
Abstract
Background:
Oxidative stress has been implicated in Alzheimer’s disease (AD) as a common pathway underlying neuronal damage causing huge impacts on cognitive functions in the AD process.
Objective:
Reduction and remodeling of γ-aminobutyric acid (GABA) signaling in AD may promote neuronal survival by regulating PI3K/Akt axis. Moreover, its activation exerts beneficial effects on AD by alleviating the neuronal oxidative stress injury. Considering these facts, we hypothesized the GABAB receptor as a novel therapeutic target for AD.
Methods:
To evaluate this hypothesis, a rat AD model was established by intraperitoneal injection of the GABAB receptor agonist (baclofen), PI3K/Akt signaling pathway agonist (740 Y-P), and antagonist (LY294002), respectively. The effects of GABAB activation on spatial memory and learning ability in the AD rats were measured by Morris water maze. Whereas the effects of GABAB and PI3K/Akt signaling pathway on apoptosis and oxidative stress injury were determined in vivo and in vitro using primary neuronal cultures.
Results:
We found that GABAB receptor activation restored spatial memory and learning ability of AD rats and suppressed the neuronal apoptosis and hippocampal atrophy by activating the PI3K/Akt signaling pathway. Additionally, GABAB receptor activation reduced the oxidative stress injury by lowering the MDA levels and increased the SOD, GSH-Px, and CAT levels via activation of the PI3K/Akt signaling pathway.
Conclusion:
Taken together, our results suggest that GABAB receptor activation repressed the oxidative stress injury implicated in neurons in AD rats via PI3K/Akt signaling pathway activation which may suggest a potential new therapeutic target for AD.
INTRODUCTION
Alzheimer’s disease (AD) is a progressive neurodegenerative condition influencing behavior, cognition, mood, and memory [1]. The main histological features of AD are extracellular accumulations of amyloid-β(Aβ) plaques along with axonal and dendritic atrophy and neuronal death leading to a progressive failure of cognition [2]. Other cellular features of AD include neurotrophic and neurogenesis failure, oxidative stress, heavy metal toxicity, and RNA modification [3]. Despite decades of intensive investigation into its clinical pathophysiology and basic biology [4], AD remained the major cause of age-related dementia, while the diagnosis of AD is an ominous prospect, given the lack of effective prevention and therapeutic strategies. Oxidative stress regulatory networks have been reported to influence the neuronal death in AD [5]. Oxidative stress injury is regarded as the final common pathway to neuronal damage from cerebrovascular accidents, contributing to mitochondrial damage in the aftermath of chronic hypoperfusion [6]. Besides, oxidative stress in AD exacerbates the neuronal stress arising from toxic protein aggregates [7]. Moreover, the participation of oxidative stress mechanisms in the pathology of AD has also been reported earlier [8]. Hence, the aforementioned findings speculate that the alleviation of oxidative stress injury presents a potential neuroprotective target in AD.
γ-aminobutyric acid (GABA) is a well-known amino acid neurotransmitter characterized by its inhibitory effects in the central nervous system. The three major types of GABA receptors are designated as GABAA, GABAB, and GABAC [9]. The metabotropic GABAB receptors, i.e., G-protein-coupled receptors, are thought to be involved in the etiologies of various neuropsychiatric disorders, presenting a potential novel target for therapeutic medications [10]. Furthermore, GABAB receptor expression correlates with working memory performance in aged rats [11]. A recent study has shown that activation of GABAB receptors in neutrophils induced PI3K-dependent neutrophil chemotaxis and microtubular rearrangement [12]. Furthermore, it has been exhibited that the PI3K/Akt signaling pathway affects neuronal survival due to its significant effects on cell growth and apoptosis under various conditions [13]. For instance, deficient insulin-PI3K-Akt signaling may trigger the AD neurodegeneration by decreasing O-GlcNAcylation thereby enhancing the aberrant neurodegeneration and tau hyperphosphorylation [14]. Considering these facts, in the present study, we hypothesized that the GABAB receptor activation could alleviate the oxidative stress injury of neuronal cells by activating the PI3K/Akt signaling pathway, thus, influencing the development of AD-like pathology.
MATERIALS AND METHODS
Ethical statement
This study was performed in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Institutional Animal Care and Use Committee of Linyi People’s Hospital.
AD modeling
Sterilized saline was used to prepare 1 nmol/μL Aβ25–35 solution (Sigma-Aldrich, MO, USA) which was stored in a 37°C incubator for 7 days. After incubation, Aβ25–35 solution was condensed and further stored in a 4°C refrigerator for future use. Sprague-Dawley (SD) rats were adaptively fed for one week and then modeled (fasting 12 h before operation). Rats were anesthetized with sodium pentobarbital (30 mg/kg body weight) by intraperitoneal anesthesia. After anesthesia, the hair on the top of the head was cut off and the rats were fixed. The hippocampal region of bilateral ventricles was injected with 10μg condensed Aβ25–35 (2 g/L) using a microinjector at one time. The needle was left for 10 min and then withdrawn slowly. Rats in the control group were injected with the same amount of normal saline.
Experimental animals
Seventy healthy male adult SD rats [specific pathogen-free (SPF); age, 2– 3 months, weighed 32.10±24.70 g; provided by the Experimental Animal Center of Sichuan Academy of Traditional Chinese Medicine (Sichuan, China)] were housed with free access to water and food in a 12/12 h day/night cycle at 25±2°C. After seven days of acclimatization, a total of 60 rats were randomly grouped with 10 rats each group. AD modeled rats were intraperitoneally injected with GABAB receptor agonist baclofen (2.0 mg/kg) [15], PI3K/Akt signaling pathway agonist 740-Y-P (10 mg/kg) [16], or PI3K/Akt signaling pathway inhibitor LY294002 (20 mg/kg) [15], or both baclofen (2.0 mg/kg) +LY294002 (20 mg/kg). Rats were treated differently for consecutive 6 weeks. The experimental protocol for treatments and behavioral tests can be seen in Fig. 1.
