In this study, the safety and tolerability of new candidate HCV vaccine named Cenv6 were screened in mice. Cenv6 peptide is composed of 6 synthetic HCV peptides (3 structural and 3 nonstructural peptides).
METHODS:
Forty eight mice were enrolled in this study, 12 controls and 36 mice (the thirty-six mice were categorized into 3 groups according to administered doses (3 monthly doses of 800 ng, 1600 ng, and 16 g/25 gm mouse body weight (bw))). Hematological, biochemical and histopathological changes were appraised.
RESULTS:
Our data indicated that the doses of 800 ng and 1600 ng of Cenv6 per 25 gm mouse body weight were safe as compared to the dose 16 g/25 gm bw (10 times more than the potential therapeutic dose) for all examined tissues while the 16 g Cenv6 dose provoked histopathological changes in kidneys, liver and lungs.
CONCLUSIONS:
The extravagant histopathological changes in different organs have exiled the 16 g dose out of acceptable range and validated that Cenv6 is safe and tolerable at the two lower doses (800 and 1600 ng/25 gm bw).
Hepatitis C virus (HCV) infection does not confer lifelong immunity but evades from the immune system resulting in chronic infection, Hepatocellular carcinoma and death [8, 13, 20]. HCV is a significant public health problem in Egypt, with a 15% frequency rate that just about 11.5 million Egyptians are infected with HCV [10], dominantly genotype 4 [18, 20]. Direct acting antiviral agents (DAAs), heighten cure rates to around 90% without the want for interferon and potently treat the majority of HCV genotypes [15, 27]. Despite the strikingly success of DAAs, there are 3 to 4 million newly infected cases each year not only in the developing countries but also in the developed ones. Therefore, there is an immense demand for the production of a prophylactic vaccine [7, 8].
Previous investigations used synthetic peptides derived from HCV envelope proteins as vaccine candidates in an attempt to provoke neutralizing antibodies capable of blocking HCV entry and disrupting the programmed viral life cycle [11, 16]. We formerly reported on the vaccination strategy based on the E1 and E2 specific epitopes in immunized animals which prosperously elicited sturdy antibody response and neutralized HCV virions in vitro [8, 9]. HCV-specific memory cytotoxic CD4 and CD8 T-lymphocytes were known to endure for 20 years following viral clearance [3]. Accordingly, an idealistic HCV vaccine will need to provoke the production of neutralizing antibodies and enduring HCV-specific T-cell responses [3, 6, 21].
Herein, we introduced a vaccine candidate named Cenv6, composed of 3 synthetic HCV structural peptides, one derived from E1 and two from E2 proteins of HCV in addition to 3 synthetic peptides derived from nonstructural proteins NS4A, NS4B and NS5A. The peptides were selected on the basis of sequence data available for genotype 4a quasi-speciesand were aligned with all HCV subtypes and isolates recorded on the HCV database (Ref Seq NC_009825.1). The six peptides were synthesized in an 8 arm multiple antigenic peptide (MAP) form which displayed efficient immune response than linear peptides as reported previously [8, 19]. The six peptide sequences were derived from HCV genotype 4a EG.ED43.Y11604. The alignment was done by using Clustal W multiple sequence alignment programs at http://align.genome.jp/.
In this study our goal was to assess the safety and tolerability of Cenv6 in Balb-c female mice. We appraised the biochemical, hematological, and histopathological parameters in 12 mice as a control group and 36 mice receiving 3 monthly injections from each of the following doses 800 ng, 1600 ng and 16 g per 25 gm bw.
Experimental
Peptides
The synthesis of the six peptides was done commercially by AnaSpec, Inc, (San Jose CA, USA), in the amide form, using standard solid phase synthesis involving 9-flurenylmethoxy carbonyl chemistry and purified using HPLC as described previously [4].
