
Abstract
BACKGROUND:
Circular RNA circ_0124644 has crucial regulation in the progression of coronary artery diseases, including atherosclerosis (AS). The aim of this study was to explore the regulatory mechanism of circ_0124644 in oxidized low-density lipoprotein (ox-LDL)-induced endothelial injury in human umbilical vein endothelial cells (HUVECs).
METHODS:
Cell viability and proliferation were assessed using cell counting kit-8 (CCK-8) assay and EdU assay. The apoptosis detection was performed by flow cytometry. Angiogenesis was evaluated through tube formation assay. The protein analysis was conducted via western blot. Inflammatory cytokines were examined by enzyme-linked immunosorbent assay (ELISA). The expression determination of circ_0124644, microRNA-370-3p (miR-370-3p) and forkhead box protein O4 (FOXO4) was performed through reverse transcription-quantitative polymerase chain reaction (RT-qPCR). Dual-luciferase reporter assay and RNA immunoprecipitation (RIP) assay were used to analyze the interaction between targets.
RESULTS:
Treatment of ox-LDL resulted in the inhibition of cell viability, proliferation and angiogenesis but the promotion of apoptosis and inflammation in HUVECs. These ox-LDL-induced cell damages were alleviated after the downregulation of circ_0124644. Circ_0124644 interacted with miR-370-3p, and the regulatory role of circ_0124644 was associated with the sponge function of miR-370-3p. Additionally, miR-370-3p targeted FOXO4 and circ_0124644 increased the expression of FOXO4 through acting as a sponge of miR-370-3p. Overexpression of miR-370-3p protected from ox-LDL-induced injury via the downregulation of FOXO4.
CONCLUSION:
All results revealed that circ_0124644 accelerated endothelial injury in ox-LDL-treated HUVECs by mediating miR-370-3p-related FOXO4 expression.
Introduction
Atherosclerosis (AS) is one of the main causes of cardiovascular morbidity and mortality worldwide [1]. Low-density lipoprotein particles can be uptake to accumulate large amounts of cholesterol in cells, thus contributing to the development of other diseases in human organs [2]. Endothelial cell apoptosis and inflammation are closely correlated to the progression of AS [3, 4]. To explore the molecular mechanism of endothelial dysfunction is essential for understanding AS pathogenesis.
Circular RNAs (circRNAs) have covalently closed-loop structures and act as microRNA (miRNA) sponges in human diseases [5]. A handful of circRNAs are involved in the pathophysiologic processes of AS, including endothelial injury [6]. Hsa_circ_0124644 (circ_0124644) has exhibited important diagnostic value in coronary artery disease [7]. Wang et al. attested that circ_0124644 exacerbated oxidized low-density lipoprotein (ox-LDL)-induced vascular endothelial dysfunction via miR-149-5p-mediated upregulation of PAPP-A [8]. It remains unknown whether the function of circ_0124644 is associated with another miRNA/mRNA signal axis.
MiRNAs can affect the progression of diseases by regulating gene expression and aerobic training can affect miRNA, platelet function and the progression of cardiovascular diseases [9]. Tian et al. found that microRNA-370-3p (miR-370-3p) suppressed ox-LDL-triggered oxidative stress and inflammatory response by targeting TLR4 [10]. Forkhead box protein O4 (FOXO4) was identified to aggravate vascular injury, then inhibiting the protective role of miR-328-3p in HUVECs [11]. The relation between FOXO4 and miR-370-3p is still unclear.
Moreover, circ_0003204 repressed endothelial cell function in AS via upregulating TGFβR2 level by targeting miR-370-3p [12]. The present study was designed to investigate circ_0124644 function with miR-370-3p/FOXO4 axis in regulating endothelial injury. The novel molecular network underlying the role of circ_0124644 might contribute to the exploration of AS.
Materials and methods
Serum collection and cell treatment
27 patients have been diagnosed with AS at the First People’s Hospital of Taizhou City. 2 mL peripheral blood was collected into a new tube, then centrifugated at 2000 rpm for 10 min. The serum samples (n = 27) were stored at –80°C. In addition, the control serum specimens (n = 27) were acquired from normal healthy crowds at physical examination center of the First People’s Hospital of Taizhou City. The written informed consent forms were provided by all subjects. The present study was performed based on the Declaration of Helsinki and the approval by Ethics Committee of the First People’s Hospital of Taizhou City.
Human umbilical vein endothelial cells (HUVECs; BEINUO BIOLOGY, Shanghai, China) were cultured with HUVEC cell culture medium (BeNa Culture Collection, Beijing, China) in a 5% CO2 incubator at 37°C. The third passaged cells were used in this research. HUVECs were treated with ox-LDL (Solarbio, Beijing, China) at various concentrations (0μg/mL, 25μg/mL, 50μg/mL, 100μg/mL) for 24 h.
Cell transfection
RNAs were bought from Genepharma (Shanghai, China): small interfering RNAs (siRNAs: si-circ_0124644, si-NC), miRNA mimics (miR-370-3p, miR-NC), miRNA inhibitors (anti-miR-370-3p, anti-miR-NC). The pCD5-ciR expression vector (GENESEED, Shanghai, China) was inserted with circ_0124644 sequence to generate pcD5-ciR-circ_0124644 (circ_0124644) plasmid. Also, pcDNA-FOXO4 (FOXO4) plasmid was constructed through pcDNA plasmid (Invitrogen, Carlsbad, CA, USA). HUVECs were cultured overnight to 70% confluence and exposed to 50μg/mL ox-LDL, then transient transfection by Lipofectamine™ 3000 Kit (Invitrogen) was performed according to the provided specification.
Cell viability assay
The analysis of cell viability was conducted via Cell Counting Kit-8 (CCK-8). After transfection for 24 h in ox-LDL-treated cells, cell incubation of CCK-8 solution (Solarbio) was performed with 10μL/well. 4 h later, optical density at 450 nm was determined by a microplate reader.
Cell proliferation assay
The 24-well plates (Corning Inc., Corning, NY, USA) were seeded with 5×104/well HUVECs overnight, followed by ox-LDL treatment and cell transfection for 24 h. The proliferation ability was examined using EdU Cell Proliferation Kit (Sangon, Shanghai, China). Cells were labeled with EdU solution for 2 h and fixed with 4% paraformaldehyde (Sangon) for 0.5 h, followed by cell detection through staining solution and nucleus dying via Diamidine phenylindole (DAPI; Sangon). The pictures of cells were acquired using fluorescence microscope (Olympus, Tokyo, Japan) and the positive cells were expressed EdU + DAPI merged cells.
