
Abstract
Alzheimer’s disease (AD) is a neurodegenerative disease characterized by a progressive cognitive impairment of patients, affecting around 12% of people older than 65 years old. WHO estimated that over 48.6 million all over the world suffer this disease. On the basis of cumulative results on our research, we have postulated the neuroimmunomodulation hypothesis that appears to provide a reasonable explanation of both the preclinical and clinical observations. In this context, the long-term activation of the innate immune system triggers an anomalous cascade of molecular signals, finally leading to tau oligomerization in the pathway to neuronal degeneration. In the present scenario of the failure of many anti-AD drugs, nutraceutical compounds provide an avenue for AD prevention and possibly as coadjuvants in the treatment of this disease. Recent discoveries point to the relevance of curcumin, a natural anti-inflammatory agent, in controlling oxidative stress and improving cholinergic function in the brain, even though the mechanisms underlying these actions are unknown. We investigated the effects of curcumin in cultures of neuronal cells. For this study, we exposed cells to prooxidant conditions, both in the presence and absence of curcumin. Our data reveal that curcumin exert a strong neuroprotective effect in N2a cells, thus preventing toxicity by oxidative agents H2O2 and Fe+3. This is supported by results that indicate that curcumin control the neurodegenerative effects of both oxidative agents, relieving cells from the loss of neuritogenic processes induced by prooxidants. In addition, curcumin was able to slow down the tau aggregation curve and disassemble tau pathological oligomeric structures. Data suggest that curcumin could be a potential compound for prevention of cognitive disorders associated with AD.
INTRODUCTION
During the last few years, there have been several studies on a natural compound denominated curcumin (diferuloylmethane, Fig. 1), principal component derived from extracts of rhizomes of Curcuma longa; a plant that is found widely distributed in India. The principal consumption of extracts from rhizomes underlies the culinary uses and in the traditional medicine in the Oriental world. Curcumin is a polyphenolic and hydrophobic compound which has evidenced to possess various properties of biological benefit, among which is the antioxidant action, among other examples, as scavenger of superoxide, radical hydroxide and hydrogen peroxide, and as strong iron chelator [1, 2]. Curcumin is an anti-inflammatory agent, acting in the inhibition of induction of: (1) cyclooxygenase-2, (2) lipoxygenase, (3) inducible nitric oxide synthase, (4) production of cytokines and tumor necrosis factor, and (5) activation of transcription factors, such as NF-κB [3]. Moreover, an anti-aggregation effect of curcumin on oligomers of Aβ protein [4] has been observed. The properties of curcumin have led to postulate its therapeutic potential for several illnesses, such as cancer, diabetes, and neurodegenerative diseases [5].

N2a and HEK293 cells’ viability is slightly affected in a wide range of concentrations of curcumin. (A) Stoichiometric structures of Curcumin. Enol form and Keto form. (B) Concentrations from 1.0 μM to 15.0 μM of curcumin tested in N2a cells. All concentrations affected viability of N2a cells at minor levels, only 15.0 μM of curcumin was able to reduce viability below 20–25%, as DMSO (values represent the standard deviation. ANOVA test, factor followed by Dunnet’s test. *p < 0.05, **p < 0.01, ***p < 0.001). (C) Human embryonic kidney cells’ viability is slightly affected in a wide range of concentrations of curcumin. Concentrations from 1.0 μM to 15.0 μM of curcumin were tested in HEK293 cells. Only the highest concentration of curcumin affected viability of HEK293 cells. Values represent the standard deviation. ANOVA test, factor followed by Dunnet’s test. *p < 0.05, **p < 0.01, ***p < 0.001.
Alzheimer’s disease (AD) is the main cause of dementia throughout the world. Areas of the brain directly involved in learning, memory, and emotional behavior are primarily affected by the disease [6]. The prevalence of neurodegenerative disorders of the brain is over 10,000 cases per million inhabitants, and their projected increase is a matter of great concern, considering the growth rate in the elderly population. There is a serious public health crisis derived from dementia worldwide.
