
Abstract
Background:
Mesenchymal stem cells (MSC) have been shown to have potent immunoregulatory effects. They are able to mitigate inflammation in many contexts of immune disorders, including autoimmune diseases and graft-versus-host disease (GVHD). Endotoxemia can induce systematic inflammation in the body. In this study, we try to investigate whether MSC can attenuate inflammation in models of LPS-induced endotoxemia.
Methods:
Bone marrow MSC (BMSC) were isolated and expanded from rats of 4∼6-week age. Adult mice were divided randomly into Control group, Model group and BMSC group. LPS were injected peritoneally into mice of Model group and BMSC group to induce endotoxemia. For BMSC group mice,
Results:
Inflammatory pathological changes in lung and liver in BMSC group were comparable to those in Model group. Moreover, in some animals, the injuries were exacerbated after BMSC treatment. Accordingly, wet/dry ratio of lung in BMSC group mice was higher than that in Model group mice. IL-1β level in BMSC-treated group mice was significantly augmented, however, no significant changes were detected between these two groups for level of TNF-α.
Conclusion:
Our results showed that application of BMSC in LPS-induced endotoxemia models couldn’t attenuate the inflammatory injuries in tissues. Although BMSC have been shown to be able to induce immune inhibition, however, in some instances, their immuno-inhibitory function might be regulated by the local environment.
Introduction
Endotoxemia is a severe clinical syndrome caused by infection, severe trauma, burns, etc, which may result in septic shock and multiple organ dysfunctions. Uncontrollable inflammatory response mediated by activated immune cells and inflammatory factors is the main pathophysiological mechanism. Macrophages and neutrophils play a crucial role in the early stage of endotoxemia. Endotoxin or lipopolysaccharide (LPS) activates intracellular signal transduction system on neutrophils, monocytes/macrophages, prompting transcription and expression of inflammatory cytokines (such as TNF-α, IL6, IL-1β), which induce further activation of more inflammatory cells including T cells, and release of more inflammatory factors. Inflammatory factors induce injuries to different organs and tissues through induction of cell apoptosis, increasing permeability of microvascular endothelium [1]. Therefore, it is very important to block the activation of inflammatory cells and secretion of inflammatory factors to treat endotoxemia in clinic.
Mesenchymal stem cells (MSC) are multipotent precursors of nonhematopoietic mesodermal lineages, which are capable of clonal differentiation into different lineages, such as osteoblastes, chondrocytes, adipocytes, and hepatocytes [2]. They can also support hematopoiesis through secretion of a series of cytokines. Moreover, MSC have been shown to be able to inhibit effectively immune response mediated by T cells, B cells, NK cells, antigen presenting cells (APC), etc. Their immunosuppressive function is realized through direct contact with immune cells mediated by Notch signaling, HLA-G, etc, or through secretion of a series of immunoregulatory factors, as IL-10, HGF, IL-6, PGE2, indoleamine 2, 3-dioxygenase (IDO) [3]. Owing to their immunoregulatory ability, they have been shown to exhibit positive outcomes in different contexts of immune disorders, including autoimmune diseases as collagen-induced arthritis [4], type 1 diabetes [5], experimental autoimmune encephalomyelitis [6], allogenic and xenogenic transplantation [7]. Experimental studies have also provided evidence that MSC may be beneficial in some inflammatory diseases, as inflammatory dilative cardiomyopathy [7] and acute lung injury [8]. Since that tissue injuries and organ dysfunctions are mainly elicited by inflammation in endotoxemia, in this study, we try to investigate whether MSC can ameliorate inflammatory injuries in LPS-induced endotoxemic animal models.
Materials and methods
Animals
Healthy 4∼6-week old Sprague-Dawley (SD) rats and 10∼12-week old male Kunming mice were purchased from the Experimental Animal Center of Wuhan University. All experimental animal procedures were approved by the Local Ethical Committee on Animal Care and Use of Wuhan University.
