
Abstract
BACKGROUND:
We previously demonstrated that a bioabsorbable nerve conduit coated with mouse induced pluripotent stem cell (iPSC)-derived neurospheres accelerated peripheral nerve regeneration in mice.
OBJECTIVE:
We examined the fate and utility of iPSC-derived neurospheres transplanted with nerve conduits for the treatment of sciatic nerve gaps in mice.
METHODS:
Complete 5-mm defects were created in sciatic nerves and reconstructed using nerve conduits that were either uncoated or coated with mouse iPSC-derived neurospheres. The survival of the neurospheres on the nerve conduits was tracked using an in vivo imaging. The localization of the transplanted cells and regenerating axons was examined histologically. The gene expression levels in the nerve conduits were evaluated.
RESULTS:
The neurospheres survived for at least 14 days, peaking at 4--7 days after implantation. The grafted neurospheres remained as Schwann-like cells within the nerve conduits and migrated into the regenerated axons. The expression levels of ATF3, BDNF, and GDNF in the nerve conduit coated with neurospheres were upregulated.
CONCLUSIONS:
Mouse iPSC-derived neurospheres transplanted with nerve conduits for the treatment of sciatic nerve defects in mice migrated into regenerating axons, survived as Schwann-like cells, and promoted axonal growth with an elevation in the expression of nerve regeneration-associated trophic factors.
Keywords
Introduction
Autogenous nerve grafts are currently the gold standard for treatments to repair peripheral nerve defects; however, they sacrifice some intact peripheral nerves, leading to donor site morbidity. Bioabsorbable nerve conduits have recently become available as treatment options for peripheral nerve reconstruction without sacrificing autogenous intact nerves [1]. However, such treatments are not perfect; nerve regeneration achieved using nerve conduits remains poorer than that using autologous nerve grafts [2]. To overcome this problem, strategies and basic studies for improving nerve regeneration using nerve conduits have been designed. They involve the addition of supportive cells such as Schwann cells, bone marrow-derived mesenchymal stem cells, and adipose-derived stem cells and growth factors to promote axonal growth via the nerve conduits [3–7].
We have previously developed a tissue-engineered bioabsorbable nerve conduit coated with induced pluripotent stem cell (iPSC)-derived neurospheres containing neural stem/progenitor cells [8,9]. We have already demonstrated that the nerve conduit coated with mouse iPSC-derived neurospheres accelerates the regeneration and functional recovery of peripheral nerves in both the short and long terms for the treatment of sciatic nerve defects in young mice [10–12]. The addition of mouse iPSC-derived neurospheres also enhanced peripheral nerve regeneration via the nerve conduits in aged mice (92 weeks old) with lower regenerative capacity [13].
However, it remains unclear how these iPSC-derived neurospheres track and promote axonal regeneration after implantation with the nerve conduit. In this study, we examined the fate of mouse iPSC-derived neurospheres transplanted with nerve conduits and their utility in the treatment of sciatic nerve gaps in both young and aged mice, using histological and in vivo imaging. The expression levels of nerve regeneration-associated factors were also examined to elucidate the mechanism that promotes axonal regeneration and repairs sciatic nerve gaps.
Methods
Ethics committee approval
The experiments were carried out in strict accordance with the Institutional Guidelines for the Care and Use of Laboratory Animals of Osaka City University (approval No. 10025 on May 19, 2011).
Cell culture, neural induction, and lentivirus transduction
Mouse iPSCs from the iPS-MEF-Ng-178B-5 and iPS-Stm-FB/gfp-99-1 cell lines were provided by RIKEN BRC via the National Bio-Resource Project of MEXT, Japan [14,15]. The iPSC culture and neural induction were performed in the same manner as described in detail in our previous report [9–13]. Briefly, the iPSCs were differentiated into primary neurospheres containing neural stem/progenitor cells via embryoid body formation and the primary neurospheres were passaged into secondary neurospheres, which were used for transplantation. Forty eight hours before seeding onto nerve conduits, the iPSC (iPS-MEF-Ng-178B-5)-derived secondary neurospheres were dissociated and infected with a lentivirus expressing ffLuc, a green fluorescence protein (modified from Venus) fused to a luminescence protein (Luciferase 2), under the control of an elongation factor (EF) promoter (pCSII-EF-Venus-Luc2), for bioluminescence imaging of the graft [16–20].