Cell apoptosis assay
Annexin V Apoptosis Detection Kit (Sangon) was performed to assess apoptosis phenomenon. Cells were suspended in500μL Binding Buffer, then added with 5μL Annexin V-fluorescein isothiocyanate (FITC) and Propidium Iodide (PI). After incubation for 20 min, cell apoptosis observation was conducted through a flow cytometer (BD Biosciences, San Diego, CA, USA). These cells with staining of Annexin V + /PI– and Annexin V+/PI+ were regarded as the apoptotic cells.
Tube formation assay
The 96-well plates (Corning Inc.) 60μL Matrigel (BD Bioscience) were enveloped with 60μL Matrigel (BD Bioscience), followed by cell seeding with 2×104 per well. After cell culture for 48 h, capillary-like branches were counted in 5 random fields through a computer-assisted microscope.
Western blot
The protein acquisition using Radioimmunoprecipitation assay lysis buffer (Sangon) and concentration determination by BCA Protein Assay Kit (Sangon) were performed as per the instruction books. 50μg proteins were used for expression analysis, and the procedures were in accordance with the previous depiction [13]. The primary antibodies targeting CyclinD1 (ab134175, 1:1000), Cleaved-caspase 3 (ab32042, 1;1000), FOXO4 (ab128908, 1:1000), GAPDH (ab128915, 1:1000) and Goat Anti-Rabbit IgG H&L secondary antibody (ab205718, 1:5000) were purchased from Abcam (Cambridge, UK). GAPDH was applied as a reference gene for expression correction, and protein level analysis was carried out through ImageJ software (NIH, Bethesda, MD, USA).
Enzyme-linked immunosorbent assay (ELISA)
Cell supernatants were harvested after treatment and transfection in HUVECs. Interleukin-1β (IL-1β) and tumor necrosis factor-alpha (TNF-α) concentrations were measured via IL-1β Human ELISA Kit (Invitrogen) TNF alpha Human ELISA Kit (Invitrogen), following the user’s guidelines.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) assay
Total RNA was isolated through Trizol Kit (Beyotime, Shanghai, China). The reverse transcription and PCR detection were performed using BeyoRT™ III First Strand cDNA Synthesis Kit (Beyotime) and BeyoFast™ SYBR Green qPCR Mix (Beyotime), referring to the operating manuals. The level analysis was conducted by the 2–ΔΔCt method, using glyceraldehyde-phosphate dehydrogenase (GAPDH) and U6 as the endogenous control genes. The forward and reverse primer sequences were exhibited in Table 1.
Primer sequences used for RT-qPCR
Primer sequences used for RT-qPCR
Circinteractome (for circ_0124644 and miR-370-3p) and starbase (for miR-370-3p and FOXO4) were adopted for predicting target binding sites. The wild-type and mutant-type (WT and MUT) luciferase plasmids were constructed using pmirGLO vector (Promega, Madison, WI, USA). HUVECs were co-transfected with WT-circ_0124644, MUT-circ_0124644, WT-FOXO4 3’UTR or MUT-FOXO4 3’UTR and miR-NC or miR-370-3p for 48 h. Immediately, the luciferase activity analysis was performed through Dual-luciferase Reporter Detection Kit (Promega).
RNA immunoprecipitation (RIP) assay
The target interaction was further explored via Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA, USA). Protein A magnetic beads were coated with antibody against immunoglobulin G (IgG) or Argonaute-2 (Ago2), followed by cell incubation and RNA isolation. Circ_0124644, miR-370-3p and FOXO4 levels were quantified by RT-qPCR.
Statistical analysis
The experiments were conducted by three independent repetitions with three parallels. Pearson’s correlation coefficient was applied to analyze the linear relationship. The normality of data distribution was analyzed using the Kolmogorov-Smirnov test, and the homogeneity of variances was assessed using the Levene test. Data were displayed as the mean±standard deviation (SD), followed by statistical analysis through SPSS 22.0 (SPSS Inc., Chicago, IL, USA). In addition, the difference was compared via Student’s t-test and analysis of variance (ANOVA) followed by Tukey’s test. Statistically, P < 0.05 was considered as a significant difference.
Results
Cell damages were induced by ox-LDL in HUVECs
HUVECs were exposed to 0μg/mL, 25μg/mL, 50μg/mL, 100μg/mL ox-LDL for 24 h. CCK-8 and EdU assays showed that ox-LDL induced significant inhibition of cell viability (Fig. 1A) and cell proliferation (Fig. 1B). Through the analysis of flow cytometry and tube formation assay, ox-LDL treatment was found to enhance cell apoptosis (Fig. 1C) and suppress angiogenesis (Fig. 1D-E) in HUVECs. Western blot data manifested that CyclinD1 protein expression was downregulated and Cleaved-caspase 3 was upregulated in ox-LDL treatment groups (Fig. 1F). The inflammatory detection by ELISA indicated that ox-LDL increased the concentrations of IL-1β and TNF-α (Fig. 1G). Altogether, ox-LDL reduced cell growth and angiogenesis but promoted cell apoptosis and inflammation in HUVECs.