Mice
Six to nine-week-old female mice (Balb c, weighing 18–20 g) were purchased and housed in the animal facility of the National Research Center. Ethical approval was obtained from the Institutional Animal Care and Use Committee of National Research Center, Egypt (Ethical approval no. 17112). Animal care and handling were performed according to the guidelines set by the World Health Organization (WHO), Geneva, Switzerland and the MREC (Medical Research Ethics Committee), Cairo, Egypt [1]. Mice were maintained in a temperature controlled environment at 24C with a 12 h light/dark cycle, and were provided with drinking water and feed ad libitum. All efforts were taken to minimize the suffering of animals used. The mice were acclimatized in the laboratory conditions for a period of 1 week before being used in the experiment under observation to exclude any intercurrent infection.
Three groups, each consists of 12 mice were categorized according to administered doses (800 ng, 1600 ng and 16 g per 25 gm bw) and 12 mice as a control group. Mice were immunized subcutaneously (S.C.) with 120 l (total volume) of peptides in a MAP form from each of the three mentioned doses.
Animals were bled through retro-orbital plexus/ sinus before each injection. The control group was treated exactly similar to test groups but injected with 120 l PBS. All experiments were conducted in accordance with the Institutional Animal Care and Use Committee (IACUC) approved protocol [28].
Blood sampling
Each mice was bled through retro-orbital plexus/ sinus before immunization and after 4, 8 and 12 weeks from each of the following doses 800 ng, 1600 ng and 16 g per 25 gm bw. One ml blood sample was collected from 3 mice. The blood samples were used for hematologic tests and for all the biochemical parameters. The collected sera were immediately stored at 70C until the time of analysis.
Hematological tests