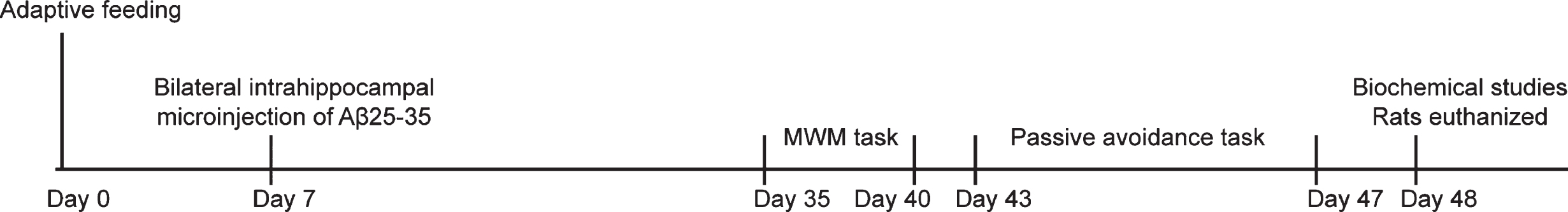
Experimental protocol for treatments and behavioral tests. AD was induced by injecting 10μg of condensed Aβ25–35 (2 g/L) into the dorsal CA1 region of the bilateral hippocampus in rats. Rats were injected intraperitoneally with balofen (2.0 mg/kg), 740 Y-P (10 mg/kg), and LY294002 (20 mg/kg), alone or in combination for 6 weeks.
Morris water maze test
Systems of rat behavior test, automatic image monitoring, and data processing were included in the Morris water maze test. Rats were studied in a pool with a diameter of 120 cm and a height of 60 cm, under the water temperature of 25±2°C. The diameter of the platform was 12 cm. The water tank was divided into 4 quadrants, and the escape platform was set in the center of one of the quadrants [17]. Rats underwent hidden platform trials for 5 days (4 trials per day; 60 s each time). Rats were then sent into the pool from 4 designated points. With one of the quadrants randomly selected and set as the initial position, rats were put into the water tank and the escape latency referred to rat finding the escape platform within 1 min. In each trial, if rats were able to find the platform within 1 min, the time rats spent was recorded and rats were allowed to stay in the platform for 1 min. If rats were unable to find the platform within 1 min, they were guided to the platform by the experimenter’s hand and stayed for 1 min. After 6 days, the platforms were removed the next day after the water tank navigation task was finished [18]. The times the rats spent in searching the pool quadrant within 1 min were recorded.
Single trial passive avoidance test
Rats were placed in an apparatus (50×16×40 cm) that was divided into illuminated and dark compartments by a sliding guillotine door. Rats were allowed to be habituated for 15 min in the first and second days. On the third day, all rats were placed in the illuminated compartment. After 3 min, the guillotine door was opened and latency to enter into dark section was measured from the time which door was lifted as an initial latency (IL). Next the guillotine door was closed and electric footstock (32 V, 6 s duration) was delivered. The rats with IL greater than 60 s were excluded. In retention trial (24 h after training), each rat was placed in the lighted chamber and after 3 min, the light was turned on and the latency to enter into dark section (step through latency, STL) was recorded. The cut-off for STL was 600 s.
Hematoxylin-eosin (HE) staining
Brain samples were removed from three rats in each group after anesthesia and dehydrated by gradient alcohol, cleared with xylene, and paraffin-embedded. The paraffin blocks were sliced into sections of 5μm thickness, mounted on glass slides, and dried overnight. Sections were from approximately the same stereological coordinates. After dewaxing, the sections were dyed with 0.5% hematoxylin, rinsed with clean water, differentiated with 75% hydrochloric acid in alcohol, and then treated with ammonia water to return the blue color. Subsequently, the sections were stained with eosin, dehydrated with gradient alcohol, cleared with xylene, and sealed with neutral gum in a fume cupboard overnight. Finally, histopathological changes were observed under an optical microscope (DMM-300D, Shanghai Caikon Optical Instrument Co., Ltd., Shanghai, China).
Dichloro-dihydro-fluorescein diacetate (DCFH-DA) for fluorescent quantitation
Whole hippocampus samples were mechanically homogenized using a glass-Teflon apparatus in ten volumes of PBS. After centrifugation, the supernatant was collected and incubated with DCFH-DA fluorescent probe solution at a final concentration of 10μmol/L at 37°C in 5% CO2 for 20 min. Afterward, the fluorescence intensity of DCFH was measured by a fluorescence spectrophotometer with excitation wavelength set at 488 nm and an emission wavelength of 525 nm. The mean fluorescence intensity of DCFH was used to calculate the reactive oxygen species (ROS) concentration.
Enzyme-linked immunosorbent assay (ELISA)
Three rats were randomly selected from each group and the hippocampal tissues were mechanically homogenized with minimum essential medium (MEM) to obtain 10% tissue homogenate. The bicinchoninic acid (BCA) kit (T9300A, Takara Bio Inc., Otsu, Shiga, Japan) was used to quantify the protein content of uncentrifuged homogenates. The activity of superoxide dismutase (SOD) was determined by the xanthine oxidase method and expressed as U mgprot–1. The glutathione peroxidase (GSH-Px), catalase (CAT), and iNOS activities were determined according to the instructions of the ELISA kits and expressed by U·mgprot–1. The malondialdehyde (MDA) concentration was determined by the thiobarbituric acid method and expressed by nmol·mgprot–1. The ELISA kits contained MDA kit (CSB-E08559m, Cusabio Biotech, Wuhan, China), SOD activity detection kit (BC1095, Solarbio Science & Technology, Beijing, China), CAT kit (BC0200; Solarbio Science & Technology), GSH-Px assay kit (Mito Sciences, Eugene, OR, USA), and iNOS kit (SEKH-0501; Solarbio Science & Technology). Specifically, the standard well was added with 50μL standards with different concentrations while the sample wells were added with 10μL samples homogenized with MEM and 40μL sample diluent. In addition to the blank well without treatment, standard and sample wells were added with 100μL horseradish peroxidase (HRP) labeled detection antibody followed by the incubation at 37°C for 60 min with the reaction wells sealed. Each well was then washed with detergent repeatedly 5 times. Subsequently, each well was added with 50μL of substrate A or B and incubated for 15 min in the dark at 37°C. After each well was added with 50μL terminating solution, the optical density (OD) value of each well was measured at 450 nm wavelength within 15 min. Finally, the OD value of each well was evaluated by a fluorescence spectrophotometer (NanoDrop3300, Thermo Fisher Scientific, Waltham, MA, USA) at 450 nm wavelength. The activity of SOD, GSH-Px, CAT, and iNOS, as well as MDA contents, were calculated respectively.