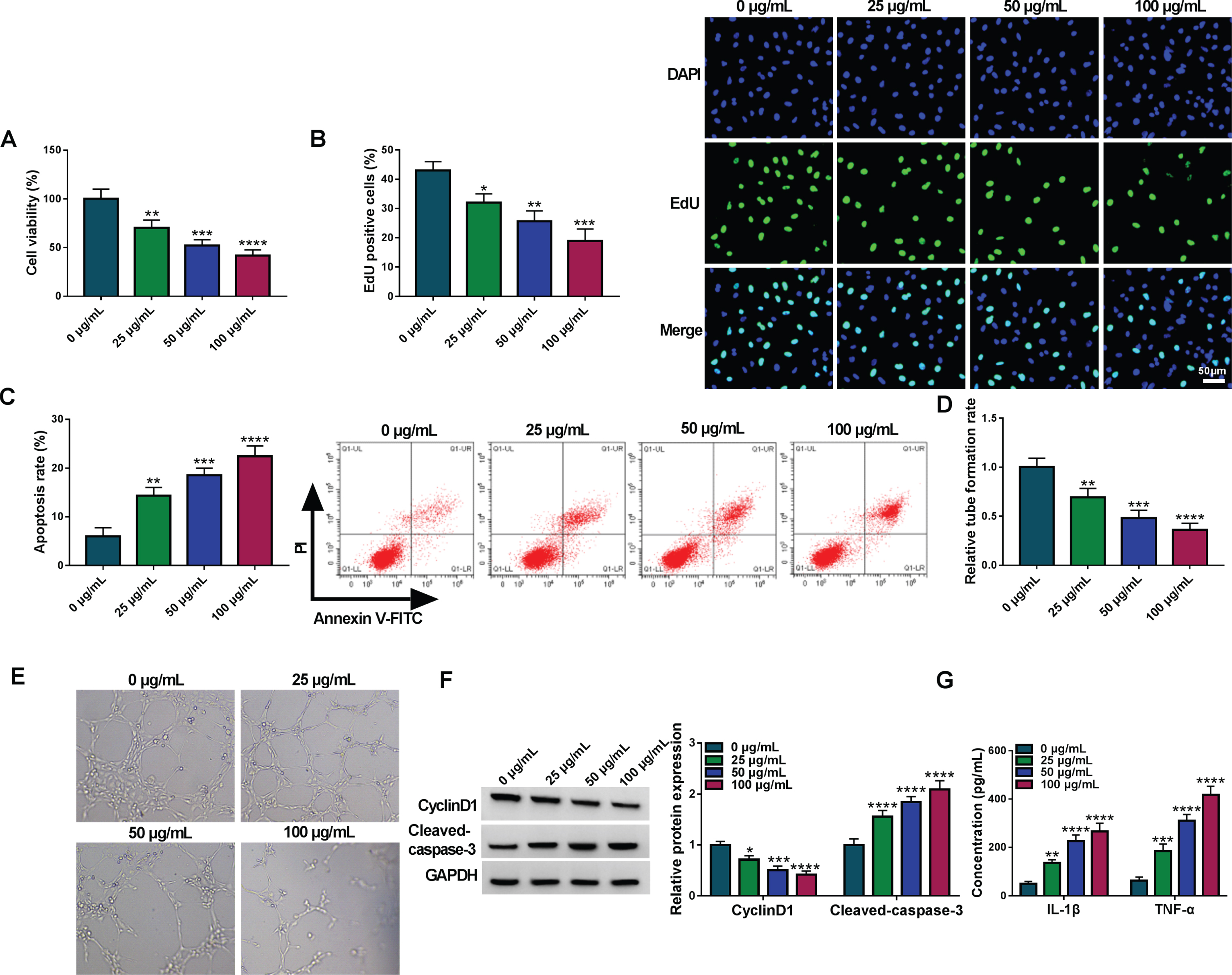
Cell damages were induced by ox-LDL in HUVECs. The ox-LDL treatment with 0μg/mL, 25μg/mL, 50μg/mL, 100μg/mL was performed in HUVECs. (A-B) CCK-8 and EdU assays were performed for the analysis of cell viability (A) and proliferation (B). (C) Flow cytometry was performed for cell apoptosis detection. (D-E) Tube formation assay was conducted for assessing angiogenetic ability. (F) Western blot was conducted for protein determination of CyclinD1 and Cleaved-caspase 3. (G) ELISA was conducted for the examination of inflammatory cytokines. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
The expression quantification exhibited that circ_0124644 was highly expressed in AS serum samples relative to normal serum samples (Fig. 2A). Also, circ_0124644 level was upregulated in 25μg/mL, 50μg/mL and 100μg/mL ox-LDL groups compared with 0μg/mL ox-LDL group (Fig. 2B). Then, 50μg/mL was used in the subsequent assays. Circ_0124644 upregulation by ox-LDL was inhibited after transfection of si-circ_0124644, showing that si-circ_0124644 was efficient to knock down the level of circ_0124644 (Fig. 2C). The influences of ox-LDL on cell viability (Fig. 2D), proliferation (Fig. 2E), apoptosis (Fig. 2F) and tube formation ability (Fig. 2G) were partly abolished by knockdown of circ_0124644. With the downregulation of circ_0124644, the protein changes of CyclinD/Cleaved-caspase 3 (Fig. 2H) and the release of inflammatory cytokines (Fig. 2I) caused by ox-LDL were relieved. Thus, circ_0124644 inhibition has prevented ox-LDL-induced cell injury.
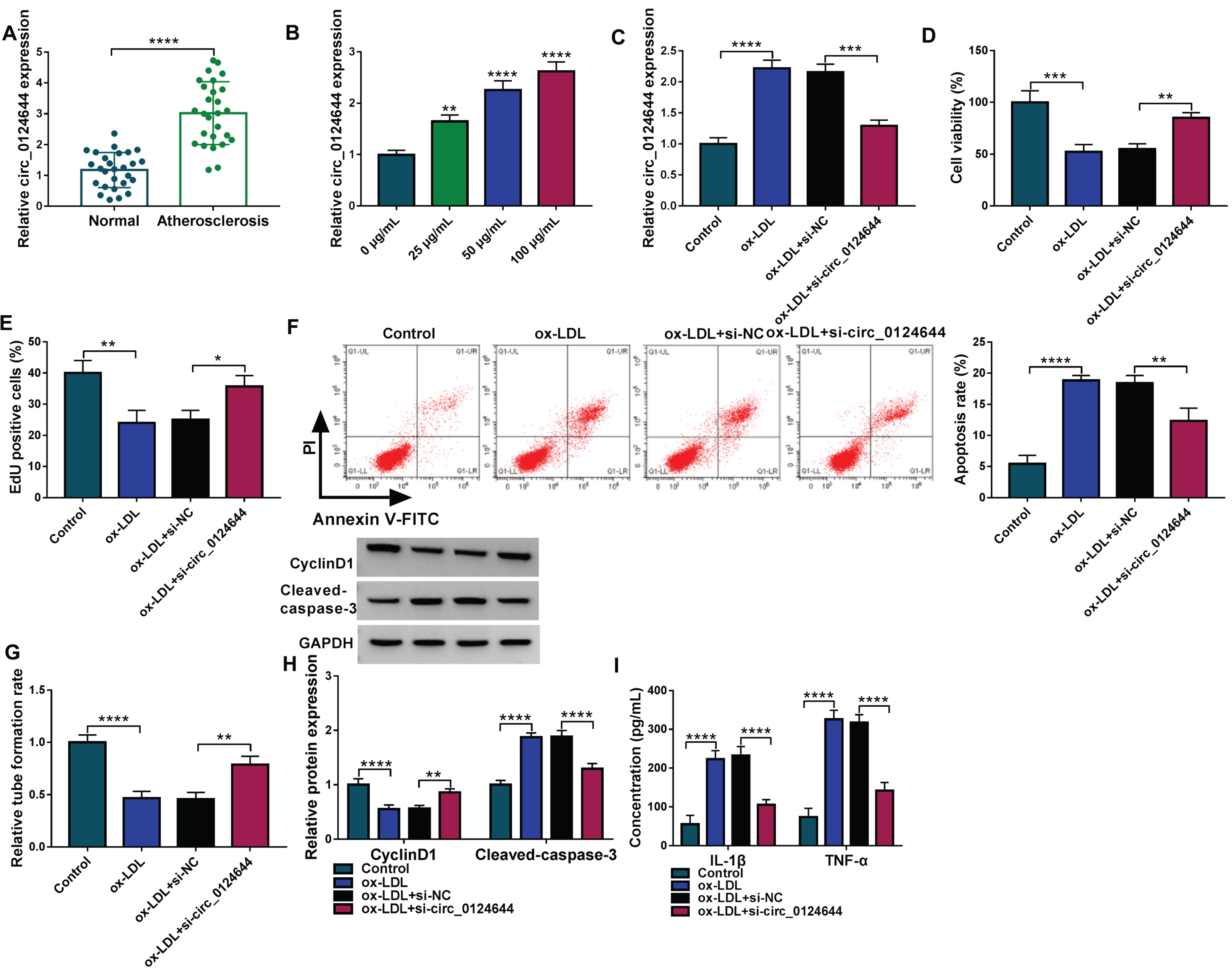
Silence of circ_0124644 alleviated the ox-LDL-induced cell injury in HUVECs. (A-B) The circ_0124644 level was assayed using RT-qPCR in AS serum samples (A) and ox-LDL-treated HUVECs (B). (C) Circ_0124644 expression was detected through RT-qPCR in control, ox-LDL (50μg/mL), ox-LDL + si-NC, ox-LDL + si-circ_0124644 group. (D-E) Cell viability (D) and proliferation (E) were determined using CCK-8 and EdU assays. (F) Cell apoptosis was evaluated using flow cytometry. (G) Angiogenesis was assessed using tube formation assay. (H) CyclinD1 and Cleaved-caspase 3 protein levels were examined using western blot. (I) The concentrations of IL-1β and TNF-α were analyzed using ELISA. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