To explore the hematological changes that might occur in response to doses of peptides, Complete blood count (CBC) including Red blood Cell counts (RBCs), hemoglobin concentrations (Hb), differential leucocytic counts and platelets counts were measured in all immunized mice groups as well as in control group.
Biochemical tests
To test changes in liver and kidney functions following different doses of Cenv6 injections, serum levels of alanine aminotransferase (ALT) and creatinine were measured in the mice pre immunization and after 4, 8 and 12 weeks using commercial kits (Abcam).
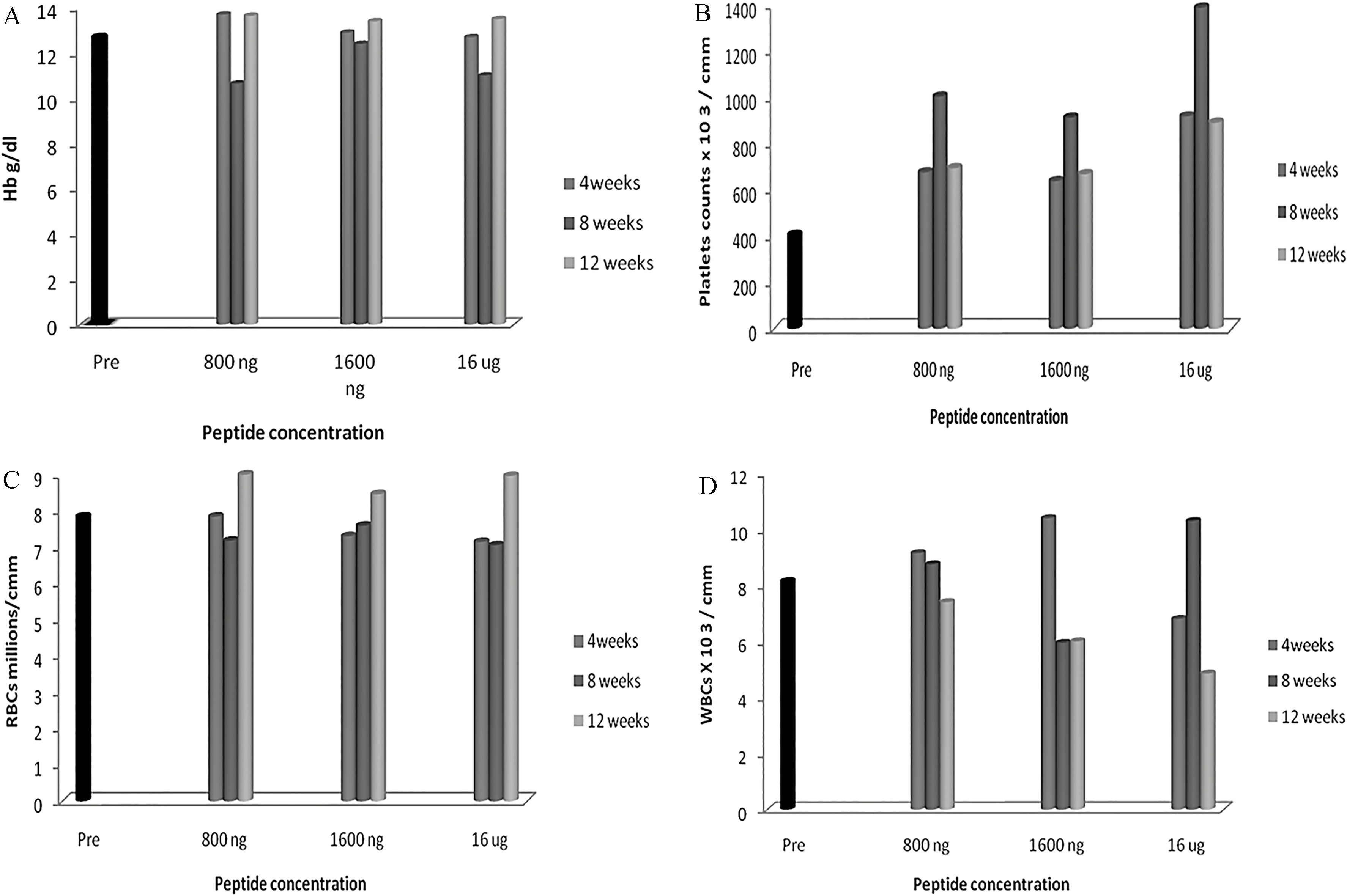
Hematological variations in blood cells of mice immunized with Cenv6 peptide. Three mice groups (12 mice per each group) were immunized subcutaneously with 3 different doses of Cenv6 vaccine (800 ng, 1600 ng, and 16 g) as well as 12 mice as a control group. Animals were sacrificed after 4, 8 and 12 weeks from immunization and 3 ml blood sample was taken. Hb of non immunized mice ( 12) were contrasted with Hb of immunized mice ( 36) (A), Platelets (B), RBCs (C) and WBCs (D).
Histopathological examination
The histopathological changes were visualized after mice immunization with 800 ng, 1600 ng and 16 g of Cenv6 per 25 gm bw (3 animals per group). Animals were sacrificed after 4, 8 and 12 weeks from immunization and samples from liver, bone marrow, spleen, kidneys, heart, lungs, ovaries and brain were obtained. In general, tissue samples were fixed in 7% buffered formalin (eyes in Davidson’s solution), embedded in paraffin, sections prepared and routinely stained with hematoxylin and eosin and observed under a light microscope (Leica DMI4000 B; Leica Microsystems, Wetzlar, Germany).
Results
Hematological changes
The results elaborated that the three injected doses of 800 ng, 1600 ng and 16 g of Cenv6 did not provoke changes in Hb concentrations as compared to control group (12.7 0 g/dL). Mean of Hb for mice immunized by 800 ng, 1600 ng and 16 g of Cenv6 by three repeated s.c. injections, each with a 4-week interval were (3.2 g/dL, 13.7 0–13.65 0), (3.2 g/dL, 12.9 0–13.4 9.8), and (3.1 g/dL, 12.7 0–13.5 0.7) respectively (see Fig. 1A and Table 1).
