
Abstract
Blood microrheology depends on the constituents of blood plasma, the interaction between blood cells resulting in red blood cell (RBC) and platelets aggregation, and adhesion of RBC, platelets and leukocytes to vascular endothelium. The main plasma protein molecule –actuator of RBC aggregation is fibrinogen. In this paper the effect of interaction between the endothelium and RBC at different fibrinogen concentrations on the RBC microrheological properties was investigated in vitro. Laser tweezers were used to measure the RBC-endothelium interaction forces. It was shown for the first time that the interaction forces between RBC and endothelium are comparable with the RBC aggregation forces, they increase with fibrinogen concentration and reach the saturation level of about 4 pN at the concentration of 4 mg/ml. These results are important for better understanding the mechanisms of RBC and endothelium interaction and developing the novel therapeutic protocols of the microrheology correction in different pathologies.
Introduction
Blood microrheology and tissue perfusion depend on various factors, such as red blood cell (RBC) aggregation, interaction between blood cells, between RBC and vascular endothelium, deformation of RBC, etc. [1, 2]. In this work, two aspects affecting blood microrheology are considered, i.e., RBC aggregation and the interaction of RBC with vascular endothelium.
RBC aggregation is the reversible process of formation of coin-like stacked structures of RBC [1]. RBC aggregation mainly determines blood viscosity at low shear and, therefore, blood circulation in general [3–5]. For instance, it was shown that the RBC aggregation correlates with the blood velocity in cutaneous capillaries [6]. In in vivo conditions, the RBC aggregation causes the axial migration of RBC to the center of vessels, which leads to the formation of a cell-poor layer in the marginal flow zone [7]. Also, the axial migration of RBC displaces white blood cells and platelets to the marginal flow zone, which helps them to implement their functions [8, 9]. There are additional effects of RBC aggregation on blood circulation, leading to both an increase and a decrease in resistance to the blood flow [1]. There is evidence in favor of the concept of optimal RBC aggregation because it changes (mainly, arises) dramatically in many pathologies [3, 10–13]. It is well known that RBC aggregation can occur only in the solutions with high molecular weight molecules [14]. In these solutions, the aggregation of RBC can change due to many endogenous and exogenous factors, such as the alterations of blood plasma composition, temperature, RBC age, RBC deformability, the effect of gasotransmitters (e.g., nitric oxide), etc. [1, 16].
In blood plasma, the main inducer of RBC aggregation (as well as platelets aggregation) is fibrinogen [17, 18]. Previously, it was shown that RBC aggregation forces increase almost linearly with increasing the fibrinogen concentration [19, 20]. In general, an increased concentration of fibrinogen in the blood is one of the main factors for the pathological development of hemorheological (RBC aggregation, plasma viscosity) and vascular (increased blood flow resistance, increased blood pressure, etc.) events [21]. In case of many pathologies the elevated concentration of fibrinogen correlates with the enhanced RBC aggregation.
Changes in the structure of fibrinogen can also alter the aggregation properties of RBCs. For example, congenital dysfibrinogenemia is based on various changes in the structure of the fibrinogen molecule, leading to a variety of disorders in the process of blood coagulation [22]. It has been shown that dysfunction of the fibrinogen molecule has a more significant impact on the RBC aggregation compared to normal fibrinogen [23].
In addition, there is evidence that the degree of expression of the inducing or inhibitory role of macromolecules in RBC aggregation can be considered according to the hydrodynamic radius of the macromolecules [24]. Macromolecules with hydrodynamic radius below 4 nm inhibit RBC aggregation, while those with hydrodynamic radius higher than 4 nm are proaggregants. At present, the understanding of RBC aggregation mechanisms remains unclear and, in some respects, contradictory, i.e., well known “depletion” theory and the “bridging” model of RBC aggregation predict different effects [14, 25]. The interaction between RBC and endothelium is unclear as well for now: in several works the interaction between RBC and endothelium in pathological conditions is also explained by the “depletion” theory or the “bridging” model [26–28].
Endothelium is the layer of cells lining the inner surface of blood arteries, veins, and capillaries. Endothelial cells do not only form a semipermeable layer between blood and tissues, but also play an important role in hemostasis, in the regulation of vascular blood flow due to the atrombogenicity and thrombogenicity of the cell membrane under physiological conditions and regulation of the transcapillary exchange [29–31]. Different factors leading to the development of an inflammatory process and other pathological conditions change the anticoagulant state of endothelium into procoagulant one [32]. Since endothelial cells interact directly with blood cells, they can also affect microrheological properties such as RBC aggregation and deformability [33, 34]. Vascular endothelial cells can be a source of such signaling molecules for blood cells as gas transmitters [35]. Although RBCs do not adhere to the vascular endothelium in physiological conditions, in a number of pathological conditions, adhesion of RBCs to the vascular endothelium can be a leading factor to the formation of a microcirculatory obstruction, reduced perfusion and tissue ischemia [36]. To the best of our knowledge, so far, the RBC-endothelium interaction has been mostly studied under pathological conditions. For example, in the work [37] it was shown that the adhesion of RBC to the endothelium monolayer for “healthy” RBC is considerably lower compared to “sickle” RBC.