There was a binding site between circ_0124644 and miR-370-3p sequences by circinteractome (https://circinteractome.nia.nih.gov/), as predicted in Fig. 3A. The miR-370-3p upregulation inhibited luciferase activity of WT-circ_0124644 group but not MUT-circ_0124644 group, suggesting that circ_0124644 could bind to miR-370-3p (Fig. 3B). Furthermore, the enrichment of miR-370-3p and circ_0124644 by Ago2 protein validated the interaction between circ_0124644 and miR-370-3p in HUVECs (Fig. 3C). The expression of miR-370-3p was reduced in AS specimens contrasted with normal specimens (Fig. 3D). We have performed linear analysis between circ_0124644 and miR-3703-p levels, and the result revealed that circ_0124644 was negatively related to miR-370-3p (r = –0.7565, p < 0.001) in AS samples (Fig. 3E). The level of miR-370-3p was downregulated after exposure to ox-LDL with 25μg/mL, 50μg/mL and 100μg/mL (Fig. 3F). The transfection of circ_0124644 enhanced circ_0124644 expression in ox-LDL-treated HUVECs, which demonstrated that overexpression efficiency of circ_0124644 was conspicuous (Fig. 3G). The ox-LDL-mediated miR-370-3p downregulation was ameliorated following knockdown of circ_0124644, while circ_0124644 overexpression aggravated this effect (Fig. 3H). These findings validated that circ_0124644 acted as a sponge of miR-370-3p.
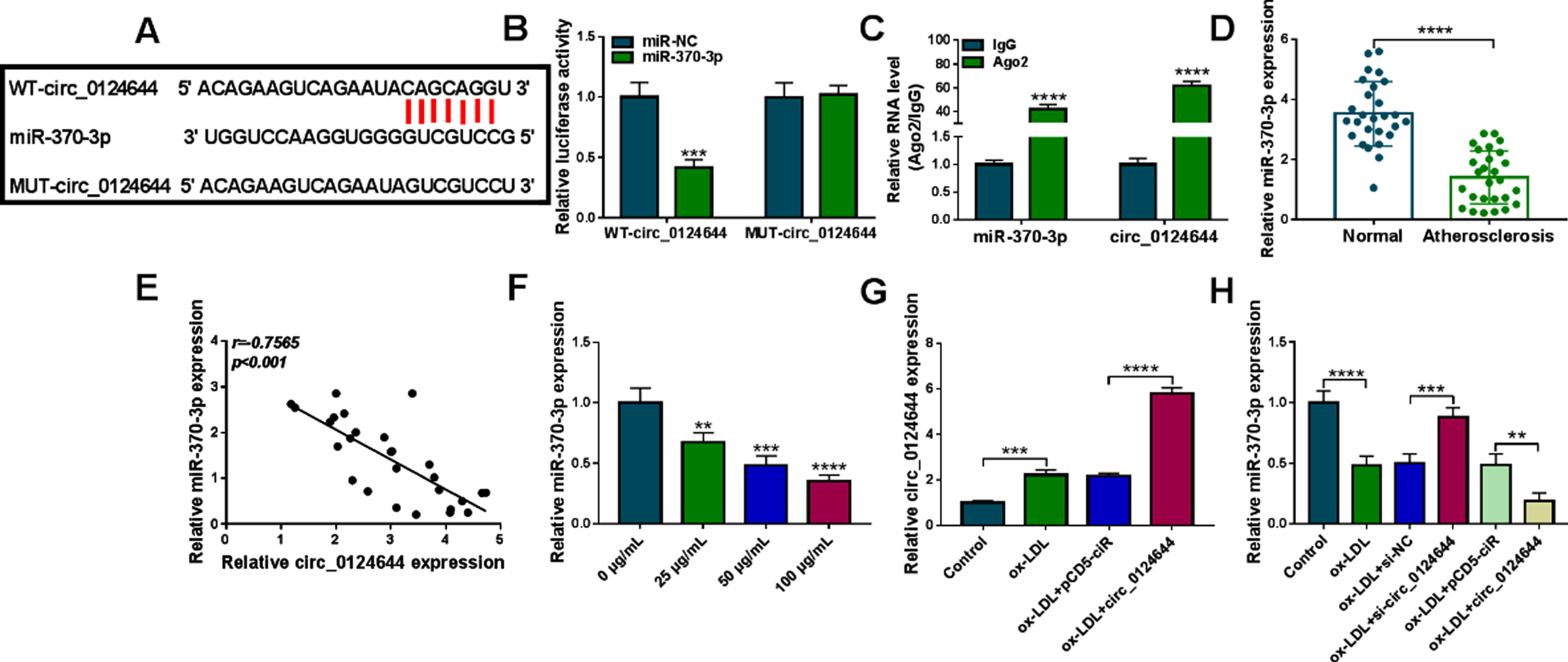
Circ_0124644 was affirmed as a miR-370-3p sponge. (A) Circinteractome was used to predict the binding between circ_0124644 and miR-370-3p. (B-C) The interaction between circ_0124644 and miR-370-3p was confirmed using dual-luciferase reporter assay (B) and RIP assay (C). (D) The expression of miR-370-3p was quantified by RT-qPCR in AS serum samples. (E) The linear relation between circ_0214644 and miR-370-3p was analyzed by Pearson’s correlation coefficient. (F) The miR-370-3p level was detected through RT-qPCR after ox-LDL treatment with different concentrations. (G) The circ_0124644 quantification was performed by RT-qPCR in control, ox-LDL (50μg/mL), ox-LDL + pCD5-ciR, ox-LDL + circ_0124644 group. (H) The determination of miR-370-3p expression was conducted via RT-qPCR in control, ox-LDL (50μg/mL), ox-LDL + si-NC, ox-LDL + si-circ_0124644, ox-LDL + pCD5-ciR, ox-LDL + circ_0124644 group. **P < 0.01, ***P < 0.001, ****P < 0.0001.