Hematological and biochemical parameters of immunized mice with different doses of Cenv6 vaccine
Dose
Sampling
Mice
HB
RBCs
WBCs
Platelets
ALT
Creatinine
(weeks post
number
g/dL
10
10
(cmm)
(IU/ml)
mg/dl
injection)
cells/L
cells/L
(mean SD)
(mean SD)
Non immunized mice
( 12)
12.7
0
7.84
0
8.15
7.07
405
7.07
60
6
0.39
0
800 ng ( 12)
4 weeks
3
13.7
0
7.84
0
7.3
0
675
147.1
70
10
0.49
0
8 weeks
3
10.65
0
7.18
0
7.59
0
100
48.1
73.3
11.5
0.67
0
12 weeks
3
13.65
0
9
0
8.46
0
693
55.2
42.6
6.4
0.64
0
Mean/12
3.2
2
2
198
15.49
0.15
1600 ng ( 12)
4 weeks
3
12.9
0
7.3
0
10.4
0
637.5
217
63.3
15.2
0.38
0
8 weeks
3
12.4
0
7.59
0
5.95
0
912
281
49.3
11.01
0.7
0
12 weeks
3
13.4
9.8
8.46
0
6
0
667
127
81.3
1.15
0.6
0
Mean/12
3.2
1.95
1.9
184
16.16
0.14
16 g ( 12)
4 weeks
3
12.7
0
7.2
0
6.8
0
917
222
96.6
18.9
0.44
0
8 weeks
3
11
0
7.05
0
10.3
0
1387
188
61
18.5
0.63
0
12 weeks
3
13.5
0.7
8.96
0
4.85
0
890
7.77
92.3
26.3
0.67
0
Mean/12
3.12
1.94
1.8
266.17
20.83
0.145
Hb: heamoglobin, RBCs: red blood cells, WBCs: white blood cells, PC: platelets count and ALT: alanine aminotransferase level.
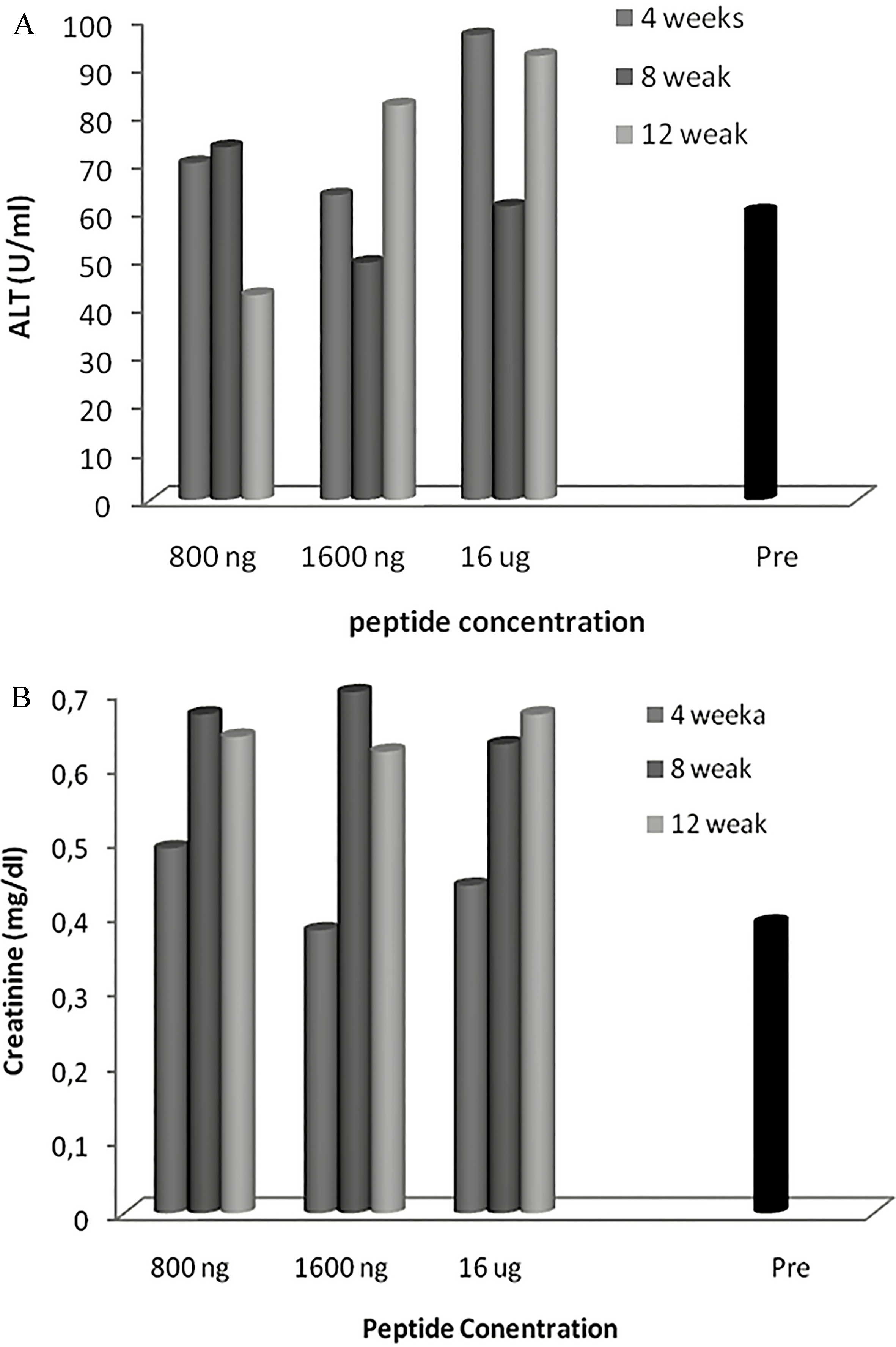
Biochemical variations in blood cells of mice immunized with Cenv6 peptide. Three mice groups (12 mice per each group) were immunized subcutaneously with 3 different doses of Cenv6 vaccine (800 ng, 1600 ng, and 16 g) as well as 12 mice as a control group. Animals were sacrificed after 4, 8 and 12 weeks from immunization and 3 ml blood sample was taken. ALT of non immunized mice ( 12) were contrasted with ALT of immunized mice ( 36) (A), Creatinine (B).