The main purpose of this study was to investigate the interaction between endothelium and RBC of healthy volunteers at different concentrations of fibrinogen (as the main promoter of RBC aggregation) at the stationary conditions at the level of individual cells in vitro. By varying the concentration of fibrinogen, it is possible to model the normal and pathological conditions.
Materials and methods
Laser tweezers
Laser tweezers are a scientific tool allowing to trap and manipulate single live cells and measure the interaction forces between them [38]. The schematic layout of the setup used in the experiments (i.e., home-made double-beam laser tweezers with fixed and movable trap) is presented in Fig. 1. The basic elements are the diode-pumped Nd:YAG laser (λ=1064 nm, maximum output power 1 W), and the water-immersion objective OLYMPUS with high numerical aperture (NA = 1.00). The wavelength of the laser is in the range of optical transparency of hemoglobin to minimize the heating effect of the laser trapping [39]. The optical transparency range of wavelengths of endothelium belongs to the same region [40].

The schematic layout of home-made laser tweezers used in the experiment.
Human umbilical vein endothelial cells (HUVEC) were isolated according to the methods of Jaffe [41] and Scheglovitova [42]. Umbilical cords were obtained after normal parturition from healthy donors following informed written consent. For the experiments, human umbilical vein endothelial cells (HUVEC) were grown on the cover glasses placed in 24-well plates in a CO2 incubator at 37 °C until a monolayer was formed. Endothelial cell growth medium (medium 199 (Gibco), gentamycin, heparin, 10% fetal calf serum (HyClone), endothelial growth factor (200μg/ml)) was used to culture endothelial cells. Only the cells of the first passage were used for the experiments. The morphology and function of endothelial cells in tissue varied noticeably from one passage to another, regardless of whether the cells were seeded on a polymer surface or glass [43, 44]. According to Ulfman et al., the expression of P-selectin decreased significantly in the second cell passage compared to the first passage [45]. On the other hand, the plasminogen-activator-inhibitor 1 increased, while the angiotensin-converting enzyme activity and its antigen expression decreased. Bala et al. found that endothelial cells started losing their characteristic morphology between the 3rd and 6th passages [46].
Blood for all experiments was drawn from the cubital veins of healthy male donors (n = 4) to obtain serum using tubes with SiO2 clot activator (5 ml) and whole blood with EDTA anticoagulant (4 ml). All volunteers were informed on the purpose of the study and gave written informed consent in accordance with the Declaration of Helsinki. The study was approved by the Ethics Committee of Medical Research and Educational Center of M.V. Lomonosov Moscow State University (protocol No. 11/22 05.12.2022). The sample to be placed in the cuvette consisted of serum with the addition of bovine fibrinogen (MDL number: MFCD00163624) at a certain concentration and a small amount of whole blood (1 part of blood to 1000 parts of serum with fibrinogen). In this study, we used bovine fibrinogen which is structurally close to the human fibrinogen. It was shown that there is no significant difference between human or bovine fibrinogen in endothelial adhesive properties [47]. In addition, as the evidence of bovine fibrinogen application instead of human, platelet aggregation is quite similar in the solutions with these both types of fibrinogens as inductors [48].
The following concentrations of fibrinogen in serum were used: 0, 2, 4, 6, 8 mg/ml. The sample was placed into the cuvette based on a glass slide, upon which a cover glass with endothelium monolayer was placed (see Fig. 1). A coverslip was then placed on top, and vacuum gel was used to isolate the cuvette edges from the air to prevent oxidative stress for endothelium. The sample and the cover glass with endothelium monolayer were placed into the cuvette a few minutes before the measurement. Between the preparations of the experimental cuvettes the endothelium was kept at 37°C with CO2 surrounding to prevent oxidative stress.
