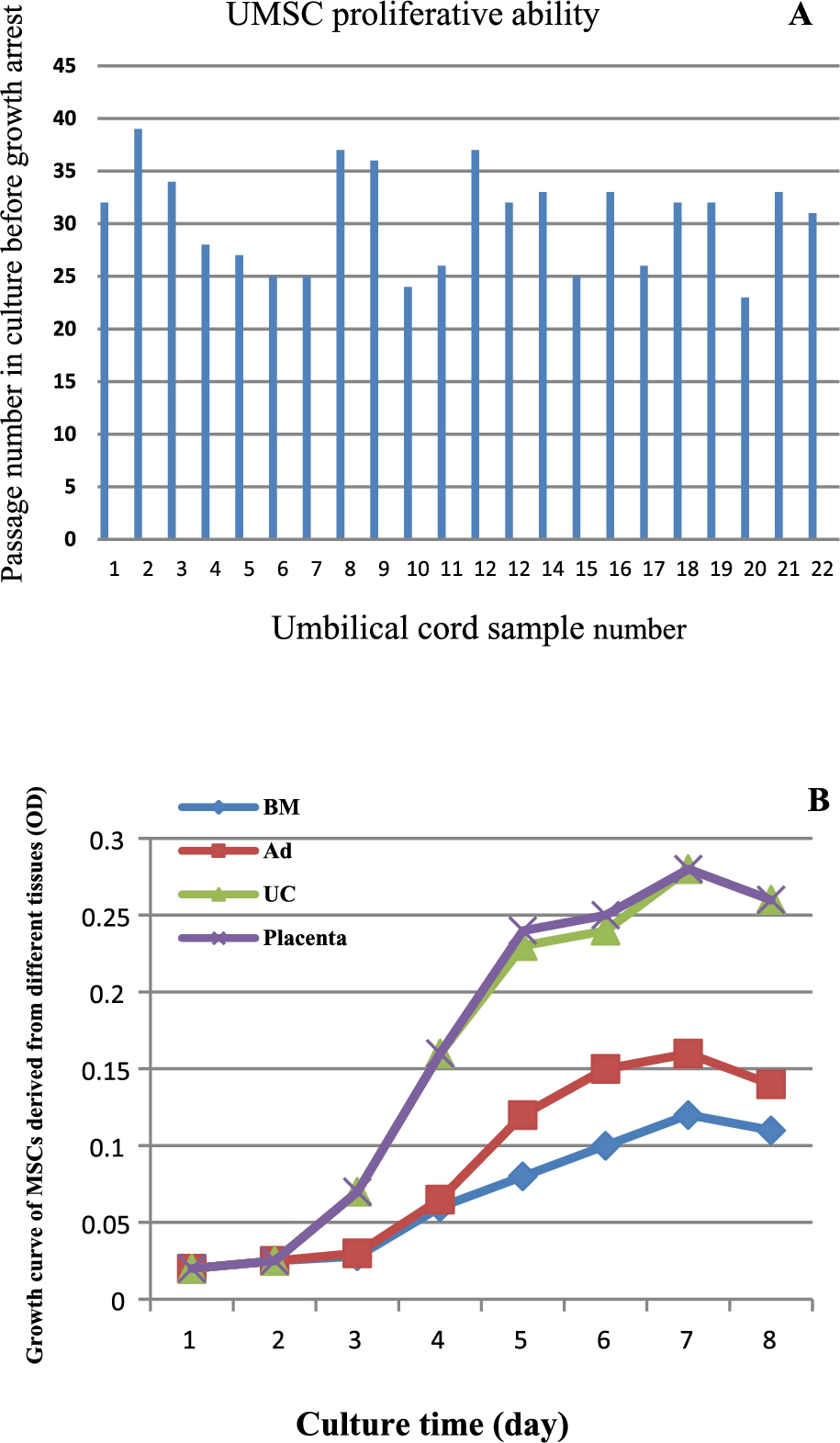
Comparison of proliferation ability of mesenchymal stem cells (MSCs) derived from umbilical cord (UC), bone marrow (BM), adipose (Ad) and placenta chorionic villi. (A) the passage number of MSCs derived from individual umbilical cords during long-term culture. (B) the growth curves of BMSCs and AMSCs derived from three pairs of the same adult donors and UMSCs and PMSCs derived from three pairs of the same baby donors.
MSCs isolated from different tissues were cultured in different specific induction medium for three weeks. All types of MSCs displayed their ability to differentiate into adipogenic, osteogenic and chondrogenic lineages. However, BMSCs and AMSCs had higher adipogenic differentiation ability than perinatal tissue MSCs. In addition, BMSCs seems the best among four sources of MSCs in the promotion of osteogenic and chondrogenic differentiation (Fig. 2).