Detection of nitric oxide (NO) and glutathione (GSH) contents
Proteins from tissue and cell samples were quantified by BCA kit (T9300A, TaKaRA, Tokyo, Japan).The contents of NO and GSH were measured following the instructions of NO kit (BC1470, Solarbio Science & Technology) and GSH content detection kit (BC1175, Solarbio Science & Technology).
Bioinformatics analysis
GABAB receptor was identified as an important gene in AD by existing literature. The online analysis tool MEM (https://biit.cs.ut.ee/mem/index.cgi) was used to predict the co-expression relationship among the top 5,000 significant genes which were obtained by removing duplicates. KEGG enrichment analysis was performed by KOBAS (http://kobas.cbi.pku.edu.cn/kobas3) and the downstream pathways were identified by combining with existing literature.
Isolation and culture of hippocampal neuronal cells
Hippocampus was isolated bluntly accompanied by vessels and meninges removed. Hippocampus was sectioned into pieces with a diameter of 0.4 mm. Sections were then reacted with 0.25% trypsin and 0.04 DNA enzyme for 12 min and the reaction was terminated by adding horse serum. Cells were dispersed by pipette trituration for 10 times. The cell suspension was cultured in a MEM containing 10% FBS, 5% horse serum, 25 mmol/L KCI, 10 mmol/L HEPES, 105 U/L penicillin and 0.1 g/L streptomycin. Following the filtration by a nylon mesh filter with a diameter of 75μm, samples were cultured in a 35 mm dish coated with Poly-L-lysine hydrobromide slides and incubated in 5% CO2 and 95% O2 at 37°C with a density of 0.6×109 L–1. At day 3, 5μmol/L cytarabine was supplemented to the culture and the solution was replaced after every 24 h. At day 7 of post culture, samples were subjected to neuron-specific enolase immunocytochemistry staining via the SP method to identify hippocampal neuronal cells. Cultured cells were treated with Aβ (with the final concentration of 25μmol/L), baclofen (with the final concentration of 25μmol/L), 740 Y-P (with the final concentration of 20μmol/L),or LY294002 (with the final concentration of 10μmol/L), alone or in combination. Each treatment lasted for 24 h. Untreated cells were taken as control.
Flow cytometry
The successfully identified hippocampal neurons were cultured for 7 days followed by the grouping. Then, the grouped neuronal cells were cultured for 48 h. Cells were fixed overnight in precooled 70% ethanol at 4°C, and then incubated with 10μL of RNase enzyme for 5 min at 37°C. Cells were then stained with 1% propidium iodide (PI) (40710ES03, Shanghai Qian Chen bio Science & Technologies Co., Ltd., Shanghai, China) in the dark for 30 min. Afterward, the cells were analyzed with a flow cytometer (FACSCalibur, BD, FL, NJ, USA) at the excitation wavelength of 488 nm for cell apoptosis detection.
Western blot analysis
Total proteins were extracted from the hippocampal tissues. Protein concentration was measured according to the instructions of the BCA protein quantitative kit (23225, Pierce, Rockford, IL, USA). The protein was separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (P1200, Beijing Solarbio Life Sciences Co., Ltd., Beijing, China) and transferred onto polyvinylidene fluoride (PVDF) membranes (HVLP04700, Millipore, Bedford, MA, USA). Then the membranes were washed twice with Tris-buffered saline with Tween (TBST), blocked with 5% skimmed milk at room temperature for 2 h, and washed again with TBST three times. Subsequently, the membrane was incubated with the primary rabbit anti-mouse antibodies against Bcl2 Associated X (Bax), (1:1000, ab32503), B-cell lymphoma-2 (Bcl-2) (1:1000, ab59348), PI3K (1:1000, ab191606), phosphorylated PI3K (p-PI3K, 1:500, ab182651), Akt (1:500, ab8805), phosphorylated Akt (p-Akt, 1:500, ab38449), caspase-3 (1:500, ab13847), Cleaved-caspase 3 (1:1000, ab2302) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 1:2000, ab9485) at 4°C overnight. All antibodies were procured from Abcam Inc. (Cambridge, UK). After that, the membranes were incubated with HRP-labeled goat anti-rabbit immunoglobulin G (IgG) secondary antibody (1:2000, sc-2004, Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA) at room temperature for 2 h. Following after the membranes were washed three times with TBST (10 min per wash) and developed with the addition of DAB solution. Gel imaging instrument (GelDocXR, Bio-Rad, Hercules, CA, USA) was used to make photographs of the membranes. Analyzed using Quantity One v4.6.2 software, the relative protein level was expressed as the ratio of the gray value of the target protein band to that of the GAPDH internal reference band.
Statistical analysis
SPSS 21.0 statistical software (IBM SPSS Statistics, Chicago, IL, USA) was used for data analysis. The measurement data were expressed as mean±standard deviation. Data between the two groups were analyzed using unpaired t-test. The comparisons among multiple groups were analyzed by one-way analysis of variance (ANOVA) with Tukey’s post hoc test. The comparisons at different time points were analyzed using repeated-measures ANOVA with Bonferroni’s post hoc test. A value of p < 0.05 demonstrated statistical significance.
RESULTS
Memory impairment and oxidative stress changes were observed in AD rat models
To verify if AD rat models were established successfully, the Morris water maze test was initially performed. It was found that compared with the normal group, the average escape latency of rats in the AD group was significantly increased (Fig. 2A) and the average number of times crossing the platform was significantly decreased (Fig. 2B). Passive avoidance test showed that the initial latency IL of rats in the AD group was not significantly different from that in the normal group, but the STL time of rats in the normal group decreased after electric footstock (Fig. 2C). HE staining was performed on the brain tissues of rats in the AD group and the normal group, and it was found that the neuronal cells in the normal group were arranged neatly and round with complete cellular structure, accompanied with clear cell membrane and nucleus. There was no obvious swelling and necrosis. In contrast, the neuronal cells in the AD group were extremely disordered, irregular in size and shape and the number of neuronal cells was significantly reduced with blurred cellular structure, indicating that the AD rat models were established successfully (Fig. 2D).