The association of miR-370-3p with circ_0124644 in ox-LDL-induced cell injury was further investigated. The transfection of anti-miR-370-3p eliminated si-circ_0124644-mediated upregulation of miR-370-3p in ox-LDL-treated HUVECs, implying that anti-miR-370-3p greatly reduced miR-370-3p level (Fig. 4A). The promotion of cell viability (Fig. 4B) and proliferation (Fig. 4C) but apoptosis inhibition (Fig. 4D-E) and angiogenesis enhancement (Fig. 4F) by si-circ_0124644 were attenuated by miR-370-3p inhibitor in ox-LDL-treated HUVECs. Meanwhile, anti-miR-370-3p has abrogated the reversed effects of si-circ_0124644 on ox-LDL-induced CyclinD1 downregulation or Cleaved-caspase 3 upregulation (Fig. 4G) and inflammatory response (Fig. 4H). The miR-370-3p inhibition was associated with the function of circ_0124644 in ox-LDL-treated HUVECs.
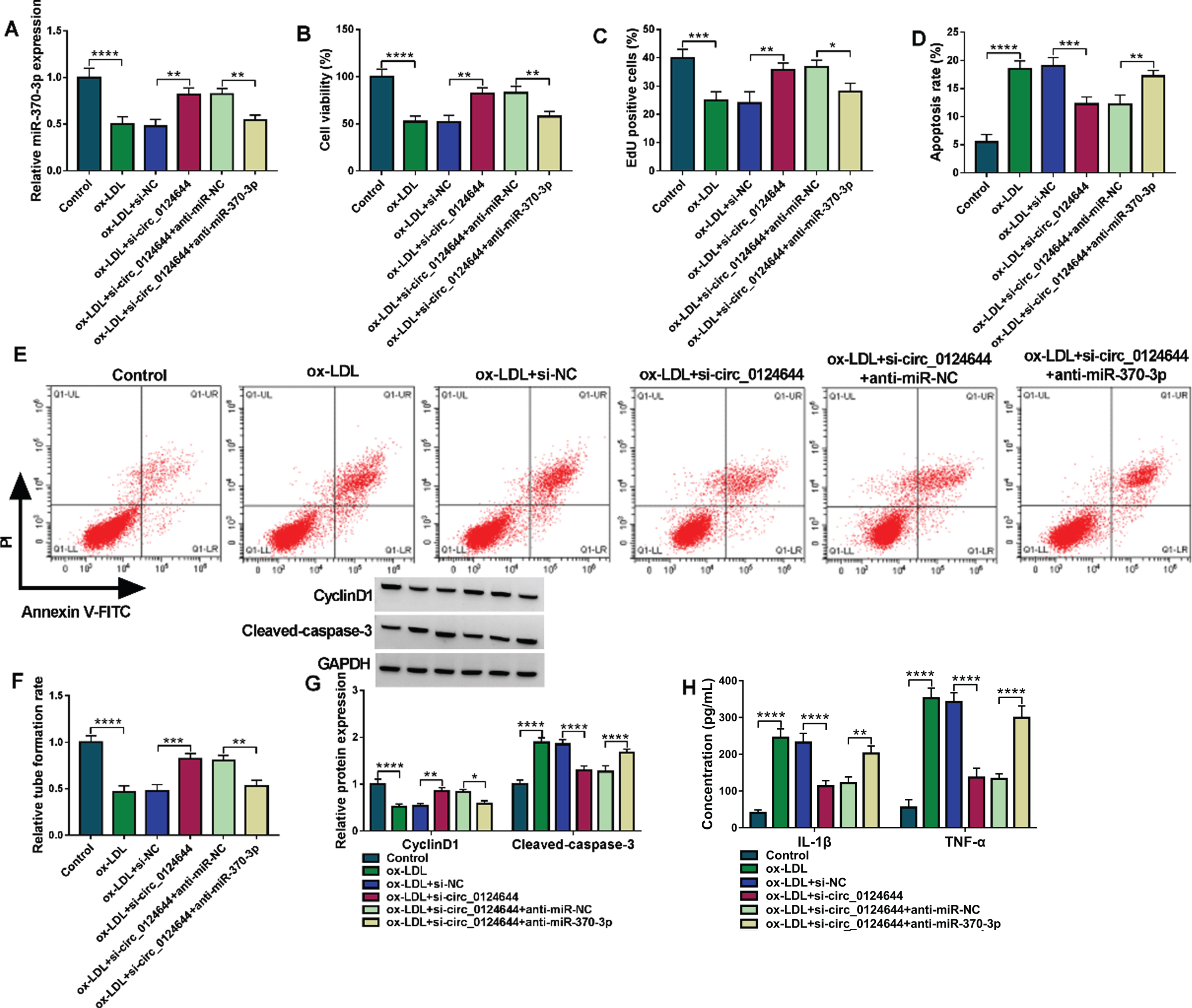
The miR-370-3p expression reduction inhibited the protective function of si-circ_0124644 in ox-LDL-treated HUVECs. HUVECs were treated with control, ox-LDL (50μg/mL), ox-LDL + si-NC, ox-LDL + si-circ_0124644, ox-LDL + si-circ_0124644 + anti-miR-NC, ox-LDL + si-circ_0124644 + anti-miR-370-3p group. (A) RT-qPCR was applied to examine the level of miR-370-3p. (B-C) CCK-8 and EdU assays were applied to assess cell viability (B) and proliferation (C). (D-E) Flow cytometry was applied to measure cell apoptosis. (F) Tube formation assay was applied to determine cell angiogenesis. (G) Western blot was applied to detect the protein expression of CyclinD1 and Cleaved-caspase3. (H) ELISA was applied to analyze the release of inflammatory cytokines. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
The starbase (https://starbase.sysu.edu.cn) has shown that 3’UTR of FOXO4 contained the binding sites of miR-370-3p (Fig. 5A). Dual-luciferase reporter assay (Fig. 5B) and RIP assay (Fig. 5C) affirmed that miR-370-3p interacted with FOXO4 in HUVECs. FOXO4 mRNA level was higher in serum samples from AS patients than that in serum samples from normal controls (Fig. 5D). Interestingly, we observed a negative relation (r = –0.7562, p < 0.001) between the levels of miR-370-3p and FOXO4 in AS samples (Fig. 5E). The upregulation of FOXO4 was detected in ox-LDL-treated HUVECs by western blot (Fig. 5F). Transfection efficiencies of miR-370-3p and anti-miR-370-3p were excellent in ox-LDL-stimulated HUVECs (Fig. 5G). Moreover, ox-LDL-trigged elevation of FOXO4 protein expression was weakened by miR-370-3p overexpression and promoted by miR-370-3p inhibitor (Fig. 5H). Furthermore, the effect of circ_0124644 on FOXO4 was assessed. As shown in Fig. 5I, transfection of si-circ_0124644 after ox-LDL treatment reduced protein expression of FOXO4 while this expression reduction was abated by the downregulation of miR-370-3p. Taken together, miR-370-3p directly targeted FOXO4 and circ_0124644 regulated the level of FOXO4 via absorbing miR-370-3p.