Endothelial cell viability was assessed by MTTrmbox1 []rmbox13-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide. reduction assay [49]. Briefly, confluent monolayer of endothelial cells in 96-well plates (15000 cells in 100μL) was incubated in a CO2 incubator for 24 hours, then endothelial cell growth medium (medium 199 (Gibco), gentamycin, heparin, 10% fetal calf serum (HyClone), endothelial growth factor (200μg/ml)) was removed, and blood serum diluted 1 : 1 in the medium was added. The medium was used as a control. After additional 24 hours of incubation, the MTT test was used to determine the functional activity of the cells. The optical density of solutions was determined using a spectrophotometer at 540 nm. There was no statistically significant decrease in the optical density of the serum sample between controls. It was shown that the serum is non-toxic for endothelium and even has an activating effect.
Measurements of the interaction forces
All measurements were performed under room temperature (22°C). In the experiments, the interaction forces between RBC and endothelium as well as the interaction forces between individual RBC were measured. Only the main (fixed beam) optical trap was used during force measurements between the RBC and the endothelium [50]. The force between RBC and endothelium is defined as the minimum force required to trap the adhered RBC and move it freely along the endothelium monolayer. Measurements of the RBC-endothelium forces were conducted on 7–10 pairs of the cells at each fibrinogen concentration and lasted approximately 30 min.
The interaction forces between individual RBC (aggregation and disaggregation) are the forces leading to the paired aggregation of RBC [10]. In particular, the aggregation force is the minimum force required to prevent the RBC aggregation process of two partly overlapped and sticked to each other cells, whilst the disaggregation force is the minimum force required to break the interaction of partly overlapped cells. Each force for each fibrinogen concentration was measured at least 5 times for the blood of each donor. Measurement with one RBC pair using laser tweezers took approximately 3-4 minutes, which does not harm the cells due to optical trapping [51]. Force calibration of the laser tweezers was performed using the Stokes force calibration method [10].
Statistical analysis
All data and all figures were processed using Origin 2022b software. Statistical differences of the samples were calculated using Mann–Whitney U test. Two samplings were considered statistically significantly different if the p-value was less than 0.05 (*p<0.05; **p<0.01; ***p<0.001; ****p<0.0001).
Results and discussion
Our measurements showed that with an increase in fibrinogen concentration in the serum, the interaction force between RBC and endothelium monotonously increases up to fibrinogen concentration of 4 mg/ml, and in the range from 4 to 8 mg/ml the interaction force reaches saturation (see Fig. 2a). It seems that saturation is reached when the concentration of fibrinogen is above the physiological limits. The physiological fibrinogen concertation lies in the range 2-4.5 mg/ml, with the median of about 3 mg/ml, and slightly differs for males and females [52]. In case of pathological and inflammatory processes in the body the concentration of fibrinogen is elevated, e.g., it was shown that mean fibrinogen values in patients with diabetic foot ulcer (grade ≥ 3) was about 5 mg/ml [53]. Based on these results, it can be concluded that fibrinogen concentration is not a leading factor in the adhesion of RBC to the endothelium at high fibrinogen concentrations for RBC from the blood of healthy volunteers and endothelium in physiological conditions. The effect of fibrinogen on RBC adhesion to the endothelium in the case of pathology requires additional studies. Most likely it will be different, because in pathology there is a significant increase in RBC adhesion to the endothelium [37]. We demonstrated the interaction between RBC and endothelium in serum at the fibrinogen concentration 0 mg/ml (see Fig. 2a) that implies presence of other factors inducing the interaction (adhesion). This may also imply that the interaction between RBCs and the endothelium is a result of synergistic effect of several plasma macromolecules, as does the RBC-RBC interaction [54]. The difference of RBC-endothelium interaction forces was quite high between donors. It may be caused by variations in the endothelium from different donors. Normalized interaction force (by force at the zero concentration of fibrinogen of each donor to reduce dispersion in the forces as factor of different endothelium cells) gives clearer dependence (see Fig. 2b).