RBCs count subsisted in the level of control group (7.84 0 10 cells/L). Mean of RBCs count for mice immunized by 800 ng, 1600 ng and 16 g were (2 10 cells/L, 7.84 0–9 0) (1.95 10 cells/L, 7.3 0–8.64 0) and (1.94 10 cells/L, 7.2 0–8.96 0) respectively (see Fig. 1C and Table 1). WBCs count subsisted in the level of control group (7.84 0 10 cells/L). Mean of WBCs count for mice immunized by 800 ng, 1600 ng and 16 g were (2 10 cells/L, 7.3–8.46), (1.9 10 cells/L,10.4–6) and (1.8 10 cells/L, 6.8–4.85) respectively (see Fig. 1D and Table 1).
On the other hand the 16 g dose elaborated an increment of the platelets counts. Mean of the platelet count was (266.2 cmm, 917 222–890 7.77) than control group (405 7 cmm) while, the mean of platelets count at the doses 800 ng and 1600 ng were (198 cmm, 675 147.07–693 55.15) and 184 cmm, 637.5 217–667 127) respectively (see Fig. 1B and Table 1).
Biochemical parameters of immunized mice with different doses of Cenv6
The data presented in Table 1 showed that the mice immunized with the dose 800 ng of CENV6/25 gm bw showed a slight increment of serum ALT level (70 10 IU/ml, 73.3 11.5) at weeks 4 and 8 respectively contrasted with control group (60 6 IU/ml). Moreover at week 12, the ALT level returned back to the normal levels of non immunized mice. The dose 1600 ng of CENV6 per 25 gm mouse body didn’t show increase of serum ALT at weeks 4 and 8 (63.3 15.2 IU/ml) and (49.3 11.01 IU/ml) respectively. On the other hand, it was slightly elevated at week 12 (81.3 1.15 IU/ml). The dose 16 g of Cenv6 per 25 gm body weight of mice displayed a modest elevation of ALT (20.83 IU/ml, 96.6 18.9–92.3 26.3). Creatinine level remarkably elevated at week 8 and 12 for the three vaccine doses than control (0.39 0 mg/dl). Mean of Creatinine level for mice immunized by 800 ng and 1600 ng and 16 g were (0.15 mg/dl, 0.49–0.64), (0.14 mg/dl, 0.38–0.6) and (0.145 mg/dl, 0.44–0.67) respectively. See Fig. 2.
