
Abstract
Background:
Parkinson’s disease (PD) is the second most prevalent progressive neurodegenerative disease, second only to Alzheimer’s disease, with motor disorders and cognitive impairment. Bergenin (Berg), extracted from the herb of Saxifrage stolonifera Curt. (Hu-Er-Cao), has anti-tumor, anti-inflammation, anti-oxidative stress, and neuroprotective properties.
Objective:
In this study, we wanted to investigate the effects of Berg on PD and the underlying mechanisms.
Methods:
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) was used to introduce PD symptoms in mice. The expression levels of tyrosine hydroxylase, dopamine, and Iba-1 were examined. The levels of a series of inflammatory mediators were measured by qPCR. In addition, the PI3K/Akt signaling pathway was investigated to illustrate the underlying mechanism. In vitro, PC12 cells subjected to lipopolysaccharide (LPS) were treated with Berg.
Results:
We found that MPTP injection introduced motor deficits, apoptosis of neurons and inflammation, as well as inhibited the PI3K/Akt signaling pathway. However, Berg treatment suppressed the MPTP-induced alterations. In vitro, Berg attenuated the cytotoxic effects on PC12 cells induced by the culture supernatants derived from LPS-induced microglial cells.
Conclusion:
Berg attenuated the PD symptoms via activating the PI3K/Akt signaling pathway in vivo and in vitro.
Keywords
INTRODUCTION
Parkinson’s disease (PD) is one of the most prevalent progressive neurodegenerative diseases, second only to Alzheimer’s disease (AD). Both PD and AD are progressive through the years and show characteristic but complex histopathological features in brain tissues [1]. The clinical manifestations of PD and AD are not the same; however, the molecular pathogenesis of them overlap [2]. Thus, study on PD is not only important to PD itself, but also helpful to the treatment of AD. PD is characterized by the loss of dopaminergic neurons in substantia nigra of midbrain, leading to movement dysfunction [3]. Besides motor deficits, other disorders, such as cognitive decline, mood disturbance, sleep dysfunction, gastrointestinal problems and dysautonomia, also appears in the later stage [4]. The incidence of PD increases with age, affecting up to 1% of people over the age of 60 worldwide [5].
Accumulating evidence indicates that abnormal neuroinflammation plays key roles in the progression of PD [6]. Activated microglia were reported to release a series of pro-inflammatory factors, contributing to dopaminergic neurodegeneration [7, 8]. The PI3K/Akt signaling pathway was considered crucial for regulating neuroinflammation and dopaminergic neuron loss [9, 10]. A recent study using PD mouse models identified PI3K/Akt as an important regulator in mitigating inflammatory responses and thus protecting dopaminergic neurons [11]. Furthermore, PI3K/Akt was reported important for microglia activation and neuroinflammation [12].
Chinese traditional medicine, with low toxicity, were reported to have neuroprotective effects. Some of them have been used in the clinical treatment of PD [13]. Pharmaceuticals-induced suppression of inflammation could ameliorate PD degree in mice and cells [14–16]. Bergenin (C14H16O9; Berg), an isocoumarin compound isolated from the herb of Saxifrage stolonifera Curt, which has been widely used in China for many years, possesses hepatoprotective, anti-tumor, anti-inflammatory, antioxidative, and neuroprotective effects [17–20]. For instance, Barai et al. revealed that Berg (40 mg/Kg and 80 mg/Kg) exerted neuro-modulatory activity, cholinesterase inhibition, antioxidant activity, and anti-dementia effects in AD models, both in vivo and in vitro [20]. Hironobu et al. synthesized a series of Berg derivatives and showed that most of them exhibited high antioxidant activity and neuroprotective activity on rat cortical neurons [17]. Jung et al. demonstrated that synthetic Berg compounds showed potent anti-inflammatory effects by inhibition of NO expression [21]. However, whether Berg could exert anti-inflammatory and neuroprotective effects on PD remains unknown. Thus, we aimed to explore the role of Berg played in PD progression and the underlying mechanisms.
In this study, we showed that Berg could rescue the movement dysfunctions and the loss of dopaminergic (DA) neurons in MPTP-induced PD mice, and exerted neuroprotective effects by suppressing abnormal neuroinflammation. Specifically, the PI3K/Akt signaling pathway might be involved in the underlying mechanism.