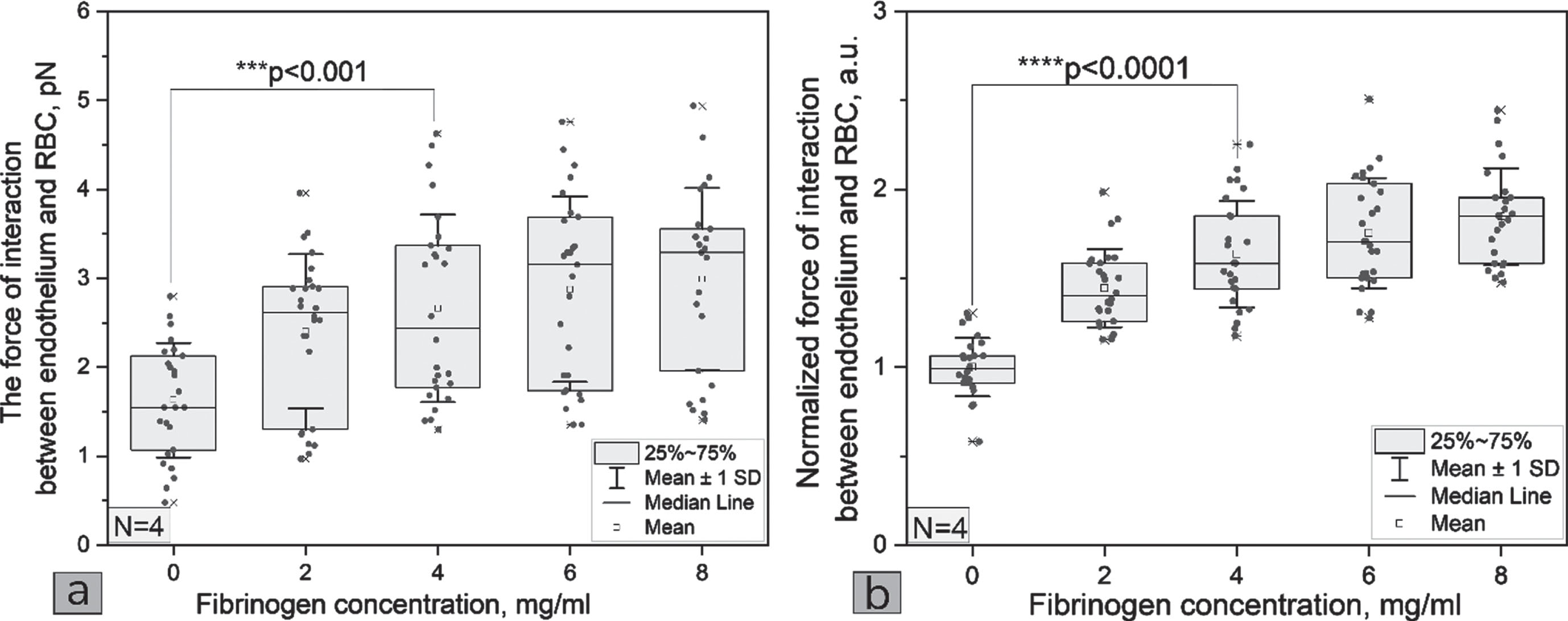
(a) The dependence of the interaction force between RBC and endothelium on the fibrinogen concentration in the blood serum and (b) the normalized (by the concentration 0 mg/ml for each donor) dependence of the interaction force between RBC and endothelium on the fibrinogen concentration in the blood serum. p-values are calculated by using Mann–Whitney U test. Each point represents one measurement with one of the samples of the four donors.
The expected results were also obtained that the RBC aggregation force as well as the RBC disaggregation force increases monotonously with increasing fibrinogen concentration (see Fig. 3ac). These results are in good agreement with our results obtained earlier for RBC in autologous blood serum [19]. Interestingly, the aggregation forces of RBC are correlated in magnitude (∼pN) with the interaction forces between RBC and endothelium. Compared to the RBC-endothelium normalized interaction forces (Fig. 2a), the normalized aggregation and disaggregation forces show a difference between donors (see Fig. 3bd).