This disease is a neurodegenerative disorder, characterized by the formation of intracellular neurofibrillary tangles (NFTs) and extracellular deposits of amyloid-beta peptide (Aβ) in plaques, as well as microglial and astroglial activation, and finally, leading to neuronal dysfunction and death [7]. NFTs are composed of aggregates of hyperphosphorylated tau assembled in a paired helical filament (PHF) formation [8]. Phosphorylated tau, but not the unphosphorylated counterparts incorporates after oxidative stress reactions into pathological aggregates [9]. NFT and PHF deposition causes loss of synaptic function and finally neuronal death [10]. As a result of neurodegeneration tau aggregates are released to the extracellular domain, and evidence exists of their neurotoxicity [11, 12]. Thus, tau oligomers and PHF-tau appear to be neurotoxic agents, and several mechanisms have been suggested for its role associated to neurodegeneration. In addition, we must consider that tau filaments can stimulate the neuroinflammatory cascade [13].
Neuroinflammation is a pathological event that seems to be involved at the initial phase of several neurodegenerative disorders. Thus, inflammatory processes are a major feature of AD pathophysiology [14].
After suffering an injury, microglial cells trigger an immediate inflammatory response in defense to noxious stimuli, which leads to the repair of the site of injury. However, if the stimulus remains steadily in time, a permanent inflammatory condition develops, a phenomenon causing cumulative injury over time due to chronic inflammatory reaction [15]. These events preceding neuronal degeneration may also cause complex interactions and feedback loops between glial and neuronal cells, leading to cell damage and the development of a neurodegenerative disease [14]. Thus, this phenomenon should be seen as neuroinflammatory considering its effects to brain microenvironment results.
In the context of the neuroimmunomodulation theory, in AD, the liberation of pro-inflammatory factors (chemokines, cytokines, oxidative species, among others) from an activated glial cell (as astrocytes and microglia) induced tau hyperphosphorylation in neurons. This leads to the formation of tau aggregates, which during neurodegeneration are then released toward extracellular medium with the consequent neuronal death. The aggregated tau protein has pro-inflammatory effects, glial activation (triggering pro-inflammatory factor liberation) and, as we said, induction of the formation of news tau aggregators in surrounding neurons, with a subsequent neuronal death [13 , 16]. According to what we have shown so far, curcumin has a therapeutic potential on AD, regulating the effects produced by pro-inflammatory factors, in neuronal cells and above all in the regulation of the consequences produced by tau aggregation, where curcumin can act as anti-aggregating agent. An effect of curcumin at this level is controlling the inflammatory process involved after microglial activation.
We investigated the diverse properties of curcumin in the mouse cell line neuroblastoma Neuro-2a (N2a), as a cellular model for neurodegeneration of AD type. In order to assess curcumin effects on another type of cells, we used HEK293 kidney cells line. We assayed antioxidant properties of curcumin, inducing oxidative stress by H2O2 and neuroprotective effect through induction of damage with Fe+3; verifying the antioxidant and neuroprotective effect of curcumin by immunofluorescence studies. Moreover, we analyzed the effect of curcumin in the aggregates formation of recombinant protein hTau40 human by thioflavin S (ThS) assay, finding disaggregation effects of curcumin.
MATERIAL AND METHODS
Materials
Curcumin was purchased from Sigma-Aldrich (code: C7727). For its use in our experiments, curcumin powder was dissolved in DMSO, and subsequently serial dilutions were made in water to rule out the effect of the organic solvent on cells. Heparin and MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) were purchased from Calbiochem, Methylene blue and Thioflavine S were obtained from Sigma-Aldrich.
Mouse neuroblastoma N2a and human kidney HEK293 cell lines
Neuroblastoma N2a and HEK293 cells obtained after the third passage were used. N2a cells were cultured in a DMEM full medium supplemented with 5% fetal bovine serum (Gibico) at 37°C in 5% CO2. 24 h after the seeding, cultured cells were treated with different concentrations of curcumin (1–15 μM), in the presence or the absence of either 50 μM H2O2 or 80 μM Fe-NTA, according with the experimental protocol, and incubated for 24 h at 37°C with 5% CO2.