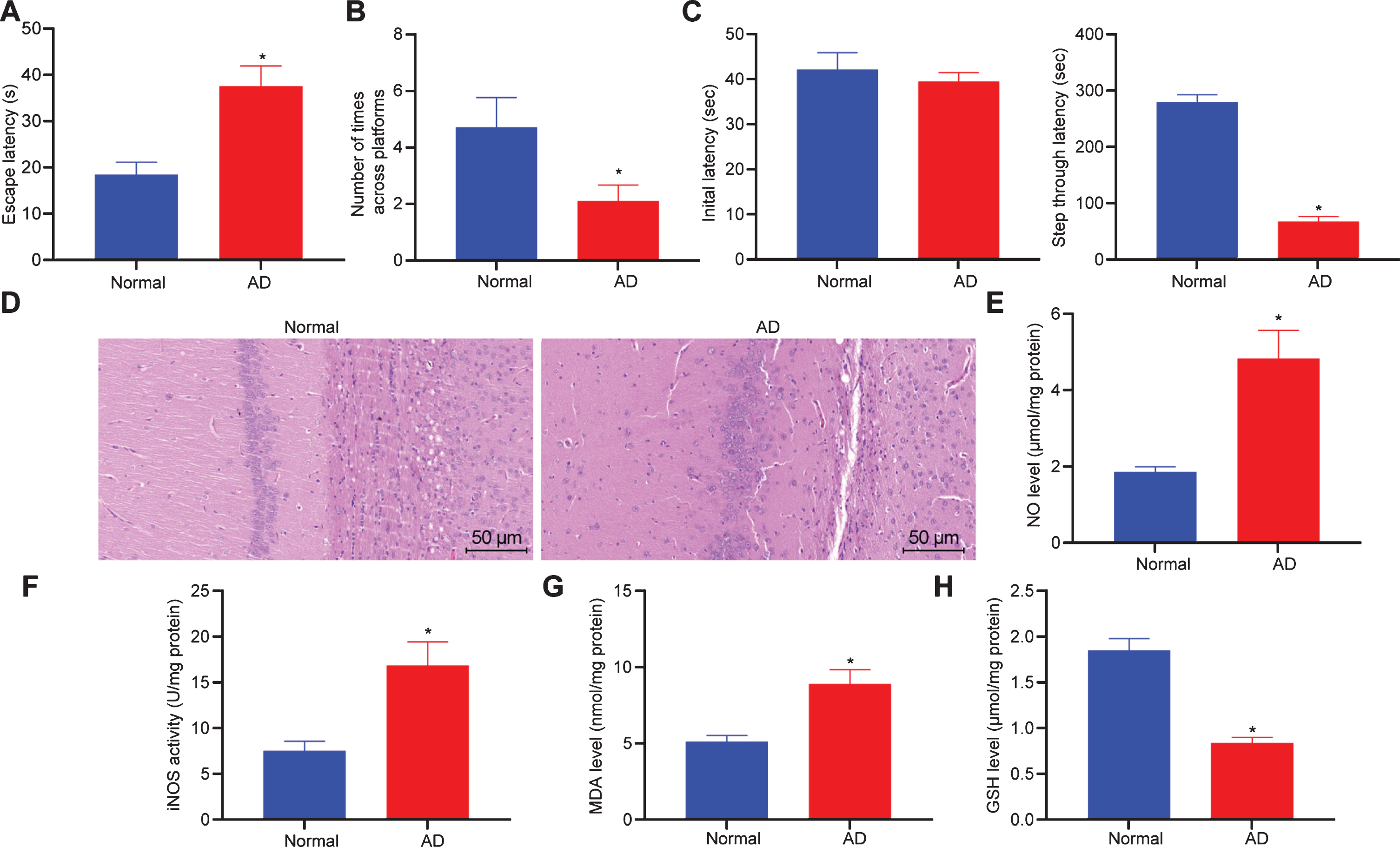
AD modeling induces memorial impairment and oxidative stress change. A, B) Escape latency (A) and times of crossing platforms (B) between the normal and AD rats by Morris water maze test. C) Comparison of IL and STL of the normal and AD rats (n = 10) by passive avoidance test. D) Pathological changes of brain tissues of rats determined by HE staining (×200). E– H, Content of NO (E), activity of iNOS (F), contents of MDA (G), and GSH (H) in hippocampal tissues of rats detected by ELISA. Measurement data are expressed as mean±standard deviation. Data comparisons between the two groups are analyzed by unpaired t-test. *p < 0.05, *p < 0.01, compared with normal rats. Cellular experiments were repeated 3 times.
Oxidative damage is one of the main causes of neuronal degeneration and loss of function in patients with AD, which can interact with Aβ to activate corresponding signaling pathways, thereby promoting the occurrence and progression of AD. The oxidative stress status in the hippocampus of the established AD rat model was detected (Fig. 2E– H). Compared with the normal group, the levels of NO, MDA, and iNOS activity in the AD group were elevated, while the levels of GSH decreased, indicating that the oxidative stress status was changed in the brain tissues of AD rats.
GABAB receptor-mediated activation of PI3K/Akt signaling pathway suppresses cell apoptosis in AD
It is reported that GABA participates in the memory process, and activation or inhibition of GABA receptors can improve cognitive ability and inhibit acute Aβ-induced memory impairment in rats [19].The top 5,000 co-expression relationships of GABAB receptors (NCBI included: GABBR1) were predicted by MEM analysis. After removing the duplicated genes, 3,370 co-expressed genes were obtained, of which the least significant was p = 1.06E– 05, that is, all genes were significantly co-expressed with GABAB receptors. By KEGG enrichment analysis of these 3,370 genes with the online analysis tool KOBAS, we found that these genes were mainly related to metabolic, viral infection, and cancer-related pathways (Fig. 3A), of which 50 GABAB genes were significantly enriched in the PI3K/AKT signaling pathway. Meanwhile, it was found that GABAB receptor could activate PI3K/Akt signaling pathway in cerebellar neurons of rats [20] and activation of PI3K/Akt signaling pathway could inhibit apoptosis of hippocampal neurons [21]. However, whether GABAB receptor could regulate the activation of PI3K/Akt signaling pathway to affect memory impairment in AD rats induced by Aβ is largely unknown.