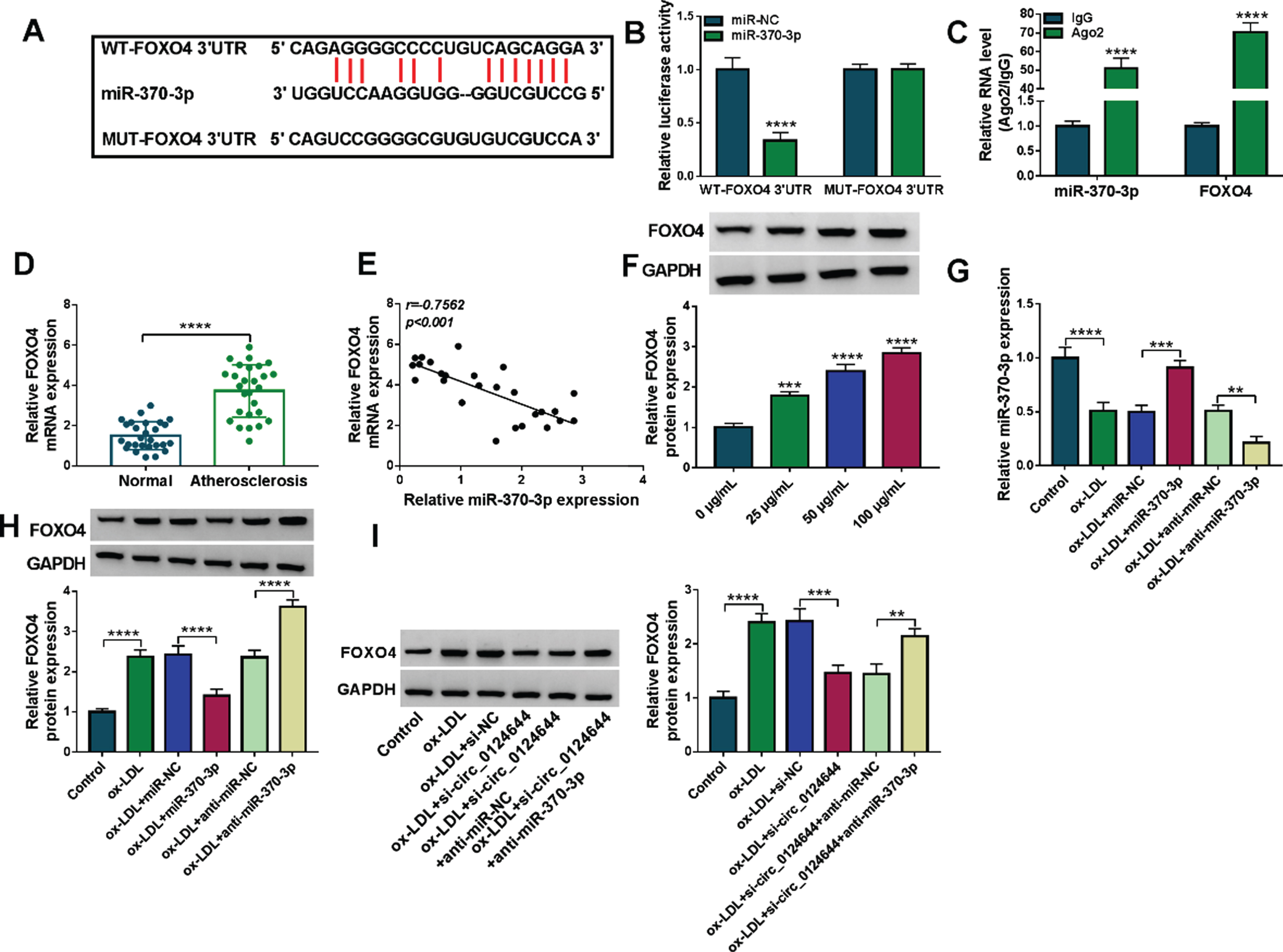
FOXO4 served as a target gene for miR-370-3p. (A) The binding sites of miR-370-3p in the FOXO4 3’UTR sequence was predicted by starbase. (B-C) Dual-luciferase reporter assay (B) and RIP assay (C) were adopted to validate the target binding between miR-380-3p and FOXO4. (D) RT-qPCR was used for mRNA expression detection of FOXO4 in AS serum samples. (E) Pearson’s correlation coefficient was used for linear analysis between FOXO4 and miR-370-3p. (F) FOXO4 protein expression was measured via western blot in ox-LDL-treated HUVECs. (G-H) RT-qPCR and western blot were used for detecting the FOXO4 mRNA and protein levels in control, ox-LDL (50μg/mL), ox-LDL + miR-NC, ox-LDL + miR-370-3p, ox-LDL + anti-miR-NC, ox-LDL + anti-miR-370-3p group. (I) The protein level of FOXO4 was determined via western blot in control, ox-LDL (50μg/mL), ox-LDL + si-NC, ox-LDL + si-circ_0124644, ox-LDL + si-circ_0124644 + anti-miR-NC, ox-LDL + si-circ_0124644 + anti-miR-370-3p group. **P < 0.01, ***P < 0.001, ****P < 0.0001.