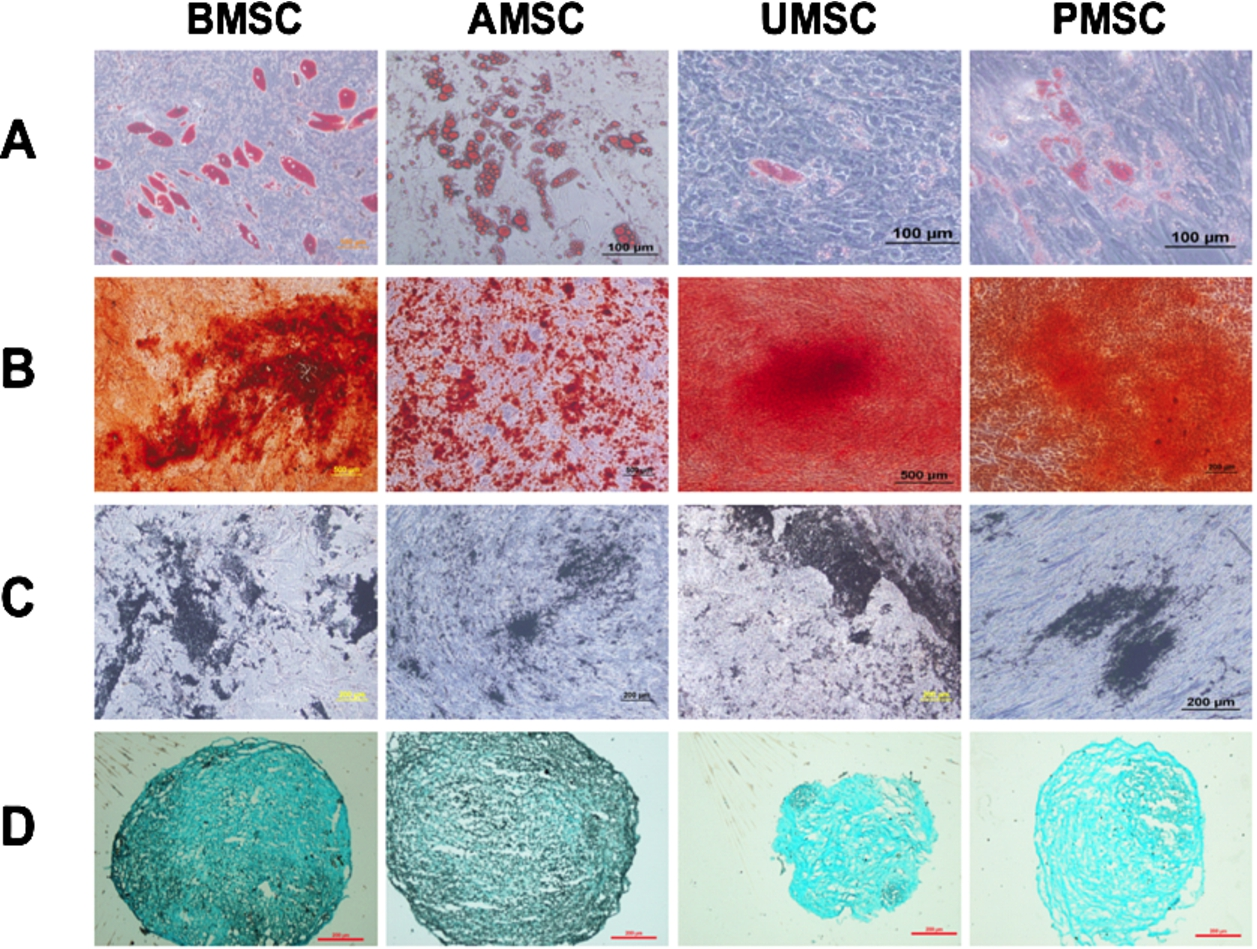
Multi-differentiation of BMSC, AMSC, UMSC and PMSC. MSCs cultured in different specific induction medium for three weeks displayed their adipogenic, osteogenic and chondrogenic commitment. (A) Adipogenic differentiation was verified by Oil Red O staining, scale bar = 100 µm. Osteogenic transformation was confirmed by staining with Alizarin Red (B) and von Kossa (C), scale bar = 200 µm or 500 µm. (D) Alcian Blue staining was performed to identify the sulfated glycosaminoglycan matrix deposition in chondrocytes trans-differentiated from BMSC, AMSC, UMSC and PMSC, scale bar = 200 µm.