Histopathology
The histopathological changes were screened following injection of 800 ng, 1600 ng and 16 g of Cenv6 candidate vaccine per 25 gm bw of mice (3 animals per group). Animals were sacrificed after 4, 8 and 12 weeks from immunization. The data shown in Table 2 indicated that the doses of 800 ng and 1600 ng of Cenv6 per 25 gm mouse body weight were safe as compared to the control group for all examined tissues. On the contrary, the 16 g Cenv6 dose (10 times more than the potential therapeutic dose) provoked histopathological changes in kidneys, liver and lungs see Table 2. Further studies should be carried out at the molecular pathology level to survey the intracellular changes post immunization.
Severity score shown by histopathological changes of immunized mice with different doses of Cenv6 vaccine
Severity score
Dose
Severity
16 g
6 + ve
More severe
1600 ng
++ ve
Nearly the same
800 ng
+ ve
Less severe
Discussion
This study evaluated the safety and tolerability of Cenv6 peptides in immunized mice to produce an affordable and tolerable HCV vaccine. Our investigations confirmed that Cenv6 is safe and tolerable at a couple of doses (800 and 1600 ng/25 gm bw) by three repeated s.c. injections, each with a 4-week interval.
Upon the immunopathogenesis of HCV infection, the idealistic HCV vaccine must gather two characteristics: It should have the potency to provoke cross neutralizing antibodies along with persistent and potent HCV-specific T-cell responses. These requirements can be met with structural peptides derived from the envelope proteins to possess B cells in addition to nonstructural peptides to trigger the HCV-specific CD4 T cell response [14, 21, 26]. Moreover, it was shown that HCV-specific memory CD4 and CD8 T cells persist for approximately 20 years after viral clearance among humans [23] and also up to 7 years among chimpanzees [2, 22]. In this study, we introduced synthetic vaccine CENV6 that comprises of 6 peptides, three peptides derived from the nonstructural regions NS4A, NS4B and NS5A which are formerly proved to be targets of cellular immune responses in patients [16]. The other three peptides are derived from E1 and E2 proteins of HCV in order to provoke humoral immune responses.
Our hematological investigations in mice receiving the three doses did not reveal remarkable changes except for platelets. Platelets count elaborated gradual but transient increment especially in mice received the 16 g dose. The noticed changes in platelets count were deemed to be a physiological responses to inflammation [8, 12].
ALT and Creatinine are unique enzymes act as indicators for liver and kidney functions. While the liver and kidney encounter any damage or inflammation, these indicators will fluctuate significantly [7, 24]. In this study, ALT subsisted within common ranges in mice receiving 800 ng and 1600 ng of Cenv6. Later, Creatinine level was hiked gradually with the dose increment. On the contrary mice received 16 g of cenv6/25 gm bw revealed a modest elevation of ALT and Creatinine. The elevated levels of ALT and Creatinine are more likely to be transient [7, 17]. We conclude that single or repeated injections of a couple of doses (800 and 1600 ng/25 gm bw) did not reveal significant changes in the mentioned parameters and so did not cause any injury to liver and kidney functions.
The histopathological investigations indicated that both doses of 800 ng and 1600 ng of Cenv6 peptide per 25 gm mouse bw were safe for all examined tissues. Otherwise, the observed histopathological changes in liver, lung, heart, spleen and kidneys have deported the 16 g dose out of acceptable range. This explicit toxicological dose (16 g/25 gm bw) is 10 times more than the proposed therapeutic dose thus extending the safety margin. No local intolerance reactions related to the vaccine were seen. There were no vaccine-related toxic effects observed either. Our results further confirm the findings of former investigators reporting excellent preclinical safety profiles of synthetic peptide vaccines designed with HCV antigen epitopes [8, 16, 25]. It was particularly strikingly to find IFN- responses occurred in considerable number of mice in ELIspot assays (own unpublished data).