MATERIALS AND METHODS
Animals
Seven- to eight-week-old male C57BL/6 mice (∼20 g) were obtained from Vital River Experimental Technology Company (Beijing, China). Mice were randomly divided into four groups (n = 6 each group): control group (saline-treated group), MPTP-treated group, MPTP + Berg (L) group, and MPTP + Berg (H) group. Mice in the control group were injected with saline intraperitoneally daily for 14 days. Mice in MPTP group were injected with 30 mg/Kg MPTP once a day for 5 days to induce PD symptoms [22], then treated with saline for 9 days. Mice in MPTP + Berg (L) and MPTP + Berg (H) groups were injected with 30 mg/Kg MPTP once a day for 5 days, and then treated with Berg at a dosage of 40 mg/kg/d and 80 mg/kg/d, respectively, for 9 days [20]. All animal experimental procedures were approved by the Laboratory of Animal Ethical Committee of The First Affiliated Hospital of Zhengzhou University.
Behavioral tests
Rotarod test
Rotarod test was performed as previously described [23]. In brief, mice were placed on the rotating bar of the rotarod unit. The speed was set at 25 rpm, and the time that each mouse stayed on the rotating bar was recorded.
Hanging test
As previously described [24], a horizontal grid was used for mice to grip on. Then the grid was made upside down. The time that each mouse hanging on it until fall down was recorded.
Narrow beam walking test
As previously described [24], a stationary wooden narrow flat beam (1 cm) positioned at a 100 cm above the floor was used for mice to walk. The time that each mouse walk from one end to another was recorded.
Cell culture
BV2 cells (mouse microglia) and PC12 cells (mouse adrenal pheochromocytoma cell line) were obtained from Institute of Basic Medical Science, Chinese Academy of Medical Sciences. Cells were subjected to Dulbecco’s Modified Eagle Medium (DMEM) plus with 10% Fetal Bovine Serum (FBS) and incubated in a humidified atmosphere of 5% CO2 at 37°C.
Real-time PCR
Total RNA was isolated from tissues or cells using Trizol reagent (Invitrogen, USA) according to the manufacturer’s instructions. After digestion with DNase (Promega, Madison, USA), the RNA was reverse transcribed into cDNA using M-MLV Reverse Transcriptase (Promega, Madison, USA). The cDNA was then used for real-time PCR by SYBR green (Takara, Japan). The internal control was β-Actin. Primers for real-time PCR were presented in Table 1.
Sequences of real-time PCR primers
Western blot
Total protein was extracted from mid brain tissues or cells using RIPA lysis buffer (Beyotime, Beijing, China). Equal amounts of protein from each sample were separated electrophoretically on SDS-polyacrylamide gels, and transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, Boston, USA). 5% non-fat milk was used to block the membranes for 2 h, and then the membranes were incubated with primary antibodies: rabbit anti-tyrosine hydroxylase (TH) (1:1000; Enzo Life Sciences, USA), rabbit anti-iNOS (1:500, Wuhan, Hubei), rabbit anti-p-PI3K (1:1000; Millipore, USA), rabbit anti-PI3K (1:1000; Millipore, USA), mouse anti-p-AKT (1:1000; Abcam, UK) and mouse anti-AKT (1:1000; Abcam, UK), and mouse anti-β-Actin antibody (1:2000, Proteintech, USA) overnight at 4°C. After washing with TBST, the membranes were then incubated with corresponding secondary horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. The signal was detected using a chemiluminescence detection kit (PerkinElmer Life Sciences, Boston, MA, USA).
MTT assay
BV2 cells were incubated in 6-well plates for four groups: control group, LPS group, LPS + Berg (L) group and LPS + Berg (H) group. Cells in LPS group, LPS + Berg (L) group and LPS + Berg (H) group were treated with LPS at a final concentration of 0.5μg/ml, and 6 h later, Berg was added into LPS + Berg (L) group and LPS + Berg (H) group at final concentrations of 15μM and 30μM, respectively [20, 21]. Twenty-four hours later, we collected the culture supernatants (CS) as control CS, LPS CS, LPS + Berg (L) CS, and LPS + Berg (H) CS. Then PC12 cells were seeded in a 96-well plate. After 6 h, the culture supernatant of PC12 cells was removed, and control CS, LPS CS, LPS + Berg (L) CS, and LPS + Berg (H) CS were added instead. Twenty-four hours later, MTT assay was performed to detect the cell viability. In short, MTT was added into the 96-well plate at the final concentration of 5 mg/ml. After an incubation of 4 h, the cell supernatant was replaced with DMSO, and a microplate reader was used to detect the optical density.