To determine their angio-vasculogenic abilities, BMSCs, AMSCs, UMSCs, and PMSCs were directly seeded on Matrigel and the tube formation was observed after 12 hours of incubation. Interestingly, intact tube structures were seen in the BMSC and PMSC groups rather than in the groups of AMSCs and UMSCs (Fig. 3(A)). Tube numbers in the BMSCs and PMSCs groups were 11.65 ± 2.92 and 6.49 ± 1.18, respectively, much higher than those in the AMSC and UMSC groups (0.91 ± 0.76 and 0.41 ± 0.20,

MSCs derived from different tissue origins display distinct tube formation ability on matrigel in vitro. MSCs were seeded on matrigel in vitro at 2 × 104 cells/well. MSCs could form tube-like structures on matrigel temporarily. (A) Representative photos were taken at 12 hours, scale bar = 500 µm. (B) The numbers of tube structures were counted and analyzed. BMSC
To determine their paracrine actions, the condition media (CM) of MSCs was analyzed for testing their stimulating effects on the proliferation and tube formation capacity of endothelial cells. It was interesting to observe that BMSCCM and PMSCCM could significantly promote endothelial cell proliferation (1.23 ± 0.06 and 1.24 ± 0.06) in contrast to AMSCCM and UMSCCM (0.84 ± 0.02 and 0.85 ± 0.07;
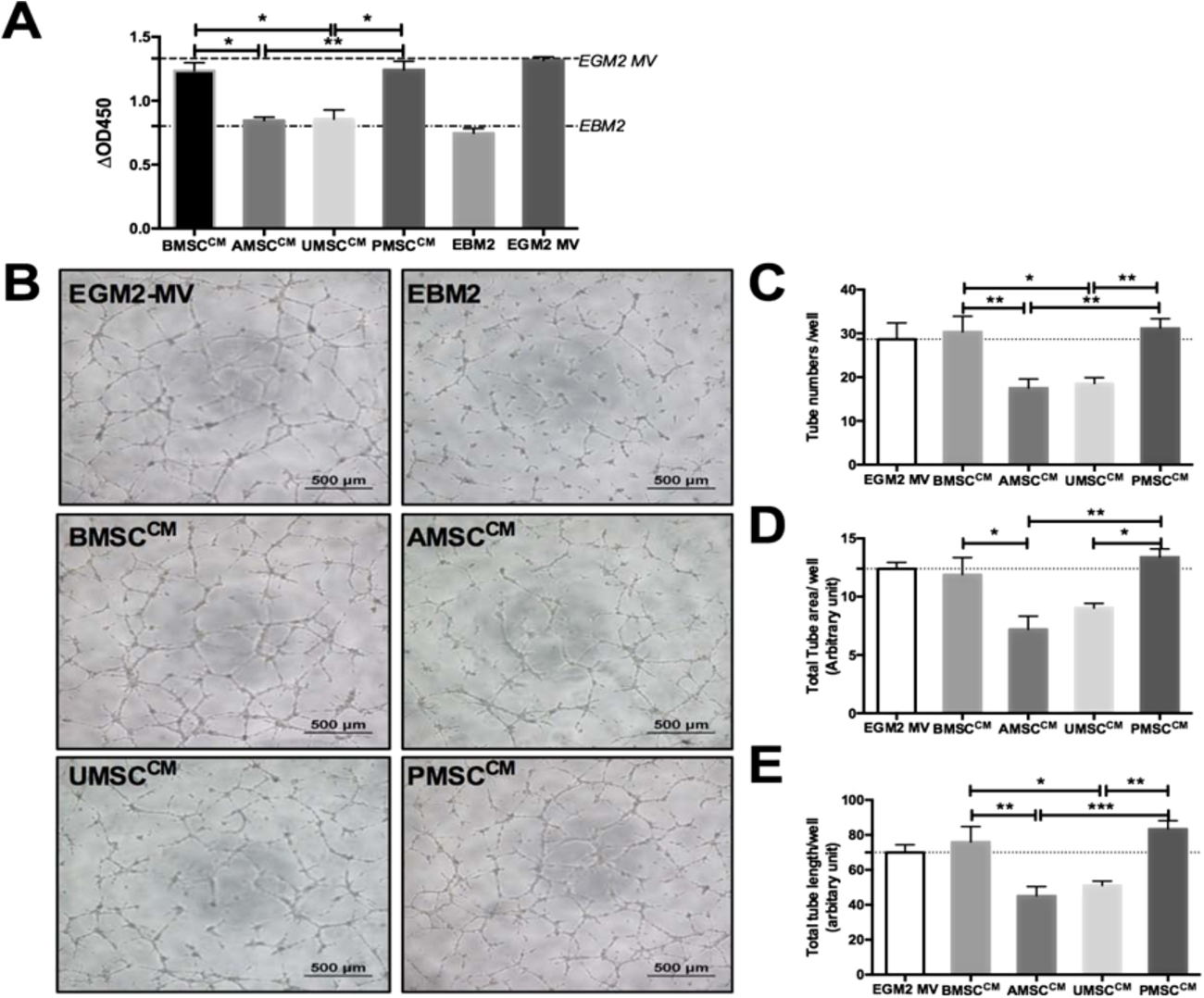
Conditioned mediums (CMs) of BMSC, AMSC, UMSC and PMSC exert various pro-angiogenic effects on endothelial cells in vitro. CMs were generated on 106 cells after 48 hours incubation with EBM2. CMs supplemented with 2% FBS, EBM2 with 2% FBS and EGM2-MV were used to comparatively analyze the paracrine activities of MSCs. The latter two were severed as negative and positive controls, respectively. (A) Similar like the EGM2-MVgroup, BMSCCM and PMSCCM significantly promote endothelial cells proliferation with comparison to EBM2 during 48 hours culture; however, AMSCCM and UMSCCM did not produce such a promotion effect. Each sample was performed in triplicates (
To determine if different sources of MSCs affect Th1 specific cytokine IFN-γ expression, we then compared levels of IFN-γ using ELISA. PBMCs secreted IFN-γ upon stimulation of PHA (10 mg/ml). All of these MSCs suppressed the expression of IFN-γ in hPBMCs in the presence of PHA (Fig. 5(A)). To exclude the possibility that the differences among MSCs were due to individual differences, we tested the UC-MSCs and CV-MSCs obtained from the same fetus origin. Interestingly, CV-MSCs showed a significantly increased inhibitory effect on IFN-γ production of the hPBMCs compared to UC-MSCs (Fig. 5(B)). Figure 5(C) shows that the concentration of PGE2 in the culture medium of placental CV-MSCs is significantly increased in comparison with other sources of MSCs.