Conclusion
The extravagant histopathological changes in different organs have exiled the 16 g dose out of acceptable range and validated that Cenv6 is safe and tolerable at the two lower doses (800 and 1600 ng/ 25 gm bw). However the intracellular changes post vaccination still require further studies, we conclude that Cenv6 has a favorable safety profile that supports the initiation of testing its immunogenicity and human clinical trials.
References
1.
Abdel-AalW.GhaffarE.A. and El ShabrawyO., Review of the Medical Research Ethics Committee (MREC), National Research Center of Egypt, 2003–2011, Curr. Med. Res. Opin29 (2013), 1411–1417.
2.
AbdelwahabK.S. and SaidZ.N.A., Status of hepatitis C virus vaccination: Recent update, World J. Gastroenterol22 (2016), 862–873.
3.
AbdelwahabS.F., Cellular immune response to hepatitis-C-virus in subjects without viremia or seroconversion: Is it important? Infect. Agent. Cancer11 (2016), 23.
4.
El AwadyM.K.EI-DemellawyM.A.KhalilS.B.GalalD. and GoueliS.A., Synthetic peptide-based immunoassay as a supplemental test for HCV infection, Clin. Chim. Acta325 (2002), 39–46.
5.
CanzaterS. and CrowleyJ.S., About this report hepatitis policy project, 2017.
6.
CashmanS.B.MarsdenB.D. and DustinL.B., The humoral immune response to HCV: Understanding is key to vaccine development, Front. Immunol5 (2014), 550.
7.
DufourD.R.LottJ.A.NolteF.S.GretchD.R.KoffR.S. and SeeffL.B., Diagnosis and Monitoring of Hepatic Injury. II. Recommendations for Use of Laboratory Tests in Screening, Diagnosis, and Monitoring (n.d.).
8.
El-AwadyM.K.a TabllA.El-AbdY.S.YousifH.HegabM.RedaM.El ShenawyR.MoustafaR.I.DegheidyN. and El DinN.G.B., Conserved peptides within the E2 region of hepatitis C virus induce humoral and cellular responses in goats, Virol. J6 (2009), 66.
9.
El-awadyM.K.TabllA.A.YousifH.El-abdY.RedaM.KhalilS.B.El-zayadiA.R.ShakerM.H.BaderN.G. and DinE., Murine neutralizing antibody response and toxicity to synthetic peptides derived from E1 and E2 proteins of …, Vaccine, 2014.
10.
ElgharablyA.GomaaA.I.CrosseyM.M.NorsworthyP.J.WakedI. and Taylor-RobinsonS.D., Hepatitis C in Egypt – past, present, and future, Int. J. Gen. Med10 (2017), 1–6.
11.
FirbasC.JilmaB.TauberE.BuergerV.JelovcanS.LingnauK.BuschleM.FrischJ. and KladeC.S., Immunogenicity and safety of a novel therapeutic hepatitis C virus (HCV) peptide vaccine: A randomized, placebo controlled trial for dose optimization in 128 healthy subjects, Vaccine24 (2006), 4343–4353.
12.
FomsgaardA.KarlssonI.GramG.SchouC.TangS.BangP.KromannI.AndersenP. and AndreasenL.V., Development and preclinical safety evaluation of a new therapeutic HIV-1 vaccine based on 18 T-cell minimal epitope peptides applying a novel cationic adjuvant CAF01, Vaccine29 (2011), 7067–7074.
13.
IbrahimM.K.SalumG.M.Bader El DinN.G.DawoodR.M.BarakatA.KhairyA. and El AwadyM.K., Transcriptional dysregulation of upstream signaling of IFN pathway in chronic HCV type 4 induced liver fibrosis, PLoS One11 (2016), 1–14.
14.
JanvierG.ChaixM.L.FontaineH.BressonJ.L.PolS. and RiviereY., The core-specific precursor T cell response is directed to the N-terminal and central parts of the protein and positively correlates to the viral load in chronically HCV-infected patients, Virology340 (2005), 318–325.