Viability assay
N2a and HEK293 cells were seeded in 96-well plates with a confluence of 7000 cells/well. The next day, cells were treated with variable concentration of curcumin (1–15 μM) for 24 h at 37°C and 5% CO2. Control groups were: (1) cells without the curcumin treatment and (2) a blank control, in which dish wells contained the medium but no cells. We used the MTT assay for cell toxicity. In each well, the medium was replaced by fresh DMEM medium without phenol red, 10 μl of MTT [12 μM] were added and incubated for 4 h at 37°C and 5% CO2. 100 μl of 5% SDS-HCl solution was added and incubated in the same conditions for 4 h and finally recording the absorbance at 550 nm using Sunrise plate reader.
Immunocytochemistry
N2a cells were plated on glass coverslips (12 mm diameter). After being exposed to experimental conditions, cells were washed with PBS and fixed with paraformaldehyde-sucrose (4%) for 30 min at 37°C. Cells were blocked with 5% BSA in PBS and then incubated with alpha-tubulin 1:1500 (Invitrogen), TO-PRO 1:500 (Life Technologies) and Alexa Fluor-488 phalloidin at 1:300 (Invitrogen). Then, slides were mounted in fluorescence mounting media (Calbiochem) and images were collected with a Carl-Zeiss Laser Scanning Confocal Microscopy. The photographs obtained were representative of the different samples.
Tau protein purification
The fragment of human full-length tau (htau40), the long isoform, was cloned into pET-28a vector (Novagen) to produce a His-tagged protein and htau40 was expressed and purified in Escherichia coli bacterial cultures as previously described [17]. This purified recombinant tau can self-assemble in the presence of heparin or arachidonic acid and resulted in an appropriate system to simulate pathological oligomerizations and evaluate the actions to protect molecules, such as curcumin and other natural compounds. The purity of proteins was verified on a Coomassie Brilliant Blue stained SDS-PAGE. Proteins were concentrated and stored at –20°C until use. The concentration of purified htau40 was determinate using the extinction coefficient at 280 nm (7700 M-1 cm-1).
Tau aggregation assays
Aggregation of htau40 for the experiments was made by incubating htau40 in the concentration of 50 μM in a final volume of 1 ml at 37°C in buffer PBS, pH 7.4 with the anionic cofactor heparin (molar ratio of tau to heparin = 4:1) for an incubation time of 8 days, with continuous shaking. In the case of control, the protein was incubated at the same concentration without the aggregating agent (heparin). On the third day of tau aggregation, curcumin was added to a final concentration of 10 μM. In control experiments, curcumin was also added at zero time.
ThS fluorescence assay
The htau40 aggregation was monitored by fluorescence Thioflavine S (ThS), as described by Friedhoff [18] and Barghorn [19]. The protein aggregation of a solution of the human htau40 variant by the action of heparin was carried out with 6.25 μM ThS. This was added to the solution and incubated for 1 h at 37°C in the dark, stirring in order to allow binding of the fluorescent ThS probe. Tau structures with a beta-sheet conformation were formed in the aggregates. Fluorescence was measured on the computer Biotek Synergy 2 excitation at 440 nm and emission at 508 nm (excitation filter 440/30 and 508/20 emission filter), minus the basal fluorescence of blank incubated in the same conditions.
Electron microscopy
The aggregation of 50 μM of htau40 protein for 8 days, according with tau assembly protocols with heparin, was carried out. The samples of htau40 were incubated in the absence of heparin, with heparin 12.5 μM or heparin 12.5 μM supplemented with 10 μM of curcumin. The micrographs were collected with a Philips Tecnai 12 Biotwin Transmission Electronic Microscope.
Processing and analysis of confocal images
The processing and analysis of confocal images were carried out in FIJI software. Statistical analyses were developed with a PRISM software.
RESULTS
Effect of curcumin on the cell viability of N2a
The first experiment was to identify the effect of curcumin on N2a cell viability, for which we used the colorimetric MTT assay. This assay is based on quantifying mitochondrial activity as a measure of cell viability and proliferation. This is how cells are metabolically active and thus the signal generated is directly proportional to the number of viable cells detected. The results using the MTT assay indicated that the percentage of N2a cell viability was slightly affected, with a decrease of around 20–25%, after incubating cells with curcumin at different concentrations (Fig. 1B). This suggests the lack of toxicity of this natural compound. With this result we were able to identify a range of concentrations of curcumin to assess their neuroprotective effect. It was important to evaluate comparatively the effects of curcumin on cell viability of another type of non-neuronal cells. We used the cell line HEK293 derived from the kidney. Only the highest concentration of curcumin affected viability of cells (Fig. 1C).