Flow cytometry
The PC12 cells were subjected to culture supernatant from BV2 cells as previously described. Forty-eight hours later, the cells were gathered and treated with Annexin V-FITC Apoptosis Detection Kit (Dojindo Laboratories, Shanghai, China), according to the manufacturer’s instructions. A flow cytometer (BD, Franklin Lakers, NJ, USA) was used wo detect the apoptosis of cells.
Immunohistochemistry and immunofluorescence assays
The brain tissues were fixed with 4% paraformaldehyde for 24 h, and soaked in 0.05 M PBS containing 30% sucrose for cryoprotection. Frozen brains were cut into 25μm coronal sections. Anti-TH antibody (Cell signaling technology, USA) was used for immunohistochemical staining and Anti-Iba1 antibody (Cell signaling technology, USA) was used for immunofluorescence staining. A microscope (Olympus, Tokyo, Japan) and a laser scanning confocal microscope were used to observe the positive cells. Six visual fields were chosen randomly for each sample.
Assay for striatal DA and its metabolites
On day 14, the right anterior striata were dissected. DA and DOPAC were detected by HPLC with electrochemical detection as previously described [25].
Statistical analysis
All data were expressed as mean±standard deviation (SD). The differences between two groups were analyzed using student’s two-tailed t-test and the differences among multiple groups were analyzed using one-way analysis of variance (ANOVA). p < 0.05 was considered statistically significant.
RESULTS
Berg mitigated MPTP-induced behavior disorder
To investigate the effect of Berg on motor dysfunction induced by MPTP, the rotarod test, hanging test and narrow beam test were assessed. As shown in Fig. 1, MPTP treatment decreased the time of rotarod test and hanging test, and increased the time of narrow beam test, which confirmed that the PD mouse model was established successfully. Additionally, Berg administration mitigated the changes induced by MPTP, indicating that Berg could alleviate the behavior disorder of MPTP-induced PD mice.

Effect of Berg on MPTP-induced behavioral parameters. Rotarod test (A), hanging test (B), and narrow beam test (C) were carried out to measure the degree of motor deficits in mice treated with MPTP or/ and Berg. **p < 0.01, compared with control group; # #p < 0.01, compared with MPTP group.
Berg alleviated the dopaminergic neuron damage in MPTP-induced PD mice
To explore the neuroprotective effect of Berg on MPTP-induced PD mice, TH (a key enzyme of dopamine), dopamine, and DOPAC were detected. TH-immunoreactivity was decreased in MPTP-induced mice, and Berg treatment prevented the reduction in a dose dependent way (Fig. 2A). Western blot was also performed to detected the protein expression level of TH. Consistently, the TH protein level was decreased in MPTP-induced mice, and Berg treatment inhibited the reduction (Fig. 2B). Furthermore, the expression levels of dopamine and DOPAC (a normal product of the metabolism of dopamine) were measured, and the results showed that the MPTP injection decreased the levels of dopamine and DOPAC. Berg treatment mitigated the reduction (Fig. 2C, D). Together, these results indicated that Berg could ease the dopaminergic neuron damage induced by MPTP.

Berg eased the dopaminergic neuron damage and increased the contents of dopamine and its metabolites in MPTP-induced mice. The expression level of TH was measured by immunohistochemistry (A) and western blot (B). The expression levels of dopamine (C) and DOPAC (B) were detected by HPLC analysis. **p < 0.01, compared with control group; # #p < 0.01, compared with MPTP group.
Berg attenuated inflammatory reactions of glia cells in MPTP-induced PD mice
As activation of microglia cells is involved in neuroinflammation, the expression level Iba1 (a marker of activated microglia) was detected by immunofluorescence staining. The results showed that the number of Iba-1-positive cells was increased upon MPTP injection, and Berg treatment attenuated this change (Fig. 3A, B). Furthermore, we measured the levels of pro-inflammatory mediators and found that MPTP injection significantly increased the levels of iNOS, TNF-α, IL-1β, and IL-10, while Berg treatment suppressed the increasement in a dose-dependent manner (Fig. 3C–F). These results indicated that Berg could mitigated the inflammatory cascade in MPTP-induced PD mice.

Berg alleviated microglia activation and inflammatory reactions in MPTP-induced mice. A) The activation of microglia was detected by Iba-1 immunostaining. The expression levels of inflammatory factors NO (C), TNF-α (D), IL-1β (E). and IL-6 (F) were examined. **p < 0.01, compared with control group; # #p < 0.01, compared with MPTP group.