GABAB receptors activate the PI3K/Akt signaling pathway to suppress hippocampal cell apoptosis. A) KEGG enrichment of 3,370 co-expressed genes of GABAB receptor conducted by KOBAS. Y-axis represents enriched entries, X-axis represents the number of genes enriched in the entries. The color red shows greater significance |lgP|, and vice versa. Color blue shows smaller significance. B) Protein bands of the PI3K/Akt signaling pathway related proteins and apoptosis-related proteins in rat models determined by western blot analysis. C) Quantitative analysis of the PI3K/Akt signaling pathway related proteins in rat models. D) Quantitative analysis of the cell apoptosis-related proteins in rat models. *p < 0.05 compared with normal rats. *, #p < 0.05 compared with AD rats. # , &p < 0.05 compared with AD rats treated with baclofen (n = 3). Measurement data are expressed as mean±standard deviation. Data comparisons among multiple groups are analyzed by one-way ANOVA with Tukey’s post hoc tested.
Moreover, the successfully modeled rats were treated with baclofen (the AD + baclofen group), 740Y-P (PI3K/Akt signaling pathway agonist) (the AD + 740-Y-P group), and LY294002 (the AD + LY294002 group), respectively, with 10 rats in each group. The hippocampal extracts from rats were differently treated for detection of the extent of PI3k and Akt phosphorylation by western blot analysis (Fig. 3B, C). The extent of PI3k and Akt phosphorylation was declined in rats of the AD + baclofen group, the AD + 740Y-P group, the AD + LY294002 group, and the AD + baclofen + LY294002 group
Furthermore, we analyzed the expression of Bax, Bcl-2, cleaved caspase 3, and Caspase-3 in the hippocampal extracts in each group using western blot analysis (Fig. 3B, D). It was revealed that the expression of Bcl-2 and Caspase-3 increased in the AD + baclofen group and the AD + 740Y-P group along with the decreased expression of Bax and cleaved caspase-3 expression; however, in the AD + LY294002 group, Bcl-2 and Caspase-3 expression was remarkably reduced while the Bax and cleaved caspase-3 expression increased. The rats in the AD + baclofen + LY294002 group exhibited downregulated expression of Bcl-2 and Caspase-3 whereas the upregulated expression of Bax and cleaved caspase-3 was observed compared with the rats in the AD + baclofen group. Collectively, these results suggested that the GABAB receptor could activate the PI3K/Akt signaling pathway to suppress apoptosis of hippocampal cells
GABAB receptor activation improves the spatial memory and learning abilities of AD rats by activation of PI3K/Akt signaling pathway
With an attempt to illustrate how the activated GABAB receptor and the PI3K/Akt signaling pathway affect spatial memory and learning abilities of AD rats, the Morris water maze experiment was performed to evaluate the spatial memory and learning ability of rats. Our results showed that in contrast with the untreated AD rats, the escape latency was shortened, yet the number of times crossing platforms was increased in the AD + baclofen group and the AD + 740Y-P group; however, an opposite trend in results was observed in the AD + LY294002 group (all p < 0.05). Relative to the AD + baclofen group, the AD + baclofen + LY294002 group exhibited prolonged escape latency and decreased the number of times crossing the platforms (all p < 0.05) (Fig. 4A, B).
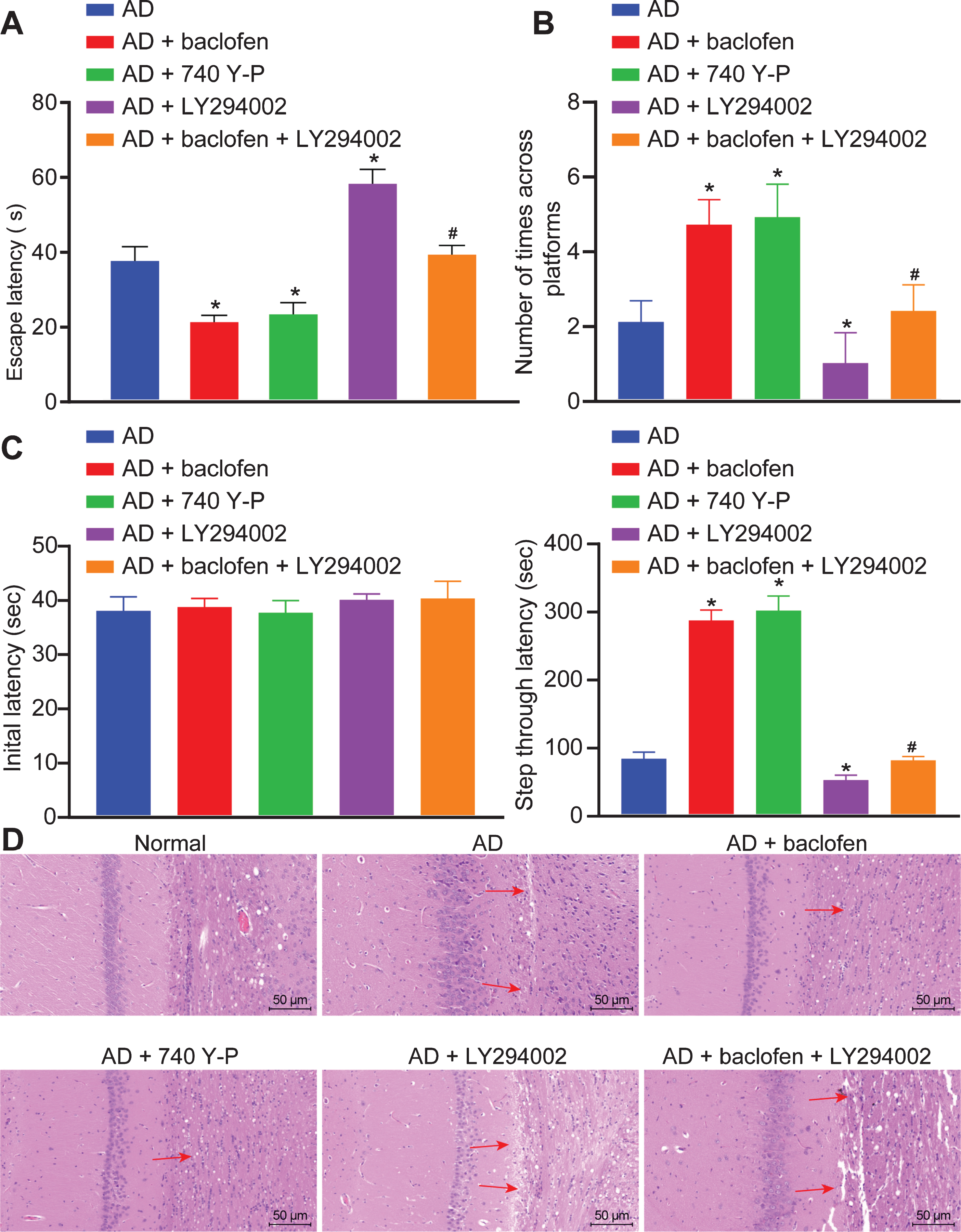
GABAB receptors increase the number of times crossing platforms and reduces escape latency in AD model rats by activation of PI3K/Akt signaling pathway. A) Comparisons of escape latency of rats in each group. B) Comparison of the number of times crossing platforms of rats in each group. C) Comparison of IL and STL of the normal and AD rats (n = 10) by passive avoidance test. D) Pathological changes of brain tissues of rats determined by HE staining (×200). *p < 0.05 versus the AD rats (n = 10);#p < 0.05 versus the AD rats treated with baclofen (n = 10). The results are measurement data and expressed as mean±standard deviation. Data comparisons among multiple groups are analyzed by one-way ANOVA with Tukey’s post hoc tested.