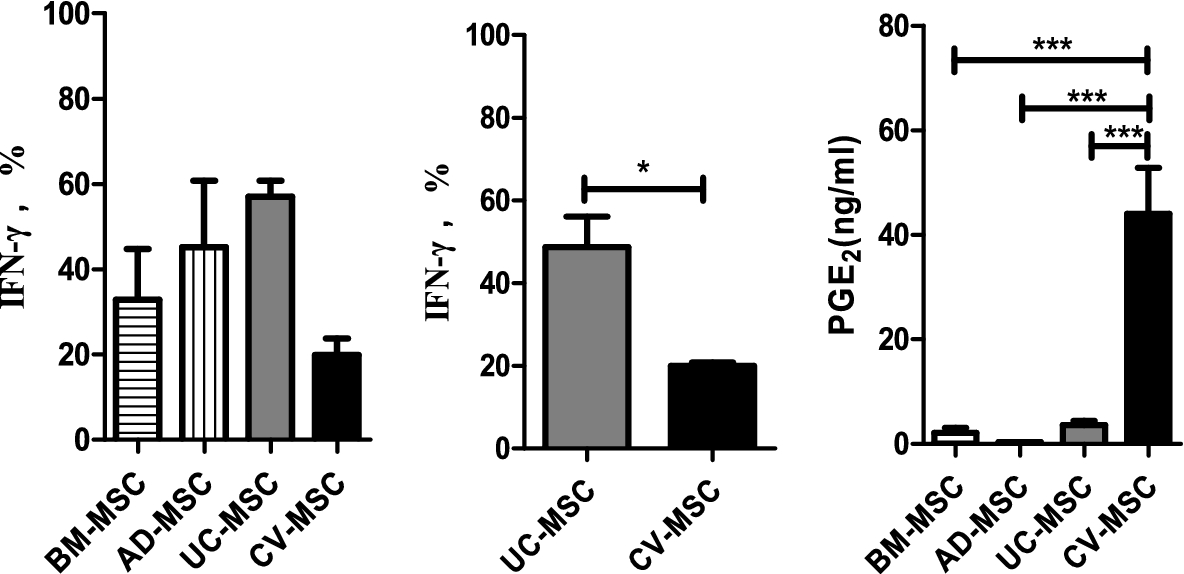
Comparison of immunomodulation ability in BM-MSC, AD-MSC, UC-MSC and CV-MSC. (A) BM-MSC, AD-MSC, UC-MSC and CV-MSC were cocultured with PHA (10 µg/ml) stimulated PBMC for 72 hours. 105 PBMC and
It was interesting to note that CV-MSCs expressed highest level of mRNA for COX-2 (Cyclooxygenase-2, inducible synthase for PGE2), IL-1a, IL-1b, IL-6 and IL-8. However, the expression of TGF-b1 was similar in each type of MSCs. The expression of IDO1 mRNA was very low in all sources unless treated by pro-inflammatory cytokines like IFN-γ. Importantly, CV-MSCs showed strongest ability to secrete PGE2 among the four sources of MSCs (44.0 ng/ml) (Fig. 6).