Isolation and expansion of bone marrow MSC (BMSC)
BMSC were isolated and expanded from 4∼6-week-old SD rats. Briefly, rats were anethetized with 7% chloral hydrate solution (350 mg/kg body weight) through intraperitoneal injection. Femoras and tibias were removed under aseptic conditions, bone marrow cells were flushed out with phosphate-balanced saline (PBS) by using a syringe. Collected cells were centrifuged at 1000 rpm for 5 minutes. Cell pellet was resuspended with Minimum Essential Medium alpha (MEM-α, Hyclone, USA) containing 10% (v/v) fetal bovine serum (FBS; Gibco, USA), 100 U/mL penicillin and 100 μg/mL streptomycin (Sigma-Aldrich, USA). Cells were then seeded into 75 cm2 culture flasks at a density
Cytometry analysis
Phenotype of expanded BMSC were evaluated by the expression of CD34, CD45, CD29 and CD44 molecules. Cells were incubated at 4°C for 30 minutes with primary FITC (fluoresceine isothiocyanate)-conjugated antibodies (Biolegend, San Diego, USA). Results were analyzed by FACS (Becton Dickinson Biosciences, USA). For cell apoptosis analysis, endothelial cells (ECV304 cells, China Center for Type Culture Collection, China) were stained with annexin-V/PI according to the manufacturer’s instructions (Keygene biotech, China), and results were analyzed by FACS.
Osteogenic differentiation
Cells were seeded in a 6-well plate at density of 5000 cells/cm2 and cultured in α-MEM supplemented with 10% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin up to confluence. Osteoblastic differentiation of BMSC was induced by incubation with low glucose DMEM (Hyclone, USA) supplemented with 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 10 mmol/L β-glycerophosphate, 0.1 μmol/L dexamethasone, 50 mg/L ascorbate-2-phosphate (Sigma-Aldrich, USA). Medium were changed every 3 days, after 21 days of induction, wells were fixed with 4% paraformaldehyde, and stained with Alizarin red (Sigma-Aldrich, USA).
Adipogenic differentiation
Cells were seeded in a 6-well plate at density of 5000 cells/cm2, 100 U/mL penicillin and 100 μg/mL streptomycin up to confluence. Adipogenic differentiation of BMSC was induced by incubation with DMEM (Hyclone, USA) supplemented with 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, 10 μg/mL insulin, 200 μM indomethacin, 50 μg/mL 3-isobutyl-1-methylxanthine (Sigma-Aldrich, USA) and 10 μM dexamethasone. Medium was changed every 3 days, after 21 days of induction, cells were fixed with 4% paraformaldehyde and stained with Oil red O (Sigma-Aldrich, USA).
Induction of endotoxemic animal model
Adult male Kunming mice were randomly divided into 3 groups: Control group, Model group and BMSC group. In Model group and BMSC group, 10 mg/kg lipopolysaccharide (LPS; Sigma-Aldrich, USA) was injected peritoneally to establish the model of endotoxemia. The same volume of PBS was injected in Control group.
Detection of lung wet-to-dry weight ratio
Left side lungs were collected and weighted (wet weight), then they were baked for 24 h at 60°C and weighted (dry weight). Wet-to-dry weight ratio were calculated.
Histochemistry detection of pathological changes in tissues
Parts of lungs and livers were rapidly removed from animals upon sacrifice, fixed in 4% phosphate-buffered formaldehyde for 24 hours. They were then dehydrated and embedded in paraffin and sectioned into 4-μm-thick slices. Hematoxylin-eosin staining was then performed. Pathological changes of tissues were analyzed under microscope.
Enzyme-linked immunosorbent assay (ELISA)
For cytokine determination in lung tissues, right lungs of animals were collected, weighted and stored on ice. Then tissues were homogenized in cold PBS (tissue weight:volume of PBS = 1:10). Homogenized mixture was centrifuged at 12,000 rpm for 20 minutes at 4°C and supernatant was collected. Cytokine levels of IL-1β and TNF-α were measured in triplicate with ELISA kits (ebiosciences, USA) according to the manufacturer’s instructions.
Isolation and culture of macrophage
Before collecting lung and liver tissues from animals, peritoneal macrophages were harvested by washing the peritoneal cavity with 5 mL cold sterile saline as described early [9]. Collected cells were centrifuged at 1000 rpm for 8 minutes. Cell pellets were then suspended in complete cell culture medium of PRMI 1640 (Hyclone, USA) supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin at density of
Statistical analysis
Results were presented as mean ± SEM. The data were analyzed using analysis of variance (ANONA) and Student’s t-test. A P value of less than 0.05 was considered to be statistically significant.
Results
Phenotype and differentiation ability of BMSC
FACS results showed that BMSC of 2∼4 passage expressed high levels of mesenchymal cell markers CD29 and CD44, whereas expression of hematopoietic markers CD34 and CD45 was very low or absent (Fig. 1(A)). BMSC could differentiate into both osteoblasts and adipocytes, as shown by positive staining of Alizarin red and Oil red O (Fig. 1(B)).