15.
El KassasM.FunkA.L.SalaheldinM.ShimakawaY.EltabbakhM.JeanK.El TahanA.SweedyA.T.AfifyS.YoussefN.F.EsmatG. and FontanetA., Increased recurrence rates of hepatocellular carcinoma after DAA therapy in a hepatitis C-infected Egyptian cohort: A comparative analysis, J. Viral Hepat25 (2018).
16.
KladeC.S.WedemeyerH.BergT.HinrichsenH.CholewinskaG.ZeuzemS.BlumH.BuschleM.JelovcanS.BuergerV.TauberE.FrischJ. and MannsM.P., Therapeutic vaccination of chronic hepatitis C nonresponder patients with the peptide vaccine IC41, Gastroenterology134 (2008), 1385–1395.
17.
KundrotasL.W. and ClementD.J., Serum alanine aminotransferase (ALT) elevation in asymptomatic US Air Force basic trainee blood donors, Dig. Dis. Sci38 (1993), 2145–2150.
18.
MohamoudY.A.MumtazG.R.RiomeS.MillerD. and Abu-RaddadL.J., The epidemiology of hepatitis C virus in Egypt: A systematic review and data synthesis, BMC Infect. Dis13 (2013), 288.
19.
PosnettzD.N.McgrathH. and TamP., A novel method for producing anti-peptide antibodies, Biochemistry263 (1988), 1719–1725.
20.
SalumG.M.Bader El DinN.G.IbrahimM.K.AnanyM.A.DawoodR.M.KhairyA. and El AwadyM.K., Vascular endothelial growth factor expression in hepatitis C virus-induced liver fibrosis: A potential biomarker, J. Interf. Cytokine Res37 (2017), 310–316.
21.
Schulze zur WieschJ.LauerG.M.DayC.L.KimA.Y.OuchiK.DuncanJ.E.WurcelA.G.TimmJ.JonesA.M.MotheB.AllenT.M.McGovernB.Lewis-XimenezL.SidneyJ.SetteA.ChungR.T. and WalkerB.D., Broad repertoire of the CD4+ Th cell response in spontaneously controlled hepatitis C virus infection includes dominant and highly promiscuous epitopes, J. Immunol175 (2005), 3603–3613.
22.
ShoukryN.H.GrakouiA.HoughtonM.ChienD.Y.GhrayebJ.ReimannK.A. and WalkerC.M., Memory CD8 + T cells are required for protection from persistent hepatitis C virus infection, J. Exp. Med197 (2003), 1645–1655.
23.
Takakia.WieseM.MaertensG.DeplaE.SeifertU.Liebetraua.MillerJ.L.MannsM.P. and RehermannB., Cellular immune responses persist and humoral responses decrease two decades after recovery from a single-source outbreak of hepatitis C, Nat. Med6 (2000), 578–582.
24.
WangY. and ChenD., Toxicity assessment of repeated intravenous injections of arginine – glycine – aspartic acid peptide conjugated CdSeTe/ZnS quantum dots in mice, Int. J. Nanomedicine9 (2014), 4809–4817.
25.
WiegandJ.CornbergM.AslanN.SchlaphoffV.SarrazinC.KubitschkeA.BuggischP.CinerA.JaeckelE.MannsM.P. and WedemeyerH., Fate and function of hepatitis-C-virus-specific T-cells during peginterferon-alpha2b therapy for acute hepatitis C, Antivir Ther12 (2007), 303–316.
26.
YusimK.RichardsonR.TaoN.DalwaniA.AgrawalA.SzingerJ.FunkhouserR.KorberB. and KuikenC., Los alamos hepatitis C immunology database, Appl. Bioinformatics4 (2005), 217–225.
27.
ZhangX., Direct anti-HCV agents, Acta Pharm. Sin. B6 (2016), 26–31.
28.
Institutional Animal Care and Use Committee Procedure Guidebook Position Statement (n.d.).