Nerve conduit
The nerve conduit (outer diameter: 2-mm; inner diameter: 1-mm; length: 7-mm) used in this study was identical to those used in the many of our previous studies for the treatment of sciatic nerve defects (Fig. 1) [9–13,21,22]. This bioabsorbable nerve conduit consists of two layers: an outer layer of a poly(L-lactide) (PLA) multifilament fiber mesh and an inner layer of a PLA/polycaprolactone (PCL) (50:50) sponge copolymer. The nerve conduit was elastic enough to maintain its tubular structure during promised axonal growth but flexible enough to allow easy handling [10–12]. The PLA and PCL copolymer sponge of the inner layer has a honeycomb structure with pore sizes of 10–50 μm, into which regeneration-facilitating cells such as iPSC-derived neurospheres can enter and proliferate as a scaffold [9,12].

Gross appearance (A) and cross section (B) of the artificial nerve conduit transplanted to repair sciatic nerve 5-mm defects in mice (C).
Nerve conduits coated with mouse iPSC-derived neurospheres were prepared according to the same procedures described in detail in our previous reports [9–13]. Briefly, the Day 7 iPSC-derived secondary neurospheres labeled with the lentivirus for expressing ffLuc were dissociated into single cells using TrypLE Select (Gibco, Tokyo, Japan) and were carefully seeded over each nerve conduit at a density of 2.0 to 4.0 × 106 cells per conduit after prewetting the conduit with 70% ethanol and rinsing with physiological saline. Then, these nerve conduits were placed in Dulbecco’s modified Eagle’s medium supplemented with 10% embryonic stem cell-qualified fetal bovine serum (all from Gibco Life Technologies, CA, USA) for 14 days. This completed the process, described in our previous detailed reports, of three-dimensionally coating the nerve conduits with iPSC-derived secondary neurospheres [9–13]. Thus, iPSC-derived secondary neurospheres were histologically differentiated into Schwann-like cells, which played the most important role in peripheral nerve regeneration; these iPSC-derived Schwann-like cells had engrafted three-dimensionally in the nerve conduits as a scaffold [9,12].
Animal model
Male 6-week-old and 96-week-old mice (C57BL/6) were used as the young and aged mice, respectively, in this study [10,11,13]. They were purchased from Japan SLC (Hamamatsu, Japan) and housed in an air-conditioned room with free access to food and water. Complete 5-mm long defects were created in the left sciatic nerves of the young and aged mice [13]. The nerve defects were reconstructed using nerve conduits that were either uncoated or coated with iPSC-derived neurospheres. Under a microscope, 1-mm lengths of both the proximal and distal stumps of the sciatic nerve were pulled into the nerve conduit and the nerve ends were sutured, at two proximal and distal locations, to the lumen wall, using 9-0 nylon sutures in a horizontal mattress pattern (Fig. 1).
Bioluminescence imaging analysis
The survival of the mouse iPSC (iPS-MEF-Ng-178B-5)-derived neurospheres on the nerve conduits was continuously tracked in the young and aged mice (n = 3 from each group) using in vivo analysis. A luciferase substrate, d-luciferin (Perkin Elmer, Waltham, MA, USA), was directly infused (50 μl) around the nerve conduit after exploring it under inhalation anesthesia using isoflurane, and the ffLuc-labelled cells were monitored using an In Vivo Imaging System (IVIS) Spectrum instrument and a CCD optical macroscopic imaging system (Perkin Elmer, Waltham, MA, USA). The bioluminescence signals observed in the luminescent imaging mode (exposure time; 1 min, field of view: 5 cm2) were measured 0, 4, 7, 14, and 28 days after transplantation [17,18]. The survival rate of the iPSC-derived neurospheres was quantified by setting the amount of luminescence on the day of transplantation (Day 0) to 100 and expressing the increase or decrease in percentages.
Histological evaluation
Four, 7, and 14 days after repairing the peripheral nerve defects, the nerve conduits coated with mouse iPSC (iPS-Stm-FB/gfp-99-1)-derived neurospheres in the young mice were harvested, fixed in 4% paraformaldehyde overnight at 4 °C, and embedded in paraffin. The localization of the transplanted cells and regenerating axons was examined in longitudinal sections of the nerve conduits, using hematoxylin and eosin (HE) staining and immunohistochemistry with an anti-GFP antibody (Invitrogen, Rockford, USA) labeled with Alexa Fluor 488 as a marker of the grafted cells, an anti-S-100β antibody (Abcam, Cambridge, UK) as a marker of the Schwann-like cells, and a secondary antibody (Abcam, Cambridge, UK) labeled with Alexa Fluor 647 [9,12,13]. The observation was performed with a BZ-8000 fluorescence microscope (KEYENCE, Osaka, Japan) [12].