Identification of BMSC. BMSC of passage 2∼4 were identified through both phenotype and their differentiation ability. BMSC were stained with primary FITC-conjugated anti-CD34, anti-CD45, anti-CD29 and anti-CD44, results were analyzed by FACS (A). Cells were induced to osteogenic and adipogenic differentiation and then stained with Alizarin red and Oil red (B).
Inflammation can cause damage to the vasculature of the lung, and induce fluid extrusion into the air spaces or parenchyma of the lungs, which results in increased wet lung weight. Our results showed that wet-to-dry lung weight in Model group and BMSC group was significantly higher than that in Control group (

Wet-to-dry weight ratio of lung tissues. Wet and dry weight of lungs were weighted, wet-to-dry weight ratios were calculated. (∗,
As shown in Fig. 3(A), mice in Model group showed marked thickened alveolar wall, decreased alveolar airspaces, and infiltration of inflammatory cells in the interstice of the lung tissues. In liver tissues, dilation and hyperemia of sinusoidal, inflammatory infiltration, swelling and necrosis of heptocytes were found. In BMSC group, no significant amelioration of pathological changes were found in both lung and liver tissues (Fig. 3). Furthermore, even severe pathological changes were found in some animals in BMSC group than in Model group.
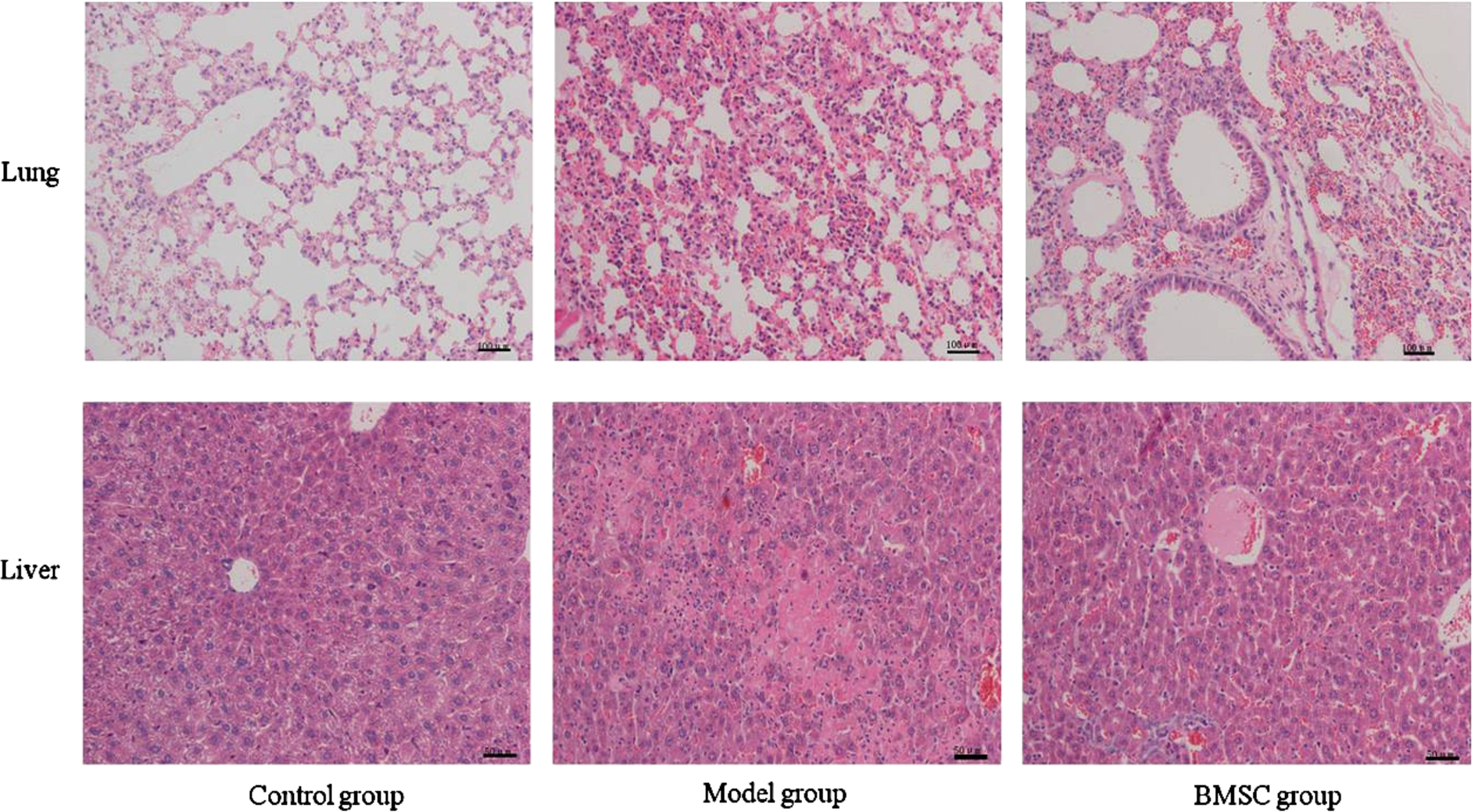