Through passive avoidance test, it was found that there was no significant difference in the IL of rats in each group (Fig. 4C). After training, compared with the AD group, STL was significantly increased in the AD + baclofen group and the AD + 740Y-P group, while inhibition of PI3K/Akt signaling pathway by LY294002 could reduce STL. STL was also decreased when rats treated with GABAB receptor activator baclofen alone, or combined with LY294002.
HE staining was used to detect the brain tissue of rats in each group, and the study found that (Fig. 4D), compared with the AD group, the hippocampal neurons were uniformly colored and arranged tightly in the AD + baclofen group and the AD + 740 Y-P group. While in the AD + LY294002 group, the number of layers and number of neurons in the hippocampal tissue were reduced with larger intercellular space and disordered arrangement. Moreover, some of the cells were shrunk with pyknotic nucleus, and rupture was observed with the whole cell shown in deeply stained red. When compared with the AD + baclofen group, the AD + baclofen + LY294002 group had a decrease in the number of neuronal cell layers with a larger intercellular space and disordered arrangement, showing aggravated brain tissue damage. In conclusion, the GABAB receptor could restore the spatial memory and learning abilities of AD rats by activation of the PI3K/Akt signaling pathway.
Activation of GABAB receptor and PI3K/Akt signaling pathway decrease ROS levels in AD models
To further evaluate the effects of GABAB receptor and PI3K/Akt signaling pathway on ROS levels, level of ROS was measured for evaluating the oxidative stress injury of hippocampal tissues in each group. Our results clarified that the degree of ROS levels was higher in all Aβ-treated hippocampal tissues than that in untreated hippocampal tissues (all p < 0.05). In contrast to the Aβ-treated hippocampal tissues, the degree of ROS levels was decreased in the Aβ-treated hippocampal tissues with baclofen or 740 Y-P treatment, however, it potentiated after LY294002 treatment (all p < 0.05). Additionally, the degree of ROS levels was increased in the Aβ hippocampal tissues treated with baclofen and LY294002 relative to the Aβ hippocampal tissues treated with baclofen (all p < 0.05) (Fig. 5).
MDA, GSH-Px, CAT, and SOD are reported to be oxidative stress markers [16, 22] in hippocampal tissues, thus we quantified these oxidative stress markers in Table 1. ELISA results showed that the normal group exhibited a significantly lower MDA level while distinctly higher levels of SOD, GSH-Px, and CAT than the AD group, the AD + baclofen group, the AD + 740 Y-P group, the AD + LY294002 group, and the AD + baclofen + LY294002 group (p < 0.05). The MDA level was notably lower while levels of SOD, GSH-Px, and CAT were significantly higher in the AD + baclofen or AD + 740 Y-P group whereas in the AD + LY294002 group, the MDA level was remarkably elevated but the levels of SOD, GSH-Px, and CAT were reduced (all p < 0.05). In contrast with the AD + baclofen group, the MDA level was increased; however, the levels of SOD, GSH-Px, and CAT were reduced in the AD + baclofen + LY294002 group (all p < 0.05). These findings suggested that activation of GABAB receptor by baclofen decreases the ROS levels, while blockade of the AKT/PI3K signaling pathway can partially counteract the neuronal oxidative damage improved by GABAB receptor activation.
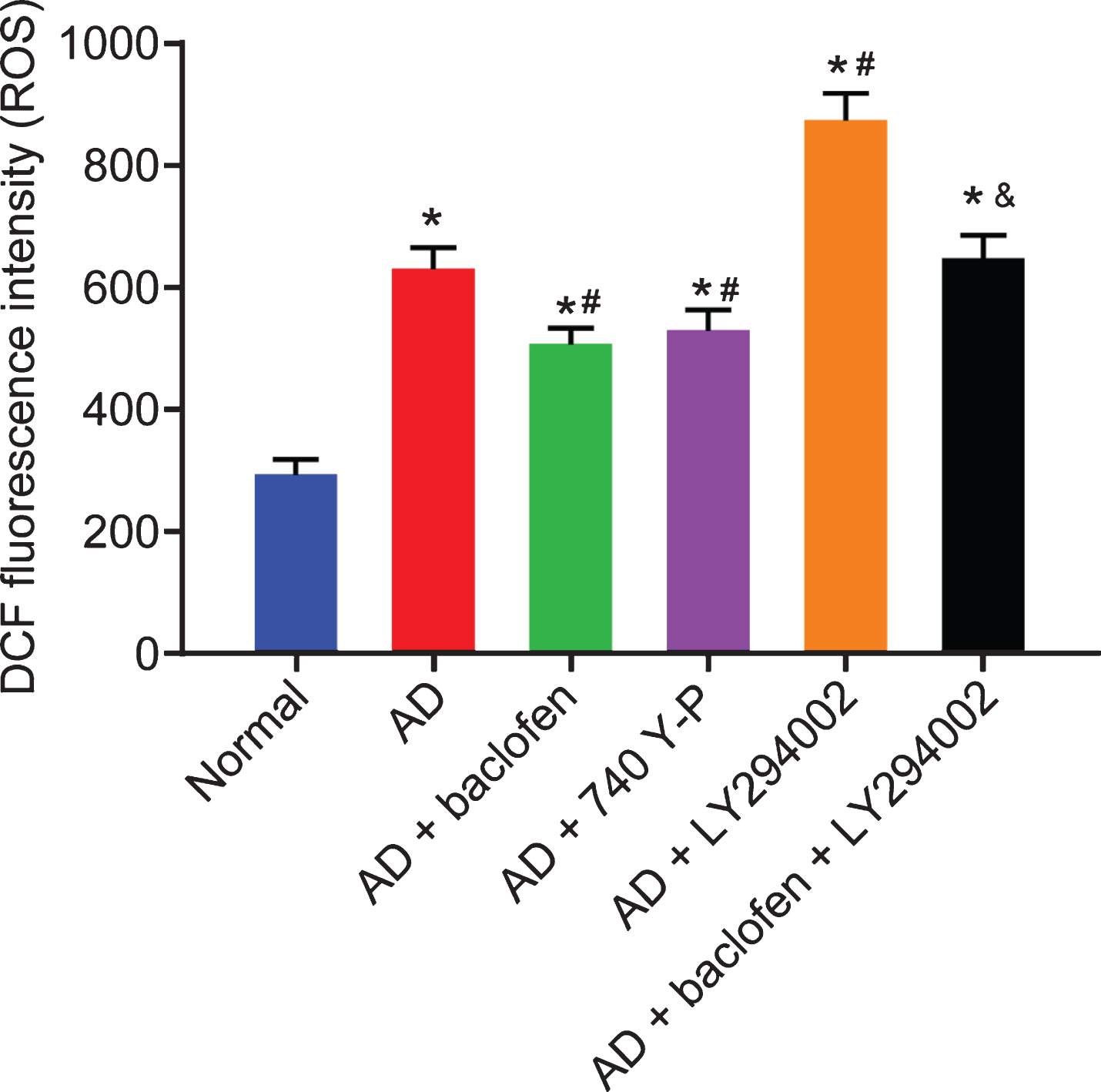
GABAB receptor activation decreases ROS levels by activating the PI3K/Akt signaling pathway. Quantification of ROS levels in neuronal hippocampal tissues of rats in each group by DCFH-DA.*p < 0.05 compared with normal rats. #p < 0.05 compared with AD rats. &p < 0.05 compared with AD rats treated with baclofen (n = 10). The results are measurement data and expressed as mean±standard deviation. Data comparisons among multiple groups are analyzed by one-way ANOVA with Tukey’s post hoc test.
Changes of GSH-Px, CAT, SOD activities and MDA content in hippocampal tissues of rats