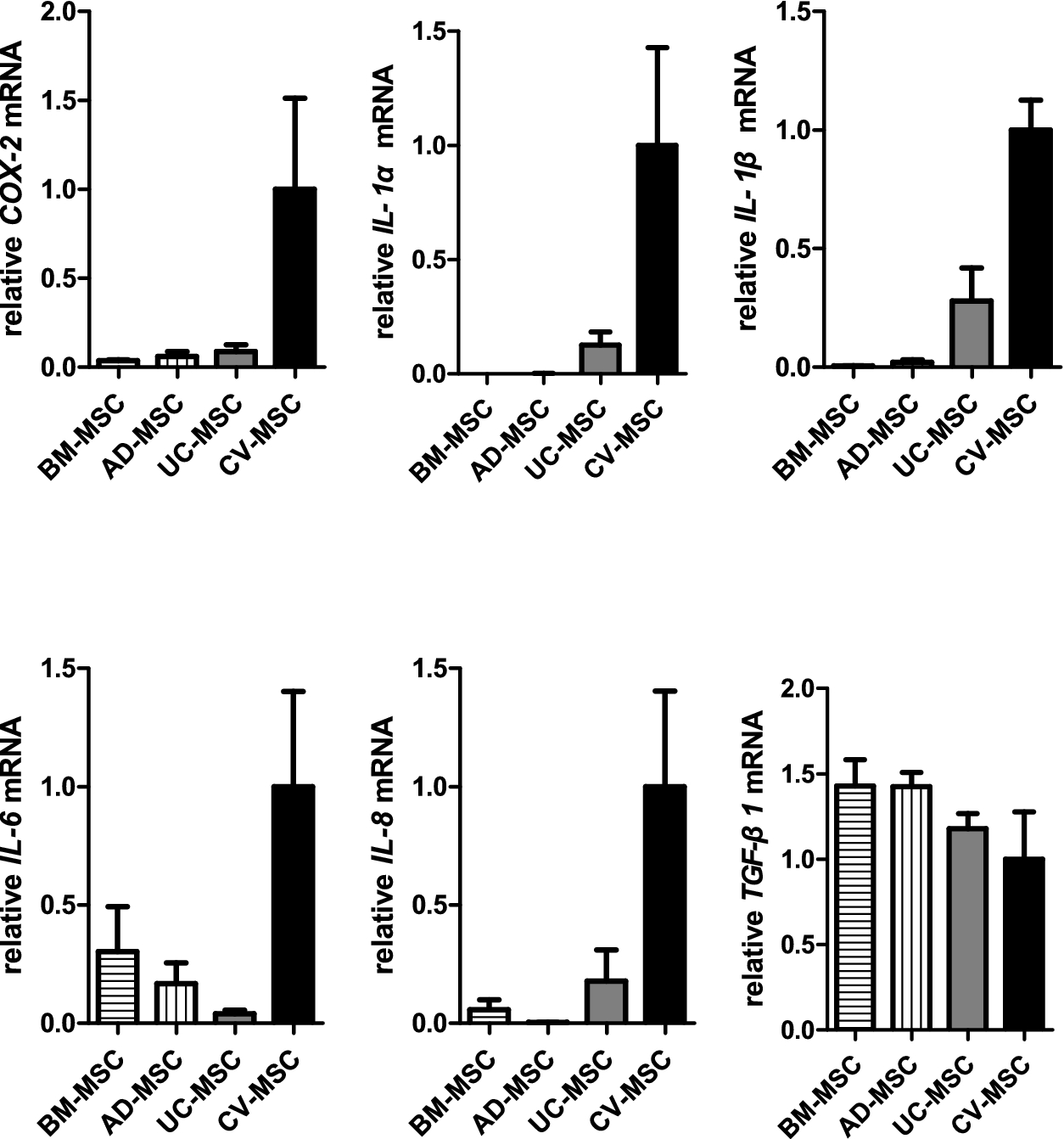
Relative mRNA expression of COX-2, IL-1α,IL-1β, IL-6, IL-8 and TGF-
PMSCs express highly CD106 (VCAM-1). We thus hypothesized that CD106+PMSCs may represent a subpopulation with increased immunosuppressive and pro-angiogenic activities. To test this hypothesis, the CD106+ cells and CD106− cells were therefore separated and co-cultured with cord blood CD4+T cells at a ratio of 1:10 for 72 hours stimulated with PHA (10 µg/ml) and IL2 (10 ng/ml). IFN-γ (A) and TNF-α (B) production in the supernatant were measured by ELISA. Percentage of IFN-γ and TNF-α was gained by direct comparison to the control (activated cord blood CD4+T cells alone). CD106+CV-MSCs or CD106−CV-MSCs did not induce the secretion of IFN-γ and TNF-α of cord blood CD4+T cells without stimulation but their addition resulted in a significant reduction of IFN-γ and TNF-α secretion from cord blood CD4+T cells stimulated with PHA. However, their inhibitory degree was different. The CD106+CV-MSCs induced an almost complete inhibition of IFN-γ secretion whereas CD106−CV-MSCs induced only partial inhibition (Fig. 7).