Hematoxylin-eosin staining of lung and liver tissues. BMSC was infused one hour after LPS injection. Lung and liver tissues were collected from animals at the end of twenty-four hours and subjected to pathological evaluation through Hematoxylin-eosin staining. Representative micrographs of lung and liver histology in animals of each group were shown here. The number of animals studied per experimental group was as follows:
ELISA results (Fig. 4) showed that levels of inflammatory cytokines in lung tissue homogenate from Model group were significantly higher than those in Control group, including TNF-α and IL-1β. Level of IL-1β in BMSC group was significantly increased compared with that in Model group (
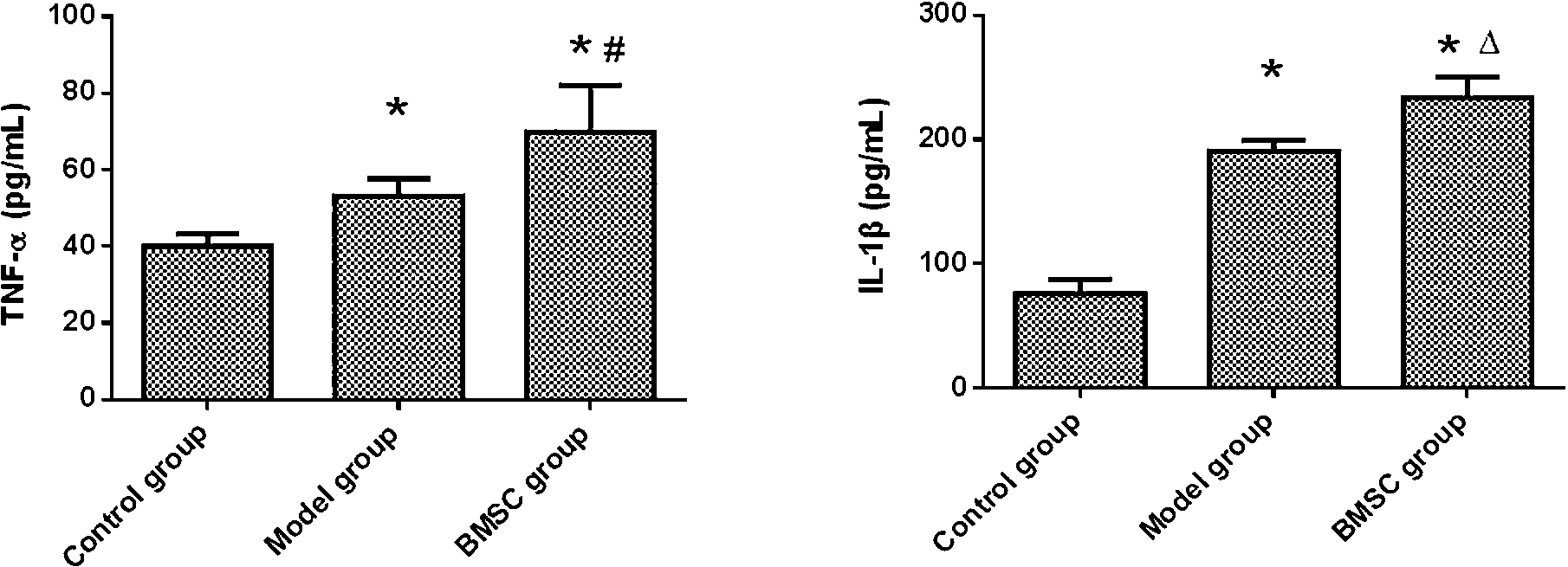
Levels of TNF-α and IL-1β in lung tissues. Parts of lungs were collected and homogenized. Expression of TNF-α and IL-1β in lung tissues were detected through ELISA. (∗,
LPS-activated macrophages can release a large number of inflammatory molecules, which can further induce cell injuries. We isolated abdomen macrophages from each group and collected supernatant from macrophages cultures. As shown in Fig. 5, supernatant from macrophages culture of Model group and BMSC group induced significantly higher levels of cell apoptosis of endothelial cells. Furthermore, supernatant of macrophages from BMSC group induced even more apoptosis than that from Model group (

Apoptosis of endothelial cells induced by supernatant of macrophages. Macrophages were collected from abdomen cavity of each animals.