The ox-LDL-treated HUVECs were transfected with miR-370-3p, miR-370-3p + FOXO4 or the matched control groups. Western blot manifested that FOXO4 protein expression was upregulated in ox-LDL + miR-370-3p + FOXO4 group compared to ox-LDL + miR-370-3p + pcDNA group (Fig. 6A). FOXO4 transfection offset miR-370-3p-mediated cell viability promotion (Fig. 6B), proliferation acceleration (Fig. 6C), apoptosis suppression (Fig. 6D-E) and tube formation promotion (Fig. 6F) in ox-LDL-treated HUVECs. Overexpression of miR-370-3p returned ox-LDL-induced inhibition of CyclinD1 or upregulation of Cleaved-caspase 3 (Fig. 6G) and concentration elevation of IL-1β or TNF-α (Fig. 6H), whereas the restoration was countermanded with the introduction of FOXO4. The above evidence demonstrated that miR-370-3p protected against ox-LDL-aroused cell injury via targeting FOXO4.
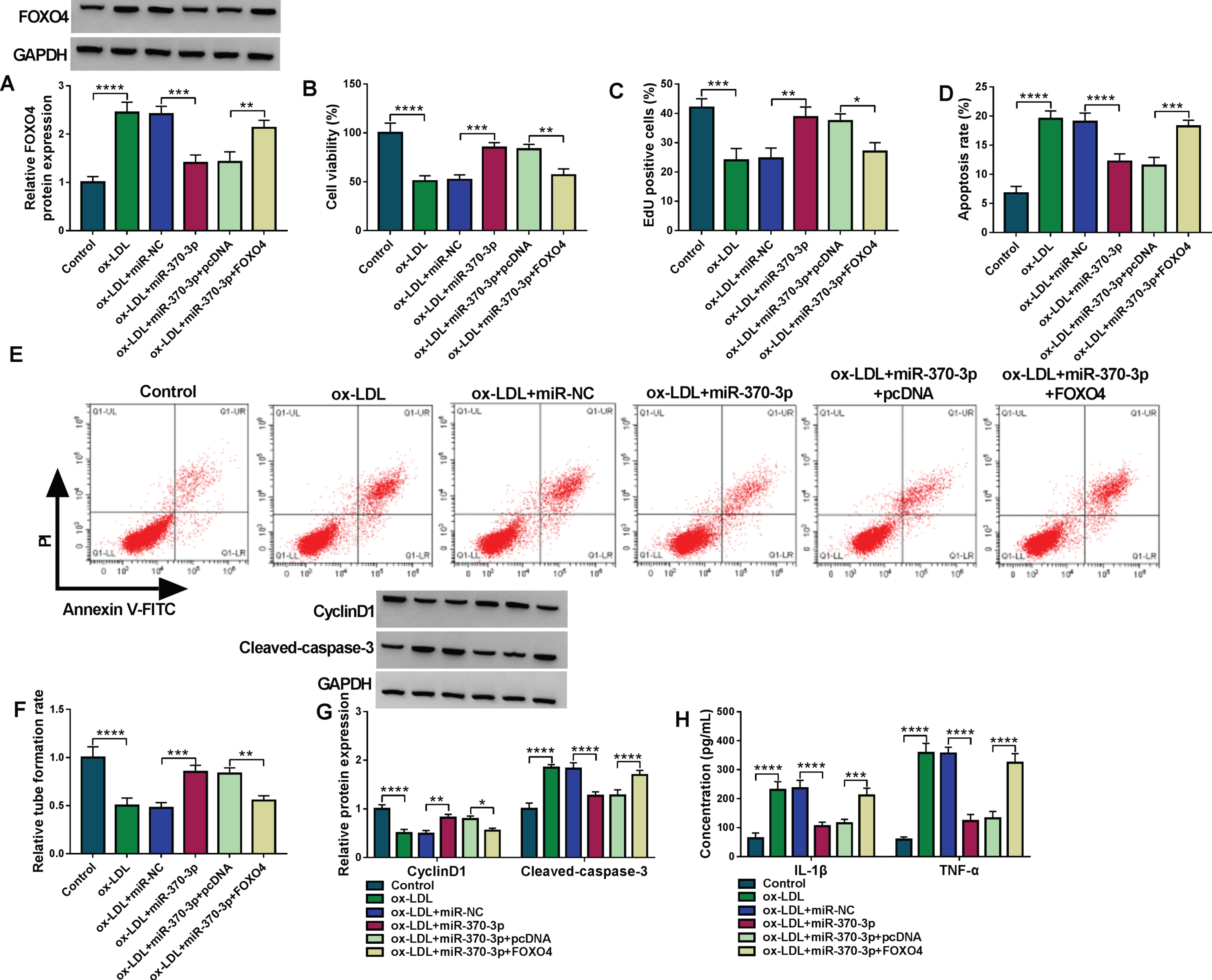
Overexpression of miR-370-3p repressed the ox-LDL-induced cell damages by downregulating the FOXO4 expression. Treatment of control, ox-LDL (50μg/mL), ox-LDL + miR-NC, ox-LDL + miR-370-3p, ox-LDL + miR-370-3p + pcDNA, ox-LDL + anti-miR-370-3p + FOXO4 group. (A) The detection of FOXO4 protein level was conducted through western blot. (B-C) The analysis of cell viability (B) and proliferation (C) was conducted through CCK-8 and EdU assays. (D-E) The measurement of cell apoptosis was conducted through flow cytometry. (F) The assessment of angiogenesis was conducted through tube formation assay. (G) The examination of CyclinD1 and Cleaved-caspase 3 protein expression was conducted through western blot. (H) The evaluation of inflammatory response was conducted through ELISA. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Circ_0124644 has been reported to promote endothelial dysfunction through targeting miR-149-5p to result in the expression elevation of PAPP-A [8]. The current evidence indicated that the regulatory effects of circ_0124644 were achieved partly via mediating miR-370-3p/FOXO4 network, providing a novel sight into circ_0124644 function in ox-LDL-related endothelial injury.
Ox-LDL is a significant risk factor for AS by acting on various cell types, such as endothelial cells, smooth muscles, macrophages and fibroblasts [14]. The accumulation of ox-LDL in the tunica intima leads to the occurrence of AS and the development of atherosclerotic cardiovascular events [15]. Herein, our data showed that ox-LDL reduced cell viability, proliferation and angiogenetic abilities. In addition, ox-LDL expediated apoptosis and inflammatory cytokine release. These effects on cell behaviors suggested that ox-LDL induced endothelial dysfunction of HUVECs.