Gene expression
To elucidate the mechanism that promotes axonal regeneration, the gene expression levels of nerve regeneration-associated factors in the nerve conduits with and without mouse iPSC-derived neurospheres were evaluated in both young and aged mice. The nerve conduits with (iPS group) and without (control group) iPSC (iPS-MEF-Ng-178B-5)-derived neurospheres were taken out from the young and aged mice 4 and 7 days after transplantation (n = 5 in each group). They were immersed in an RNA preservation reagent (RNAlater solution; Ambion, Austin, TX, USA) and stored at −20 °C. The total RNA from the tissues including the nerve conduits was isolated using a commercially available kit (QIAzol Lysis Reagent and RNeasy Plus Universal Mini Kit; Qiagen, Venlo, The Netherlands) and the cDNA was synthesized using a synthesis kit (iScript, Bio-Rad, Hercules, CA, USA). Then, RNA levels in the nerve conduits were determined using quantitative real-time PCR kits (PowerUp SYBR Green Master Mix, Thermo Fisher Scientific, USA) in the same manner as described in detail in our previous report [21]. The expression of the nerve regeneration–associated factors, activating transcription factor 3 (ATF3), brain-derived neurotrophic factor (BDNF), and glial cell line-derived neurotrophic factor (GDNF), was measured [5,6,23,24]. HPRT-1 was used as an endogenous control.
Statistical analysis
All the experimental data are expressed as the mean ± standard deviation (SD). The Mann-Whitney U test was performed using the GraphPad Prism7 software (GraphPad Software Inc., San Diego, CA, USA) to compare the iPS and control groups. p < 0.05 denoted statistical significance.
Results
Tracking transplanted iPSC-derived neurospheres in mice
Cell tracing in the IVIS showed that the mouse iPSC-derived neurospheres grafted on the nerve conduits survived for at least 14 days and disappeared 28 days after transplantation in both the young and aged mice (Fig. 2). The luminescence photon counts were higher after 4 and 7 days from transplantation than on the day of transplantation and decreased gradually in both the young and aged mice; the counts on Days 4 and 7 were generally higher in the young mice than in the aged mice.

Images (A) and quantitative analyses (B) of the photon counts in the in vivo imaging system. Bars and lines represent mean ± SD.
In the longitudinal sections of the nerve conduits, regenerated axonal fibers elongated gradually along the inner wall surface of the conduits from the proximal nerve stumps to the distal through Days 4, 7, and 14 after transplantation (Fig. 3A). The transplanted iPSC-derived Schwann-like cells were found around the proximal and distal regenerated axons (Fig. 3B). The immunohistochemical analysis showed that the cells that tested double positive for anti-GFP and anti-S-100 antibodies were in the inner layer of the nerve conduits and around the proximal and distal nerve stumps on Day 7 after transplantation (Fig. 3C). Some of these double-positive cells migrated into the proximal and distal ends of the regenerated axons. These results indicated that the grafted iPSC-derived neurospheres, which had partially differentiated into Schwann-like cells, remained so within the nerve conduits and then migrated into the proximal and distal regenerated axons to promote axonal growth.
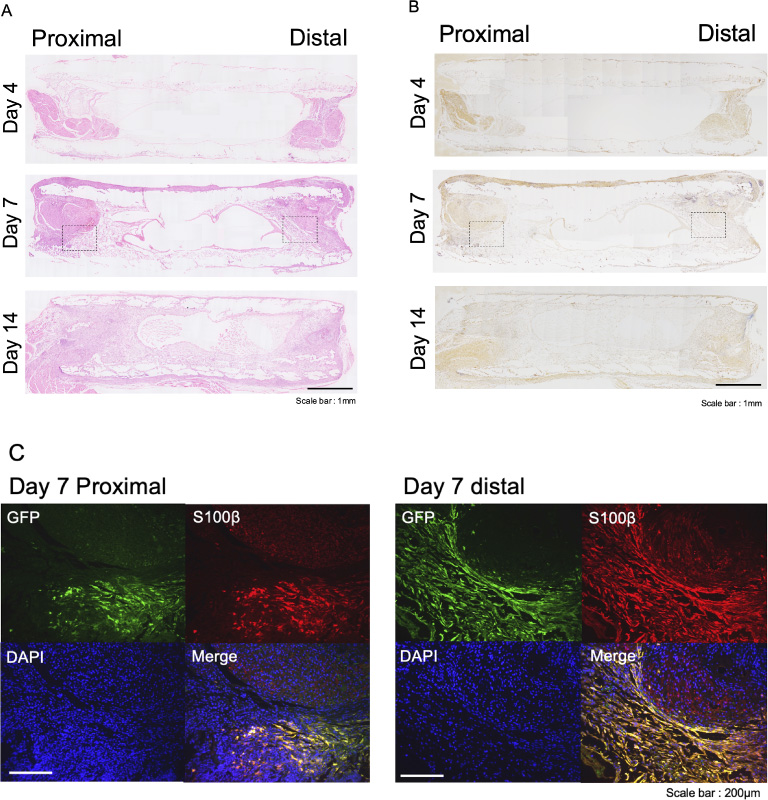
Histological images of longitudinal sections of nerve conduits coated with iPSC-derived neurospheres transplanted in young mice, using hematoxylin and eosin staining (A), immunohistochemical staining of anti-S-100 antibody (B), and fluorescence immunostaining (C). Scale bar: 1 mm (A and B) and 200 μm (C).