Curcumin neuroprotective effect against oxidative stress stimulus such as redox iron (Fe-NTA) and H2 O 2
To evaluate the neuroprotective effect of curcumin against two oxidative stress-inducing agents, N2a cell cultures were exposed simultaneously to Fe-NTA and curcumin, as well as H2O2 and curcumin. Cells were incubated with the following treatments: (1) Fe-NTA; (2) Fe-NTA and curcumin; (3) H2O2; (4) H2O2 and curcumin; (5) curcumin alone; and (6) untreated as a control. In the results we can see that the cell cultures when exposed to a stimulus of oxidative stress (Fig. 2D and 3D, and Supplementary Figure 1), as compared to a control experiment (Fig. 2A, 2G and 3A, 3G), tend to present an abnormal appearance, with signs of withdrawal in its cellular processes and short cellular processes (Fig. 3G and 4G) and decreased cell bodies, prior to the onset of cell death step. However, cells that have been treated only with curcumin did not show morphological alterations as compared to the control situation, and may even observe an increase in the extension of its cellular processes (Fig. 2B, 2C and 3B, 3C).

Effect of curcumin neuroprotective against oxidative stress in the presence of Fe-NTA. N2a cells after 24 h of treatment with curcumin and Fe-NTA. Untreated control condition (A), 10 μM curcumin (B), 20 μM curcumin (C), 80 μM Fe-NTA (D), 80 μM Fe-NTA and curcumin 5 μM (E), and Fe-NTA 80 μM and 15 μM (F). Red, α-Tubulin; blue: TOPRO (bar represent 50 μm). (G) Quantification of the length of the cellular processes (Values represent the standard deviation. ANOVA test, factor followed by Dunnet’s test. *p < 0.05, **p < 0.0002).