RT-qPCR has manifested that circ_0124644 level was aberrantly increased in AS samples and ox-LDL-exposed HUVECs. Dysregulation of circRNA expression is related to the progression of endothelial cell injury. Qin et al. stated that circ_0003645 downregulation abolished ox-LDL-induced endothelial cell inflammation and apoptosis [16]. Li et al. showed that upregulation of circ_0003575 suppressed proliferation and angiogenesis of ox-LDL-treated endothelial cells [17]. Liu et al. reported that circ_0003204 knockdown promoted endothelial cell proliferation but reduced apoptosis [18]. The previous study indicated that circ_0124644 can act as a diagnostic biomarker in coronary artery disease [7]. In addition, circ_0124644 contributed to the progression of acute myocardial infarction and ox-LDL-induced AS endothelial cell injury [8, 19]. Also, our results demonstrated that silence of circ_0124644 facilitated cell growth and tube formation ability while inhibited apoptosis and inflammation damages in ox-LDL-treated HUVECs. Circ_0124644 was validated to act as a molecular factor in the endothelial cell dysfunction. Thus, circ_0124644 can be used for clinical diagnosis of AS and AS progression in clinical patients can be delayed if circ_0124644 expression was knocked down.
CircRNAs can induce the inhibitory effect on miRNA activity by acting as natural sponges for miRNAs [20]. CircRNA-miRNA interaction has been indicated in the development of cardiovascular diseases [21]. Circ_0007623 contributed to cardiac repair by absorbing miR-297 [22]. Circ_0010729 protected against oxygen-glucose deprivation (OGD)-caused cardiomyocytic damages via targeting miR-145-5p [23]. CircRNA ciRS-7 enhanced tube formation and endothelial cell viability by sponging miR-26a-5p [24]. This study exhibited the binding between miR-370-3p and circ_0124644, and circ_0124644 downregulated the level of miR-370-3p. Furthermore, inhibition of miR-370-3p reversed the influences of si-circ_0124644 on cell behaviors after HUVECs were treated with ox-LDL. The circ_0124644 function in endothelial injury was attributed to function as a miR-370-3p sponge.
Moreover, miR-370-3p directly reduced the level of FOXO4 via targeting the 3’UTR of FOXO4. All protective effects of miR-370-3p against ox-LDL-induced damages were related to the downregulation of FOXO4. More importantly, circ_0124644 could elevate the level of FOXO4 via sponging miR-370-3p in ox-LDL-treated HUVECs. CircRNA/miRNA/mRNA regulatory networks have been discovered in regulation of endothelial injury, including circ_0006896/miR-1264/DNMT1 axis [25], circ-BANP/miR-370TXNIP axis [26], and circRSF1/miR-758/CCND2 axis [27]. In this research, circ_0124644/miR-370-3p/FOXO4 axis was first unraveled in ox-LDL-induced dysfunction of endothelial cells.
Conclusion
Taken together, circ_0124644 has aggravated vascular endothelial injury caused by ox-LDL in HUVECs through depending on miR-370-3p/FOXO4 axis (Fig. 7). These findings showed a novel miRNA/mRNA mechanism for circ_0124644 in ox-LDL-caused endothelial cell damages, which might contribute to circRNA research in AS.
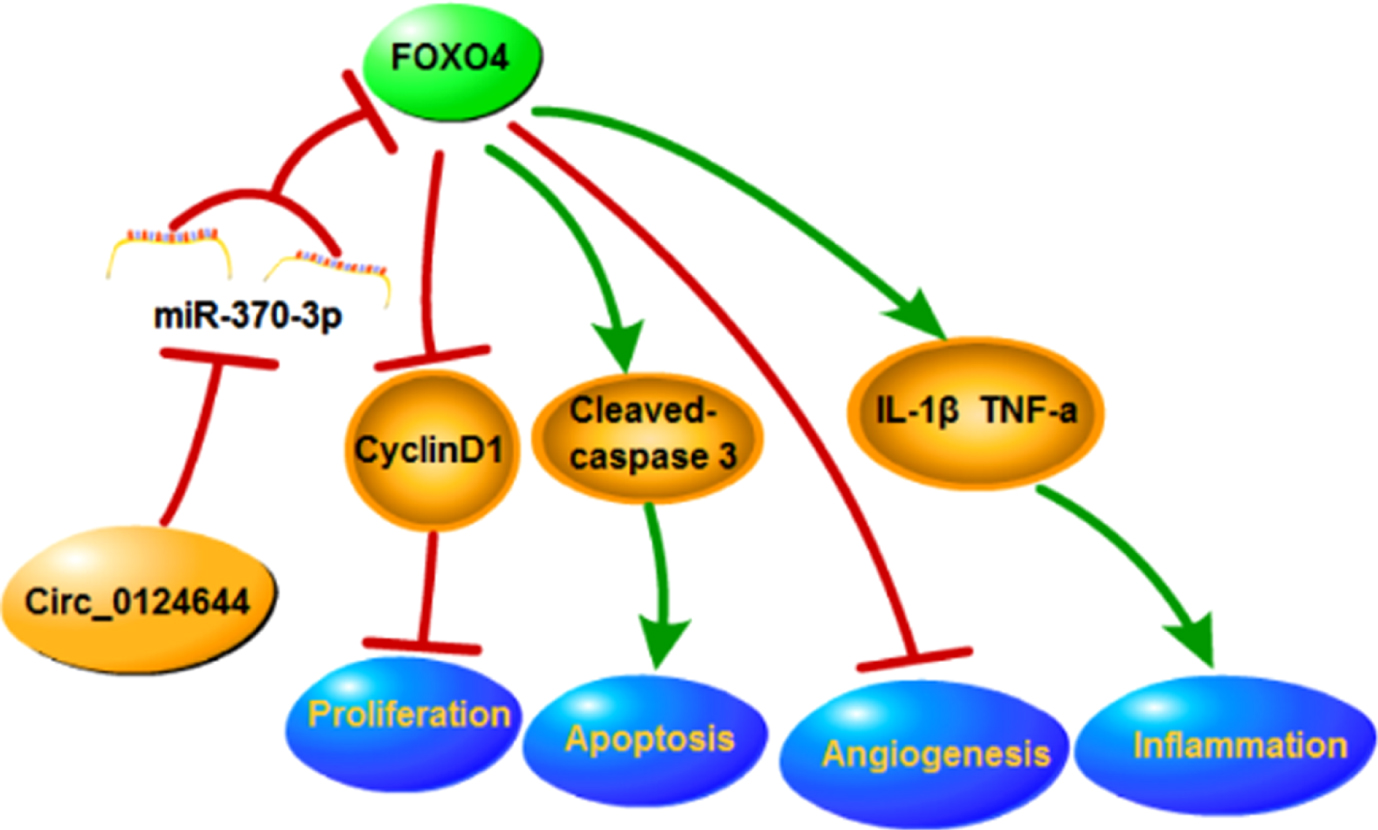
The graphical abstract of this research.
Footnotes
Acknowledgment
None.
Disclosure of interest
The authors declare that they have no conflicts of interest.
Funding
None