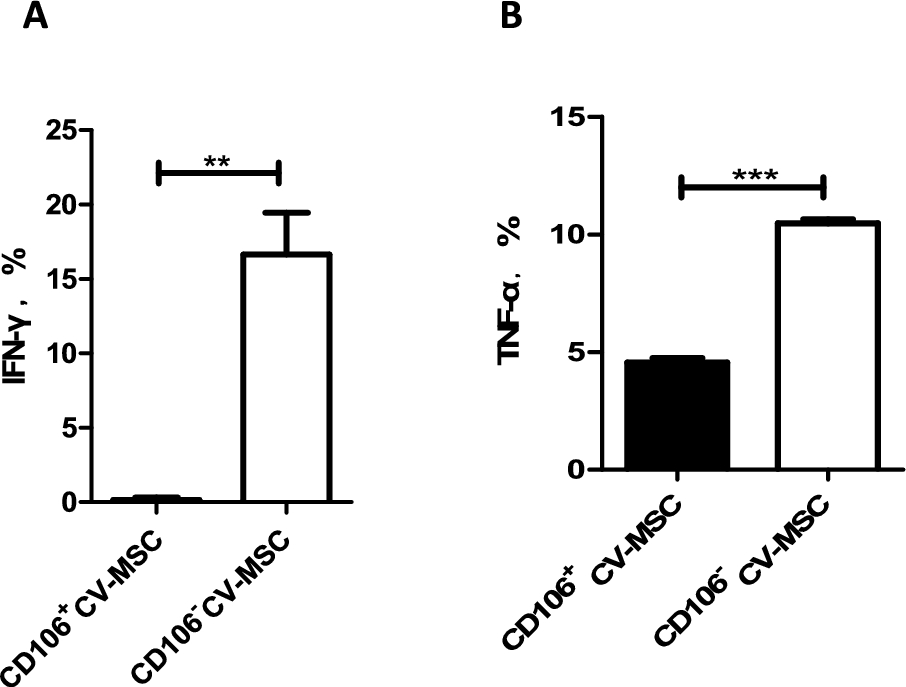
CD106+ or CD106− CV-MSCs were co-cultured with cord blood CD4+T cells at a ratio of 1:10 for 72 hours stimulated with PHA (10 µg/ml) and IL2 (10 ng/ml). IFN-γ (A) and TNF-α (B) production in the supernatant were measured by ELISA. Percentage of IFN-γ and TNF-α was gained by direct comparison to the control (activated cord blood CD4+T cells alone), each performed in at least triplicate (
We then compared the pro-angiogenic ability CD106+CV-MSCs with CD106−CV-MSCs in vitro and in vivo. To our surprise, without exogenous VEGF, CD106+CV-MSCs spontaneously formed about 4.14-fold intact tubular structures on Matrigel compared with CD106−CV-MSCs (
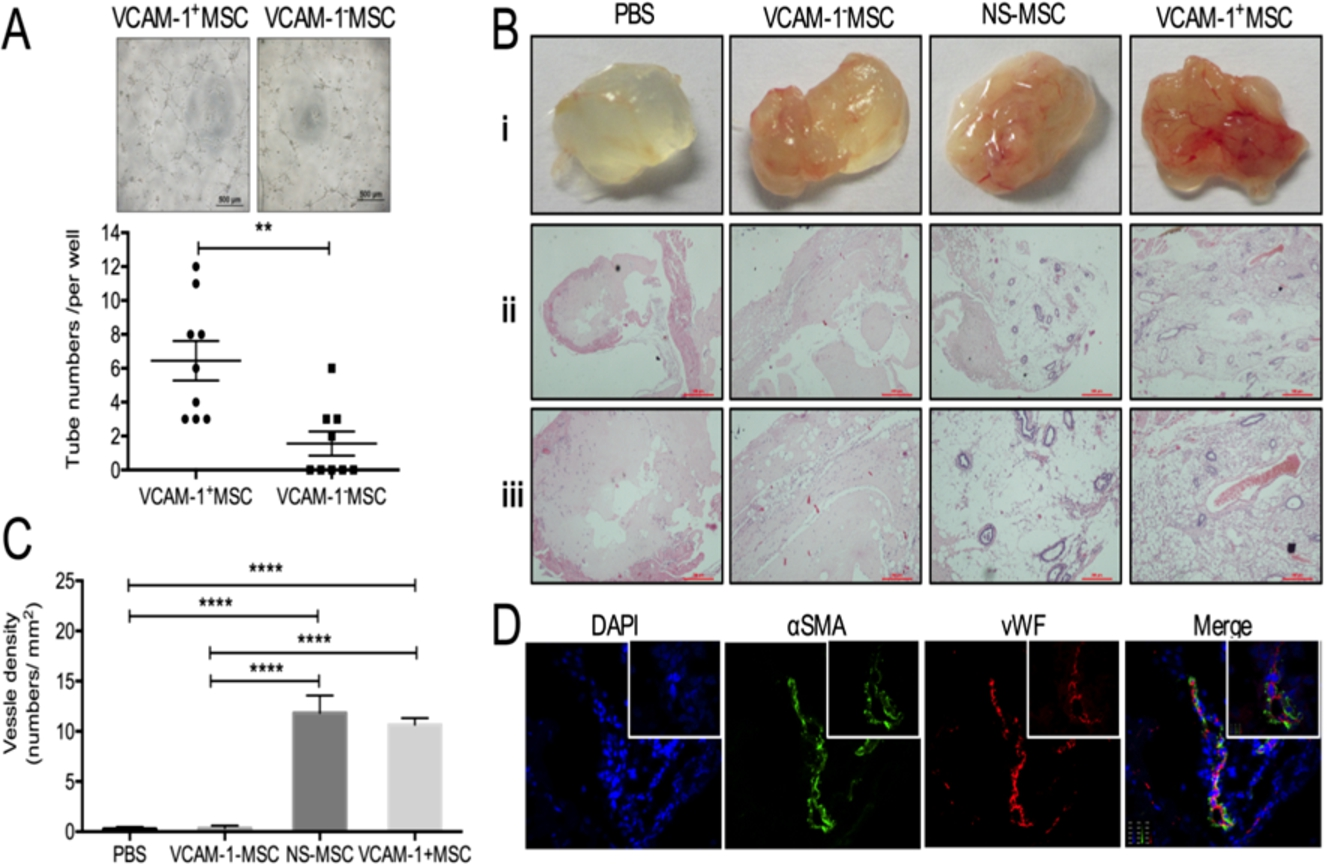
CD106(VCAM-1)+CV-MSCs revealed vasculo-angiogenic potential by angiogenesis analysis with Matrigel in vitro and vivo. (A) CD106+CV-MSCs spontaneously formed much more intact tube-structures on Matrigel than CD106− CV-MSCs (
To explore the mechanism underlying increased immunosuppressive and pro-angiogenic activities of CD106+CV-MSCs, we performed microarray to analyze the gene expression pattern of CD106+CV-MSCs and CD106−CV-MSCs, and real-time PCR was performed to confirm the results from microarray analysis. The expression of many adherence molecules and cytokines were different in CD106+ and CD106−CV-MSCs. CD106+CV-MSCs express higher levels of IL6, IL8, HGF, ANG, MMP2, VEGF-A, TGFβ, and bFGF compared with CD106−CV-MSCs (Fig. 9(A) and (B)), indicating different functions for two kinds of MSCs. Notably, the gene expression profiles associated with immunoregulatory factors were different, indicating different functions of these two subpopulations. CD106+CV-MSCs showed an increased ability to secrete PGE2 compared to CD106−CV-MSCs (Fig. 9(C)).