Expression levels of the nerve regeneration-associated factors, ATF3, BDNF, and GDNF, in the nerve conduits with and without iPSC-derived neurospheres after repairing the sciatic nerve gaps in the young and aged mice are shown in Fig. 4. In both the young and aged mice, the expression of ATF3, BDNF, and GDNF was higher in the iPS group compared to that in the control group on Day 4 after implantation. This was especially true for ATF3 expression in the young mice. In the aged mice, the expression levels of BDNF and GDNF were significantly higher in the iPS group than in the control groups on Day 4 after implantation. These increases in the expression levels of ATF3, BDNF, and GDNF were more significant after 4 days from implantation than after 7 days in both the young and aged mice. However, in the young mice, the expression levels of BDNF were significantly higher in the iPS group than in the control group on Day 7 after implantation.
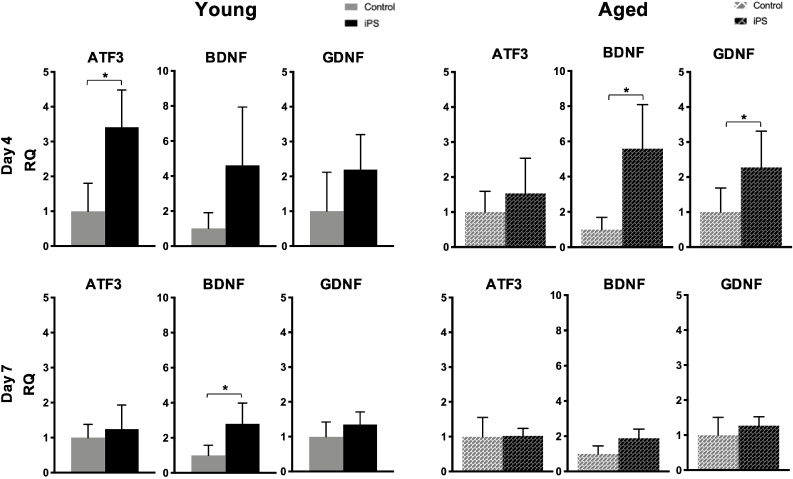
Gene expression levels of ATF3, BDNF, and GDNF in nerve conduits with and without mouse iPSC-derived neurospheres in young and aged mice. Bars and lines represent mean ± SD. *p < 0.05.
Mouse iPSC-derived neurospheres transplanted with nerve conduits for the treatment of sciatic nerve defects in young and aged mice survived for at least 14 days as Schwann-like cells and migrated into the proximal and distal regenerating axons, leading to the promotion of axonal growth with an elevation in the expression of trophic factors such as ATF3, BDNF, and GDNF.
The fate and contribution of the stem cells transplanted with nerve conduits remain controversial [25–27]. In this study, the transplanted iPSC-derived neurospheres had completely disappeared within 4 weeks due to immune rejection of the allografts because the grafted mouse iPSCs (from the iPS-MEF-Ng-178B-5 and iPS-Stm-FB/gfp-99-1 cell lines) did not originate from the same C57BL/6 strain as the recipient mice of sciatic nerve defect models. Briefly, the iPS-MEF-Ng-178B-5 cell line was generated from the fibroblasts derived from the F1 Hybrid mouse of the 129S6 and C57BL/6 strains [14,28]. The iPS-Stm-FB/gfp-99-1 cell line was generated by backcrossing C57BL/6 and 129SvJae strain mice [15,29,30]. Some authors have demonstrated that undifferentiated rat adipose tissue-derived stem cells (ADSCs) grafted with nerve conduits disappeared 14 days after allogenic transplantation in rats; the effect of the transplanted ADSCs on the axonal growth could have been mediated by an initial boost in released growth factors and/or by an indirect effect on endogenous Schwann cell activity [25]. Sowa et al. also reported that mouse ADSCs with gelatin hydrogel tubes survived for 4 weeks after allogenic transplantation in mice, without differentiating into Schwann cells, and promoted the regeneration of axons, formation of myelin, and reversal of denervation muscle atrophy [31]. Others have demonstrated that transplanted human bone marrow stromal cells (MSCs) survive 3 weeks after xenotransplantation in rats and support the regeneration of axons while retaining Schwann cell properties [26]. Wang et al. reported that neural crest stem cells derived from human iPSCs and embryonic stem cells and transplanted with nanofibrous nerve conduits survived for 4 weeks after xenotransplantation in rats; they differentiated into Schwann cells and were integrated into the myelin sheath around axons, promoting the regeneration of peripheral nerves [27]. Therefore, stem cells transplanted with nerve conduits survive for several weeks either as Schwann cells or without differentiating, and contribute to axonal regeneration via indirect trophic effects and/or direct myelin reformations. We believe that this is because the survival of the grafted cells depends on the degree of the immune response of the recipient animals.