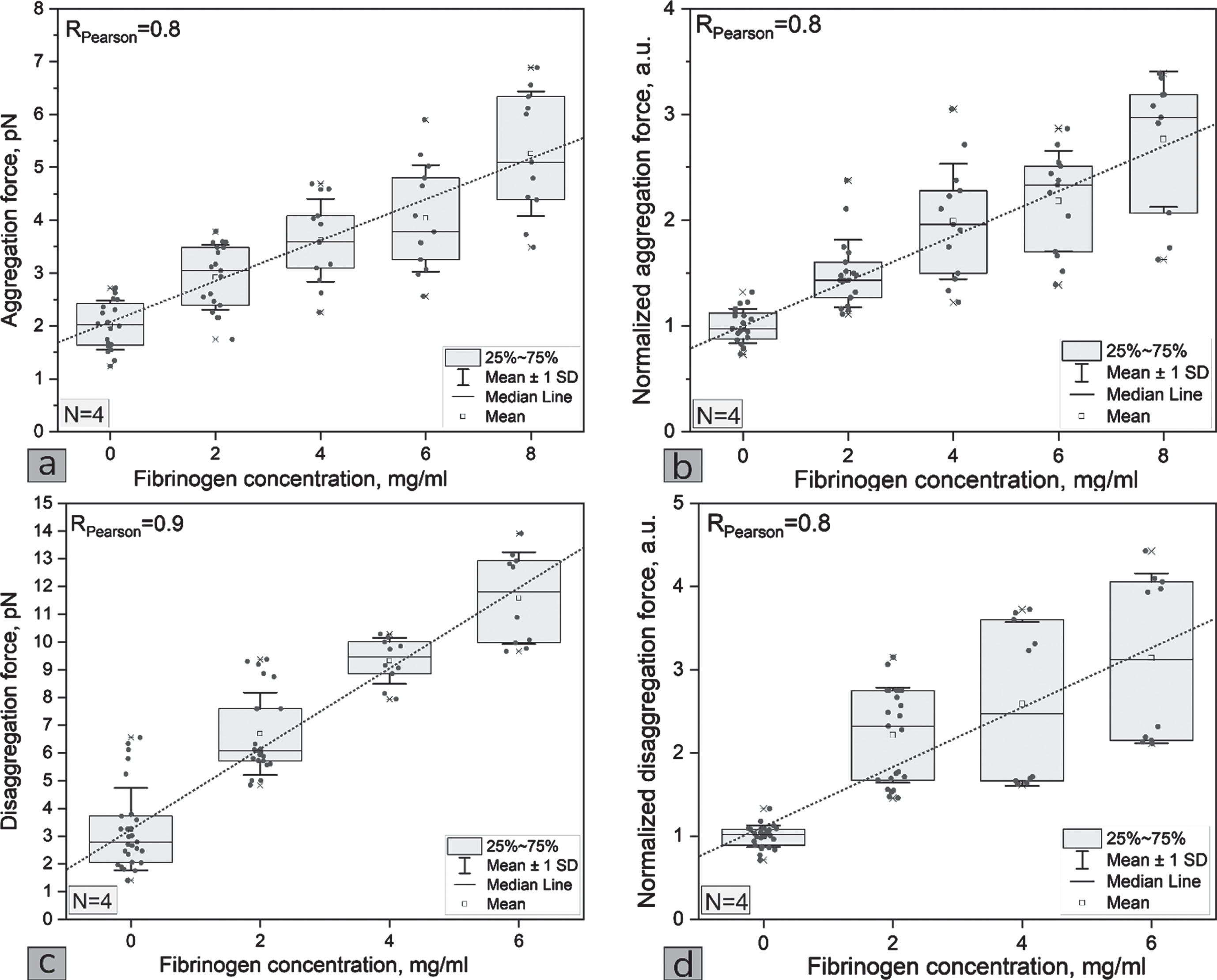
(a) The dependences of RBCs aggregation force, (b) the normalized (by the concentration 0 mg/ml for each donor) RBCs aggregation force, (c) disaggregation force and (d) the normalized (by the concentration 0 mg/ml for each donor) RBCs disaggregation force on the fibrinogen concentration in the blood serum. Each point represents one measurement with one of the four donor samples.
The forces between RBC and endothelium differ from the same forces in our previous work [50] for several reasons: (1) in this work, the serum was used as the medium rather than EDTA plasma; (2) monolayer of endothelial cells was used rather than single endothelial cells. In summary, the experimental design in this work is nearer to the in vivo conditions than in our previous work.
The force of interaction between RBC and endothelium in physiological conditions was studied previously, to the best of our knowledge, by Mohandas [55]. In this article, the measurements were conducted using the micropipette aspiration technique and the range of forces was around 2.2 nN. Since different methods of measuring forces between individual RBCs are based on different physical principles, the results may be quite different from each other and may differ from those obtained by other methods, for example, forces measured with laser tweezers are different from those measured with the micropipette aspiration or atomic force microscopy methods. The two latter ones are direct methods, in which red blood cells are subjected to a mechanical contact, and the RBC aggregation forces measured using these methods are in the nN range [56]. Since laser tweezers allow for trapping cells without mechanical contact, the range of measured forces better correspond to physiological conditions compared with that measured by micropipette aspiration or atomic force microscopy.
Using laser tweezers for in vitro measurements, it was found for the first time that with an increase in fibrinogen concentration, the interaction force between RBC and HUVEC endothelium increases to a fibrinogen concentration of 4 mg/ml, and in the range from 4 to 8 mg/ml, the interaction force reaches saturation. The study demonstrated that in the presence of endothelium, the forces of aggregation and disaggregation of RBC increase consistently with higher fibrinogen concentrations, similar to the observations made previously in the absence of endothelium. These results are important for a better understanding of the interaction between RBC and endothelium.
Footnotes
Acknowledgments
This work was supported by the Russian Science Foundation Grant No. 22-15-00120.