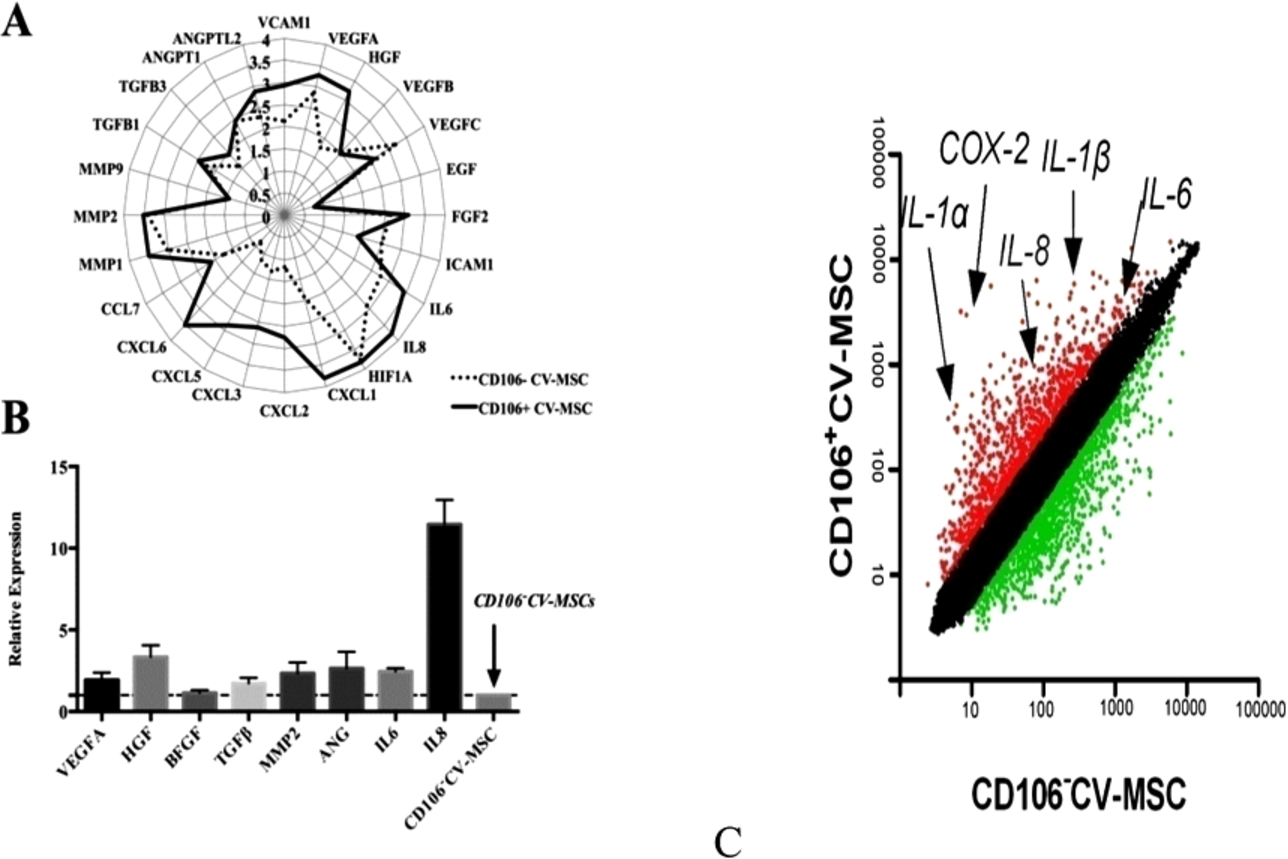
Immunomodulatory and angiogenic genes were up-regulated in CD106+CV-MSCs. (A) The gene expression profile of CD106+ and CD106−CV-MSCs were determined using Affymetrix oligoarray and the angiogenic genes were valued and expressed in
To test the ability for another important immune modulator IDO1, IFN-γ was used to stimulate the MSCs. IFN-γ significantly up regulated the expression of IDO1 in 24 hours. CD106+ cells express higher level of IDO1 than CD106− cells in the presence of IFN-γ (Fig. 9(D)). Together, CD106+CV-MSCs expressed higher level of immunomodulatory factors compared to CD106−CV-MSCs.