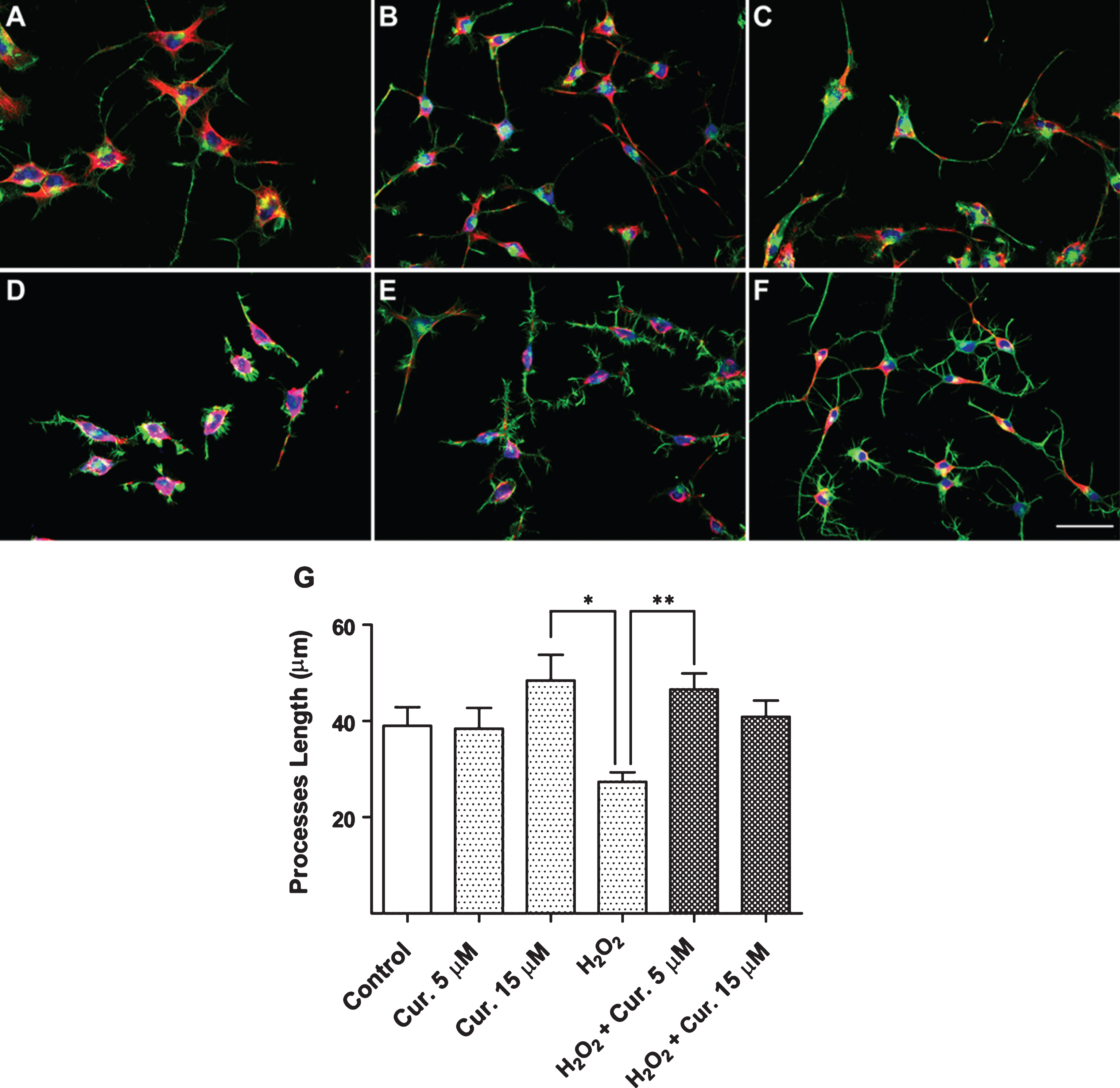
Neuroprotective effect of curcumin against oxidative stress in the presence of H2O2. Untreated control condition (A), Curcumin 5 μM (B), Curcumin 15 μM (C), H2O2 50 μM (D), H2O2 50 μM and 5 μM Curcumin (E), and H2O2 50 μM and Curcumin 15 μM (F). Red, α-Tubulin; Green. Phalloidin, Blue, TOPRO (bar represent 50 μm). (G) Quantification of the length of the cellular processes (Values represent the standard deviation. ANOVA test, factor followed by Dunnet’s test. *p < 0.05, **p < 0.0013).
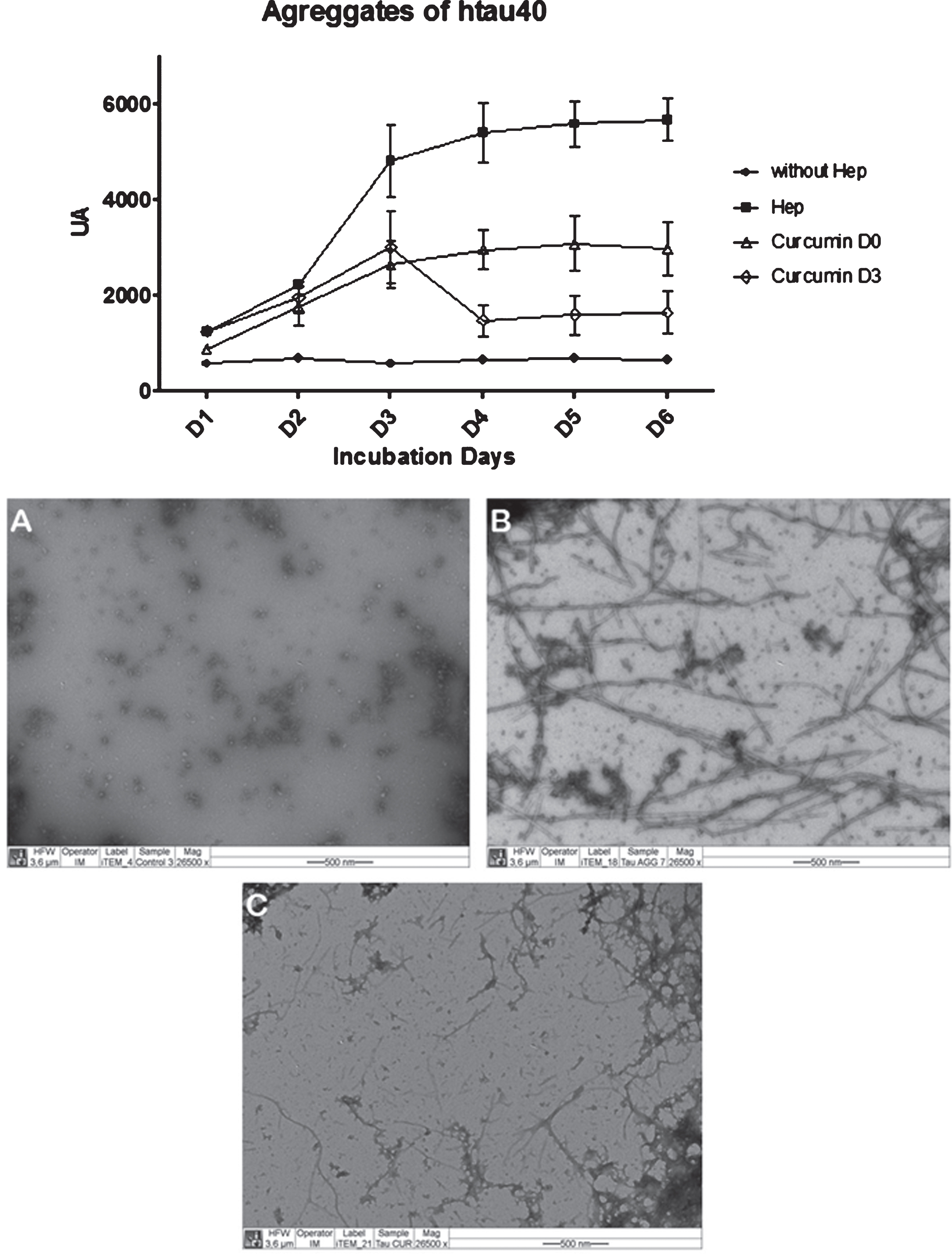
Effect of curcumin on aggregated tau protein. Aggregation kinetics were performed at 50 μM protein and 12.5 μM heparin as inducer in PBS buffer pH 7.4 at 37°C at the times indicated. Fluorescence (AU, arbitrary units) with Thioflavin S by exciting at 440 nm and emitting at 508 nm (n = 2) was quantified. After measuring on day 3, curcumin was added to a final concentration of 10 μM. Down: Images by transmission electron microscopy 50 μM protein tau incubated for 8 days, according aggregation protocol with heparin: tau incubated in the absence of heparin (A), tau incubated with heparin 12.5 μM (B), tau incubated with heparin 12.5 μM and 10 μM curcumin (C).