*p < 0.05 versus the normal group; #p < 0.05 versus the AD group; &p < 0.05 versus the AD + baclofen group. The results are measurement data, and expressed as mean±standard deviation. Data comparisons among multiple groups are analyzed by one-way ANOVA with Tukey’s post hoc test, n = 3. ANOVA: analysis of variance; GSH-Px: phospholipid hydroperoxide glutathione peroxidase; CAT: catalase; SOD: superoxide dismutase; MDA: malondialdehyde; AD: Alzheimer’s disease; IGF-1: insulin-like growth factor gene.
Activation of both GABAB receptor and the PI3K/Akt signaling pathway reduces neuronal apoptosis and ameliorates oxidative damage
To evaluate whether activation of GABAB receptor reduces apoptosis and ameliorates oxidative stress injury in cells through PI3K/Akt signaling pathway, hippocampal neurons from SD rats were extracted and cultured for 7 days followed by treatment of different concentrations of baclofen and Aβ for 24 h. The activation degree of Akt was detected by western blot assay, and the results showed that (Fig. 6A), the extent of Akt phosphorylation increased as baclofen concentration enhanced and tended to be stable at the concentration of 20μmol/L, indicating that the activation of Akt mediated by GABAB receptor was dependent by baclofen in a certain concentration range. Therefore, the concentration of 10μmol/L was chosen for subsequent experiments.

Activation of GABAB receptor and PI3K/Akt signaling pathway attenuates apoptosis of hippocampal cells. A) Neuronal cells extracted from SD rats were cultured for 7 days followed by treatment with baclofen at different concentrations (0μmol/L, 1μmol/L, 5μmol/L, 10μmol/L, 20μmol/L, 30μmol/L, and 40μmol/L) with Aβ (25μmol/L) for 24 h. Phosphorylation of Akt was detected using western blot analysis. B) Apoptosis of hippocampal cells after different treatment determined by flow cytometry (n = 3). C) Results of western blot analysis (n = 3) on protein levels of neuronal cell apoptosis-related factors and PI3K/Akt signaling pathway-related proteins. D, Oxidative status of differently treated hippocampal cells. *p < 0.05 versus the control group. #p < 0.05 versus AD rats. &p < 0.05 versus AD rats treated with baclofen. The results are measurement data and expressed as mean±standard deviation. Data comparisons among multiple groups are analyzed by one-way ANOVA with Tukey’s post hoc test.
The apoptosis of cultured hippocampal neurons was detected by flow cytometry with the combination of baclofen, 740 Y-P, LY294002, and Aβ. The results showed that (Fig. 6B), compared with the control group, the number of apoptotic cells in other groups was significantly increased, compared with the Aβ group, the apoptotic rate in the Aβ+ baclofen group and the Aβ+ 740 Y-P group was reduced, while the apoptotic rate in the Aβ+ LY294002 group was increased in contrast to the Aβ+ LY294002 group. Compared with the baclofen group, the apoptotic rate was increased in the Aβ+ baclofen + LY294002 group. Western blot was conducted to detect the expression of apoptosis-related proteins Bax, Bcl-2, cleaved caspase 3 and Caspase-3 [16, 23]. The results showed that (Fig. 6C) compared with Aβ group, the levels of Bcl-2 and Caspase-3 increased significantly in the Aβ+ baclofen group and Aβ+ 740 Y-P group, while the levels of Bax and cleaved caspase 3 decreased. As for the Aβ+ LY294002 group, Bcl-2 and Caspase-3 levels decreased distinctly, accompanied with elevated levels of Bcl-2, Caspase-3. Being compared with the Aβ+ baclofen group, the expression of Bcl-2 and Caspase-3 reduced while Bax and cleaved caspase-3 increased in the Aβ+ baclofen + LY294002 group. To evaluate how activation of GABAB receptor and PI3K/Akt signaling pathway reduced the expression of PI3K/Akt signaling pathway-related proteins in cultured hippocampal neurons, AKT and PI3K were further detected by western blot. The results showed that (Fig. 6C) compared with Aβ group, the extent of AKT and PI3K phosphorylation was increased in Aβ+ baclofen group and the Aβ+ 740 Y-P group, while the extent of AKT and p-PI3K phosphorylation prominently decreased in Aβ+ LY294002 group. When compared with the Aβ+ baclofen group, the extent of AKT and PI3K phosphorylation was curtailed in the Aβ+ baclofen + LY294002 group. These results indicated that activation of GABAB receptor could enhance downstream PI3K/Akt signaling pathway expression in Aβ-induced cells, thereby inhibiting neuronal cell apoptosis.