Mesenchymal stem cells (MSCs) have being widely evaluated their effectiveness in treatment of many diseases. However, MSCs can be isolated from many tissues and their heterogeneity and functional variability become the bottleneck for standardization of therapeutic protocols. A recurrent question is what source or population of MSCs is suitable for specific clinical indications.
In the present study, we performed a parallel comparison of biological features and therapeutic potentials of four types of MSCs derived from adult tissues (bone marrow and adipose tissue) and perinatal tissues (umbilical cord and placental chorionic villi). We demonstrate that MSCs are heterogeneous in individual donor, tissue origin, culture condition. First, we observed that the cell proliferation and differentiation abilities of MSCs are variable. The UMSCs isolated from 22 umbilical cord samples showed variable proliferative ability using identical methods. In addition, MSCs isolated from different tissues of the same adult donors also exhibited the variation in their proliferative ability. In comparison with adult MSCs, the perinatal tissue derived MSCs possess greater proliferative capacity. Furthermore, we observed that MSCs isolated from different sources showed some variability of multi-lineage differentiation. Perinatal MSCs seem less prone to adipogenic and osteogenic differentiation compared to adult AMSCs and BMSCs.
Variation in MSCs has been noted not only for proliferative and differentiation abilities among donors and within the tissue origins isolated from the same donor, but also for their unique functions in term of immunomodulation and pro-angiogenesis. All sources of MSCs have been found to suppress the expression and secretion of IFN-γ of hPBMCs stimulated by PHA. To exclude the possibility that the differences among MSCs were due to individual differences, we tested the UMSCs and CV-MSCs obtained from the same fetus origin. Interestingly, PMSCs showed a significantly increased inhibitory effect on IFN-γ production of the hPBMCs compared to UC-MSCs. Previous study has shown that MSCs exert immunosuppressive activities through a PGE2-dependent mechanism [16]. Here, we found that the concentration of PGE2 in the culture medium of placental CV-MSCs was significantly increased in comparison with other sources of MSCs, indicating that PMSCs contain a valuable population of MSCs with increased immunosuppressive activity.
We then investigate the angio-vasculogenic abilities of BMSCs, AMSCs, UMSCs, and PMSCs and noted that MSCs regardless of their tissue origins could develop similar endothelial-relevant functions in vitro in spite of their feeble expression of endothelial-related genes and proteins. Additionally, we surprisingly found that BMSCs and PMSCs could directly form tubular structures on Matrigel and their conditioned medium showed significant pro-angiogenic activities on endothelial cells in vitro compared with those of AMSCs and UMSCs.
To explore the precise mechanism through which MSCs exhibit their functional heterogeneity, we had searched for the key biomarker capable of predicting the function and efficacy of MSCs in term of therapeutic potentials. We were luck to identify CD106 (VCAM-1) as a valuable cell surface marker and to separate a subpopulation of CD106+MSCs from heterogenous MSC population. Our results showed a positive correlation between CD106 positivity and functional activities of MSCs. PMSCs highly express CD106 and also possess strongest immunosuppressive and pro-angiogenic activities among all four sources of MSCs. Comparative studies further showed that CD106+MSCs differed from CD106−MSCs in their immunosuppressive and pro-angiogenic activities.
To determine the mechanism underlying increased immunosuppressive and pro-angiogenic functions of CD106+CV-MSCs, we have compared the secretion of angiogenic an immunomodulatory factors and the gene expression pattern of CD106+CV-MSCs with CD106−CV-MSCs. We provide several lines of evidence that increased immunosuppressive and angiogenic activities of CD106+MSCs are highly related to their ability to secrete high levels of immunoregulatory and pro-angiogenic cytokines and factors.
CD106, also known as vascular cell adhesion molecule 1 (VCAM-1), is a cytokine-inducible cell surface protein capable of mediating adhesion to leukocytes expressing alpha 4 integrins, and is extensively expressed on endothelial cells [17], and also constitutively expressed on some stromal cells, existing in a particular vascular niche [18]. CD106 plays a critical role in early embryonic development since CD106-deficient mice often die early or show multiple severe defects in placental development [19]. Recent studies demonstrated that CD106 overexpression was associated with tumor angiogenesis [20–22]. We have previously reported our partial data on the roles of placental CV derived CD106+MSCs in immunomodulation [23] and pro-angiogenesis [24].
In summary, this comparative study provides solid evidence showing the heterogeneity and functional variability of MSCs, and indicates that the CD106 as valuable surface marker and potency predictor to select a unique subpopulation of MSCs with superior immunosuppressive and pro-angiogenic activities. Future work will be best directed toward on standardizing methods which allow the validation of MSC products with defined predictive of potency in suitable pre-clinical models and clinical settings.
Footnotes
Acknowledgements
This study was supported by the Natural Science Foundation of China (81330015 and 31470951) and the National Science and Technology Support Program (2013BAI01B09).
Conflict of interest
The authors have no conflict of interest to report.