Previous studies have demonstrated that BMSC are immunosuppressive, especially for suppression of allogenic and autoimmune response. They have been reported to be able to treat different inflammatory diseases [10] and immune disorders as autoimmune diseases and GVHD [11]. Moreover, due to lack of immunogenecity, allogenic or xenogenic MSC may exhibit similar immunosupressive function as autologuous MSC [12,13]. In our study, gram-negative bacilli derived LPS was used to induce endotoxemia in mice. Onset of lung and liver inflammatory injuries was observed within two hours (data not shown). In order to investigate the effect of BMSC on tissue inflammatory injuries at the early stage of endotoxemia, we injected BMSC intravenously one hour after LPS injection. However, our results showed that BMSC were unable to alleviate lung and liver injuries in endotoxemic animals. As shown in Fig. 3, in animals treated with BMSC, inflammatory injuries were comparable to animals in Model group, including thickening of alveolar septum, filtration of inflammatory cells, congestion in alveolar capillary and hepatic sinusoid, and necrosis of hepatic cells. In certain animals treated with BMSC, the inflammatory injuries were even severe. Inflammation may induce damage to capillary wall and result in fluid leakage in lung mesenchyma, which can increase lung wet weight. As shown in Fig. 2, LPS induced lung injury increased wet-to-dry weight ratio. In animals treated with BMSC, the ratio was comparable to that in animals of Model group (
Although in most studies BMSC were shown to play an effective role in inhibiting inflammation or immunity, they didn’t make the same effects in our study with LPS-induced endotoxemia. Two possibilities may exist: (1) immunomodulary effect of BMSC was changed after injection into our models, or (2) timepoint of BMSC infusion was not appropriate. It has been demonstrated that MSC possessed an immune plasticity, which meaned that their immunomodulary function was bi-directional [14]. Some researchers suggested two types of MSC might exist: MSC1, which are pro-inflammatory, and MSC2, which are anti-inflammatory [15]. Many factors may affect or change the biological function of MSC, including inflammatory factors TNF-α [10], IFNγ [16], IL10 [17] and receptors as TLRs [18,19], etc. Romieu-Mourez, R. et al has demonstrated that IFNγ priming induced human BMSC into an APC phenotype, which released a series of pro-inflammatory factors as IL-1β, IL-6, IL-8/CXCL8, and CCL5, and attracted neutrophils. An in vivo study showed that in IFNγ-treated mice, BMSC could also induce filtration of neutrophils, eosinophils, NK cells, CD11c-DC cells, and CD3 T cells [16]. Furthemore, TLRs have been shown to be expressed on human and mice MSC, which can affect biological functions of MSC [18–21]. However, different even opposite effects of TLRs on immunomodulary function of MSC have been described by different research groups. Our further studies have also shown that rat BMSC express TLR4 upon LPS stimulation, which could also change immunomodulatory ability of BMSC (data not shown).
Damage to the endothelial layer of blood vessels caused by inflammatory mediators can lead to capillary leak syndrome, dilation of blood vessels, septic shock and systematic organ dysfunction in endotoxemia. Macrophages are key inflammatory cells involved at the early stage of endotoxemia [22]. In this study, we isolated macrophages from abdomen cavity of three animal groups. Supernatant of these macrophages restimulated shortly with LPS was used to stimulate endothelial cells. We showed that macrophages from animals of BMSC group induced highest apoptosis rate. It suggested that macrophages from mice of BMSC group secreted highest levels of inflammatory factors which could induce more cell apoptosis.
The interface between MSC and immune cells in vivo is complex, and the local microenvironment may largely influence the behavior of MSC [14]. A lot of work remains to be further studied, including whether or not MSC could be effective in treating inflammatory diseases elicited by infection, at what stage of disease should MSC be administrated, etc.
Conclusion
In conclusion, our study showed that infusion of rat BMSC at early stage of endotoxemia could not effectively improve inflammatory damage in lung and liver tissues in mice models.
Footnotes
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No: NSFC 81270411) and by Program of Wuhan Science and Technology Bureau (No: 2015060101010045).
Conflict of interest
The authors have no conflict of interest to report.