After exposing the cultures to the same stimulus of oxidative stress (Fe-NTA and H2O2) in the presence of curcumin (at concentrations 5 to 15 μM), they were closest to the control situation (Fig. 2E, 2F and 3E, 3F), reversing the appearance of the harmful effect of both agents that induce oxidative stress, and in both cases, the presence of curcumin has significant crop to quantify the length of cellular process (Fig. 2G and 3G) differences.
Effect of curcumin on the pathological aggregation of tau protein
To evaluate the effect of curcumin on the pathology aggregation of tau protein, an in-vitro assay protein aggregation was performed. A recombinant protein, htau40, corresponding to the complete fragment of the human protein expressed in E. coli and subsequently purified, resulting in the protein in its monomeric state, was used. This protein was added using heparin as aggregation inducing agent in a 1:4 in terms of the molar concentration of free monomeric protein. The formation of aggregates of htau40 was followed by thioflavin-S (Fig. 4A). Measuring Thioflavin-S was performed daily, and testing using htau40 were: (1) without heparin; (2) heparin; (3) heparin and curcumin from the starting day of the assay; (4) incorporating heparin curcumin from the day 3 assay. The fluorescence emitted by exciting the thioflavine-S is directly proportional to the formation of protein aggregates (Fig. 4).
As the results of this aggregation assay, a continuous decrease was observed in the formation of pathologically aggregated tau protein from the time when curcumin was added at time zero (T0) of the assay. A decrease in fluorescence was recorded presenting the lowest fluorescence as recorded by the control aggregation situation (htau40 and only heparin).