In this study, we chose to examine the nerve regeneration–associated factors, ATF3, BDNF, and GDNF, because they are known to promote axonal growth, especially in relation to Schwann cells and aging; earlier reports state that their levels are elevated in the regenerated axons in the early stage after transection of the sciatic nerve [5,6,24,32]. ATF3, a transcription factor that is rapidly expressed in Schwann cells after peripheral nerve injury, supports axonal regeneration in response to stress on nerves [23,33]. It has also been revealed that the expression of ATF3 in Schwann cells is higher in aged mice [34]. BDNF, a neurotrophin related to the canonical nerve growth factor that is synthesized by motoneurons and Schwann cells, promotes axonal regeneration and re-myelination in injured peripheral nerves [35]. GDNF, a neurotrophic factor synthesized by Schwann cells, affects the survival and repair of injured peripheral neurons and promotes axonal guidance and re-myelination of small-caliber axons [36]. The expression of both BDNF and GDNF decreases in injured peripheral nerves with aging [32].
In this study, the addition of iPSC-derived neurospheres upregulated the expression of ATF3 only in young mice and not in aged mice. Since ATF3 was downregulated in the injured peripheral nerve stumps in the younger mice, as described above, the elevations in ATF3 expression upon the addition of iPSC-derived neurospheres were more significant in the young mice than in the aged mice. In contrast, the addition of iPSC-derived neurospheres upregulated the expression of BDNF and GDNF in both the young and aged mice. The upregulation was more significant in the aged mice, which had inherently lower BDNF and GDNF levels. Additionally, these increases in the expression levels of ATF3, BDNF, and GDNF were more emphasized after 4 days from implantation than after 7 days in both the young and aged mice. The possible reasons why the extent of mRNA upregulation in the iPS group in young mice was lower in Day 7 than in Day 4 were as follows. First, the peak of survival rate of the transplanted iPSC-derived neurospheres in young mice might be actually in Day 5 or 6, not in Day 7 (Fig. 2). Second, the mRNA expressions of these trophic factors in the iPS group would be upregulated earlier than the protein expressions of them, which might be higher in Day 7. These results suggest that the indirect effects of the transplanted iPSC-derived neurospheres (Schwann-like cells) on axonal regeneration may be mediated by an initial release of these trophic factors in the earlier stage.
Conclusions
Mouse iPSC-derived neurospheres transplanted with nerve conduits for the treatment of sciatic nerve defects in young and aged mice migrated into regenerating axons, survived for at least 14 days as Schwann-like cells, and promoted axonal growth with an elevation in the expression of nerve regeneration-associated trophic factors such as ATF3, BDNF, and GDNF.
Footnotes
Acknowledgements
The authors thank Masaya Nakamura MD, PhD, Kazuki Sato MD, PhD, Narihito Nagoshi MD, PhD (Department of Orthopaedic Surgery, Keio University School of Medicine, Japan), Hideyuki Okano MD, PhD, and Shinsuke Shibata MD, PhD (Department of Physiology, Keio University School of Medicine, Japan) for material transfer with regard to the EF promoter (pCSII-EF-IIVenus-Luc2) for the lentivirus expressing ffLuc and their excellent assistance with neural induction of the iPS cells. The authors also thank Takao Omura MD, PhD (Department of Orthopaedic Surgery, Hamamatsu University School of Medicine) for advisements about the gene expression of nerve regeneration-associated factors. Finally, we would like to thank Editage (![]() ) for English language editing.
) for English language editing.
Conflict of interest
None to report.
Funding
This work was supported by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Number JP19K12760.