The oxidative stress status in neuronal cells was detected by ELISA, and the results showed that (Fig. 6D) oxidative damage was observed in Aβ-treated neuronal cells. After treatment with GABAB activator baclofen or PI3K/Akt activator 740 Y-P, ROS, MDA, and NO levels were downregulated, together with upregulated SOD and GSH-Px activities. However, PI3K/Akt inhibitor LY294002 induced the opposite effects. The combined treatment of LY294002 with baclofen could hinder the ameliorating effects of baclofen activated-GABAB on oxidative damage in neuronal cells.
The above results further showed that activation of GABAB could enhance the extent of PI3K/Akt phosphorylation, reduce the apoptosis of neurons induced by Aβ, and ameliorate the oxidative damage in neuronal cells.
DISCUSSION
AD is a neurodegenerative disease clinically characterized by memory loss and a variety of other cognitive impairments [24]. Dysfunctions in the neurotransmitter and receptor systems of the amino acid neurotransmitter GABA might increase the fragility of hippocampal pyramidal neurons to other stressors in AD [25]. Therefore, we investigated the role of GABAB receptor in AD model rats, established by intracerebral infusion of Aβ. Collectively, our results indicated that GABAB receptor activation relieved the oxidative stress injury of neuronal cells in an AD rat model through the activation of the PI3K/Akt signaling pathway.
Notably, our study demonstrated that GABAB receptor activation promoted the phosphorylation of PI3K and Akt, reduced the ROS levels and oxidative stress markers, and inhibited neuronal apoptosis. Therefore, we speculated that GABAB receptor activation could alleviate the oxidative stress injury of neuronal cells by activating the PI3K/Akt signaling pathway. Consistently, treatment with the GABABR2 agonist has been reported to trigger the activation of Akt and modulates the migration of neutrophils in a PI3K-dependent mechanism [26]. Meanwhile, chemotaxis mediated by GABABRs is closely connected with PI3K-dependent Akt phosphorylation [12]. In this study, we reported that GABAB receptor activation increased the spatial memory and learning ability of AD rats by activating the PI3K/Akt signaling pathway. Intriguingly, GABAB receptors have attracted great attention in preclinical research as potential mediators of the etiology of major depressive disorder [27]. It has been reported that the insulin-dependent PI3K/Akt signaling pathway loses activity in AD [14]. Taken above, those references supported our statement that GABAB receptor was able to boost the activation of PI3K/Akt signaling to further alleviate oxidative stress injury in AD.
The altered levels of Bcl-2 family proteins, i.e., Bax and Bcl-2, previously have been detected in the vulnerable neuronal cells and could be speculated as major factors of neuron loss in AD [28]. Besides, the activation of caspase-3 is associated with apoptosis through nuclear lamin cleavage and nucleus breakdown [29]. Our present study also demonstrated that GABAB receptor activation remarkably inhibits the apoptosis of neuronal cells in AD via activation of PI3K/Akt signaling pathway, which was indicated by upregulated Bcl-2 levels and down regulated levels of Bax, Cleaved-caspase 3.
The existing literature shows that GABAB1 receptor exerts a neuroprotective effect in the ethanol-induced neuronal cell death model via mutual interactions with (antioxidant) anthocyanins and vitamin C [30]. Of note, a recent study has identified a potential role of GABAB receptor for neurodegenerative disorder therapy, given that their pharmacological activation of GABAB receptor retards apoptosis of neuronal cells by transactivation of IGF-1 receptor [31]. However, compared to the study of Tu et al., our investigation was mainly based on a rat AD model, which showed demonstrable behavioral deficits in a spatial learning paradigm. We further provided evidence for the importance of GABAB receptor-mediated PI3K/Akt activation in neuronal apoptosis and oxidative stress injury in vitro and in vivo. Consistently, it has been noted that the PI3K/Akt/Bad/BclXL-regulated mitochondrial apoptotic pathway inhibits the Aβ-protein-induced neurotoxicity of primary rat cortical neurons in AD [32]. Specifically, suppression of the PI3K/Akt signaling pathway has been attributed to the acceleration of cell apoptosis which further stimulates the activation of downstream pathways impairing mitochondrial integrity [33].
Notably, our study exhibited that the activation of the GABAB receptor reduced the ROS level via activation of the PI3K/Akt signaling pathway, indicating a significant reduction of MDA concentrations while the upregulated activity of SOD, GSH-Px, and CAT in brain neuronal cells, respectively. Particularly, MDA and SOD are well-known indicators of the oxidative stress response, thus alteration of their levels is closely associated with the phenomenon of transient allodynia [34]. A recent study has revealed that the concentration of GSH-Px is significantly reduced after oxidative stress injury induced by cerebral ischemia/reperfusion in rats [35], thus highlighting the relevance of GSH-Px and oxidative stress injury. Peculiarly, CAT functions as a critical antioxidant enzyme in the metabolic disposal of ROS, leading to the mediation of oxidative stress [36]. The fact that cellular injury due to oxidative stress is considered as a key factor in the occurrence and progression of most neurodegenerative diseases, including AD [37]. However, it has been revealed that GABA significantly inhibits the oxidative stress injury in gastric tissue through elevation of SOD and CAT levels and a decline of MDA level [38]. Interestingly, GABA receptors have also been reported to interact with various bioactive compounds presented in parboiled germinated brown rice to restrain CCl4-induced oxidative stress injury in the liver [39]. Consistent with our findings, it has been indicated that the activation of PI3K/Akt signaling pathway attenuates oxidative stress injury in a rat AD model [33].
Conclusion
In summary, GABAB receptor activation contributed to the inhibition of apoptosis and increased activation of the downstream PI3K/Akt signaling pathway in a rat model of AD. Additionally, baclofen induced GABAB receptor activation further reduced the ROS levels and restored the spatial memory and learning abilities of AD rats by activation of the PI3K/Akt signaling pathway. These results may provide a theoretical basis for further understanding of AD pathology.