In the case of incorporating curcumin in an advanced state of aggregation (day 3, T3), we observed a significant decrease in fluorescence, evidencing the effects of curcumin on the unbundling of the pathological polymers already formed (Fig. 4A). In parallel, after completion of the test, the formation of pathological tau aggregates by transmission electron microscopy was corroborated (Fig. 4 B-D). It was important to evaluate comparatively, the effects of curcumin on cell viability of another type of non-neuronal cells. This observation evidenced the formation of aggregates in the situation of htau40 plus heparin and disassembly of these aggregates after adding curcumin at T3.
DISCUSSION
It has already been shown that the progress of AD associated with neuronal death processes are preceded by pathological tau aggregation. Therefore, is of great interest, from the therapeutic point of view, to search for compounds capable of interfering in abnormal tau aggregation, as well as compounds that have a neuroprotective capacity, in order to ameliorate the degree of injury and prevent continuous cell damage.
Curcumin is a natural component of the rhizome of turmeric (Curcuma longa) that has been used for years in the medicine of Eastern countries. Some investigations using this compound demonstrate their neuroprotective [20, 21] and anti-inflammatory activities [5, 22]. Also, antioxidant activity [23] has been evidenced. This has been of relevance in the development of potential treatments using curcumin or derivatives for neurodegenerative diseases. On the basis of this background, we evaluated the effect of curcumin on neuron-like cell lines (N2a), under conditions of oxidative stress (using 2 different agents that cause oxidative stress injury type) and also tested the ability to interfere in the formation of pathological tau aggregates, a pathognomonic feature of AD. In this context, we first evaluated cell viability of these cells treated with curcumin. Given the results obtained in the measurement with N2a cells (Fig. 1B), it was necessary to evaluate the effect of this compound in other cell type such as HEK293 line derived from kidneys, thus confirming the safety of curcumin in these type of cells and giving us further insights for future potential applications of this natural agent (Fig. 1C).
On this basis, the neuroprotective effects of curcumin were observed in both Fig. 2 (plus Supplementary Figure 1) and Fig. 3. This is evidenced by the morphological characteristics of the compound-treated cells and the response to harmful agents. Cells that have been treated with the noxious agent simultaneously with curcumin, exhibited a morphology similar to the control situation without treatment. This would realize that in the presence of curcumin could have an attenuation of oxidative response allowing normal operation and maintenance of cells exposed to injury. Investigations performed by the group of Tiwari obtained similar results; they concluded that in the presence of curcumin, there would be a decrease in the levels of reactive oxygen species in rats exposed to ethanol [23].
The most novel finding presented in this paper is the effect of curcumin on the pathological aggregation of tau [8, 24]. To date, the pharmaceutical industry has invested substantial amounts of money in inadequate targets for the treatment of AD. On this basis, the search for new therapeutic targets is of critical relevance. The results presented in Fig. 4 can demonstrate the ability of curcumin disaggregating preformed filaments in vitro, since a significant decrease in fluorescence once was detected when incorporated curcumin into the tau aggregated.
However, it appears that there is a slight increase in fluorescence which could be associated with the formation of new protein aggregates. This same phenomenon is observed when incorporated curcumin from time zero aggregation; there is an increase in the curve, not comparable to the control aggregation situation, but that could match the previous case suggesting that curcumin may have a time and a limited capacity for action. Data appears to suggest that curcumin could interact with a site on tau involved in its self-aggregation, thus preventing the pathological tau assembly, an interaction that may depend on the folding conditions of tau structure.
These latest findings are interesting particularly if we contextualize in the framework of the neuroimmunomodulation theory postulated by our group [8 , 24–26]. It has already been shown that pathological tau aggregates that are released into the extracellular medium are capable of inducing microglial activation, generating a cycle of neuronal damage [13]. Based on these observations, curcumin appear to play an important role in interrupting the neurodegenerative cycle and prevents tau self-assembly or just disassemble the tau pathological aggregates already formed. In addition, curcumin acts by preventing microglial activation, and as a natural anti-inflammatory compound.
Conclusion
AD is an increasing puzzle to medicine and society worldwide and that is why we must strengthen the scientific initiatives that aim to find effective compounds for its treatment. Curcumin is a naturally occurring compound with neuroprotective properties based on an increasing amount of scientific evidence. Our results demonstrate the neuroprotective effect, also showing action in disaggregating pathological tau protein filaments. Finally, our findings with curcumin are of great value for AD research since it opens new paths for research and development of new therapies for the disease based on specific cell targets.