Berg activated the PI3K/Akt signaling pathway in MPTP-induced PD mice
To further identify the mechanism underlying Berg-mediated neuroprotection, we performed western blot to examine the protein expression levels of p-PI3K and p-Akt. As shown in Figure 4, MPTP injection decreased the protein levels of p-PI3K and p-Akt, while Berg induced a significant increasement compared to the MPTP mice. These results suggested that Berg induced the activation of PI3K/Akt signaling pathway in MPTP-treated PD mice.
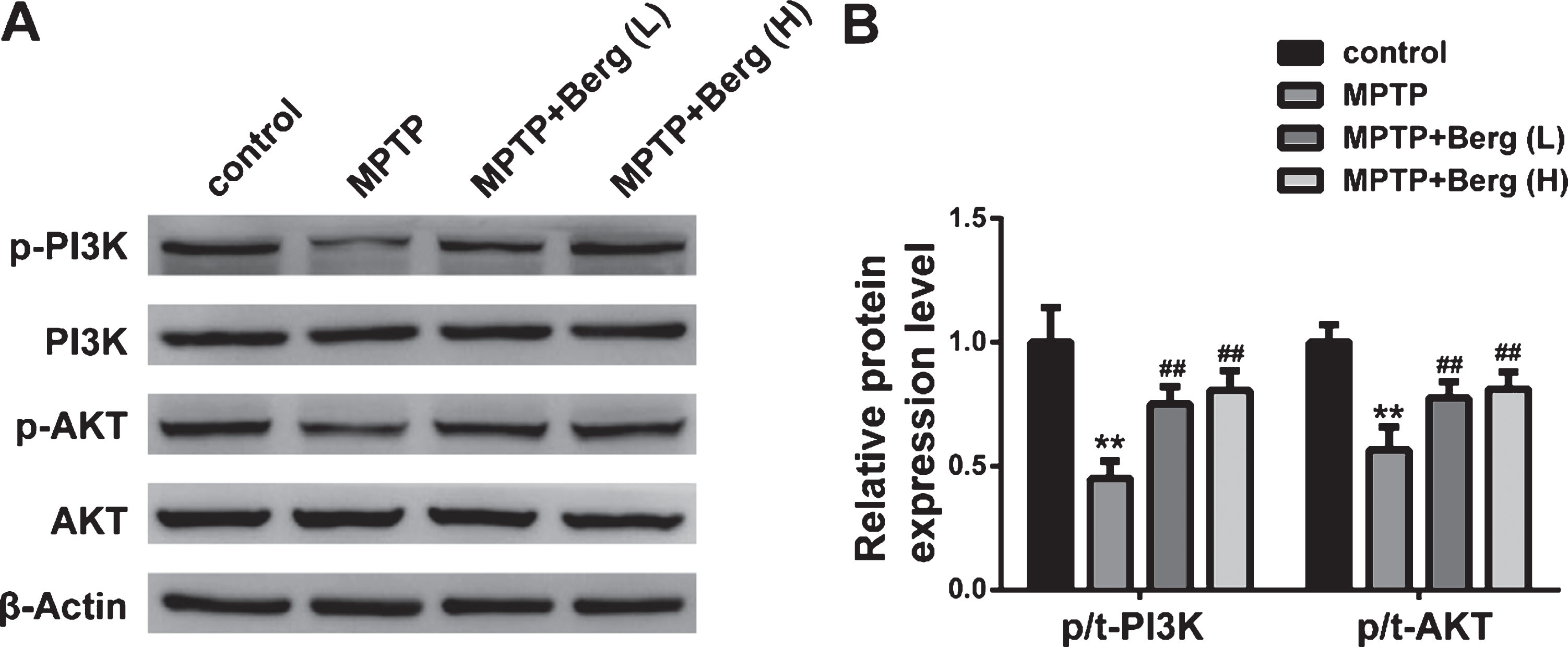
Berg activated the PI3K/Akt signaling pathway in MPTP-induced mice. A) The protein expression levels of p-PI3K, PI3K, p-Akt, Akt, and β-Actin were detected by western blot. B) Quantitative analysis of p-PI3K normalized to PI3K and p-Akt normalized to Akt. **p < 0.01, compared with control group; # #p < 0.01, compared with MPTP group.
Berg alleviated inflammation in LPS-treated BV2 cells
BV2 cells were used as the major model for an in vitro study. As shown in Fig. 5, compared to control, the expression levels of NO, TNF-α, IL-1β, and IL-10 were upregulated by LPS treatment. However, Berg suppressed the increasement of these inflammatory mediators. The results demonstrated that Berg could inhibit the production of inflammatory mediators in vitro.
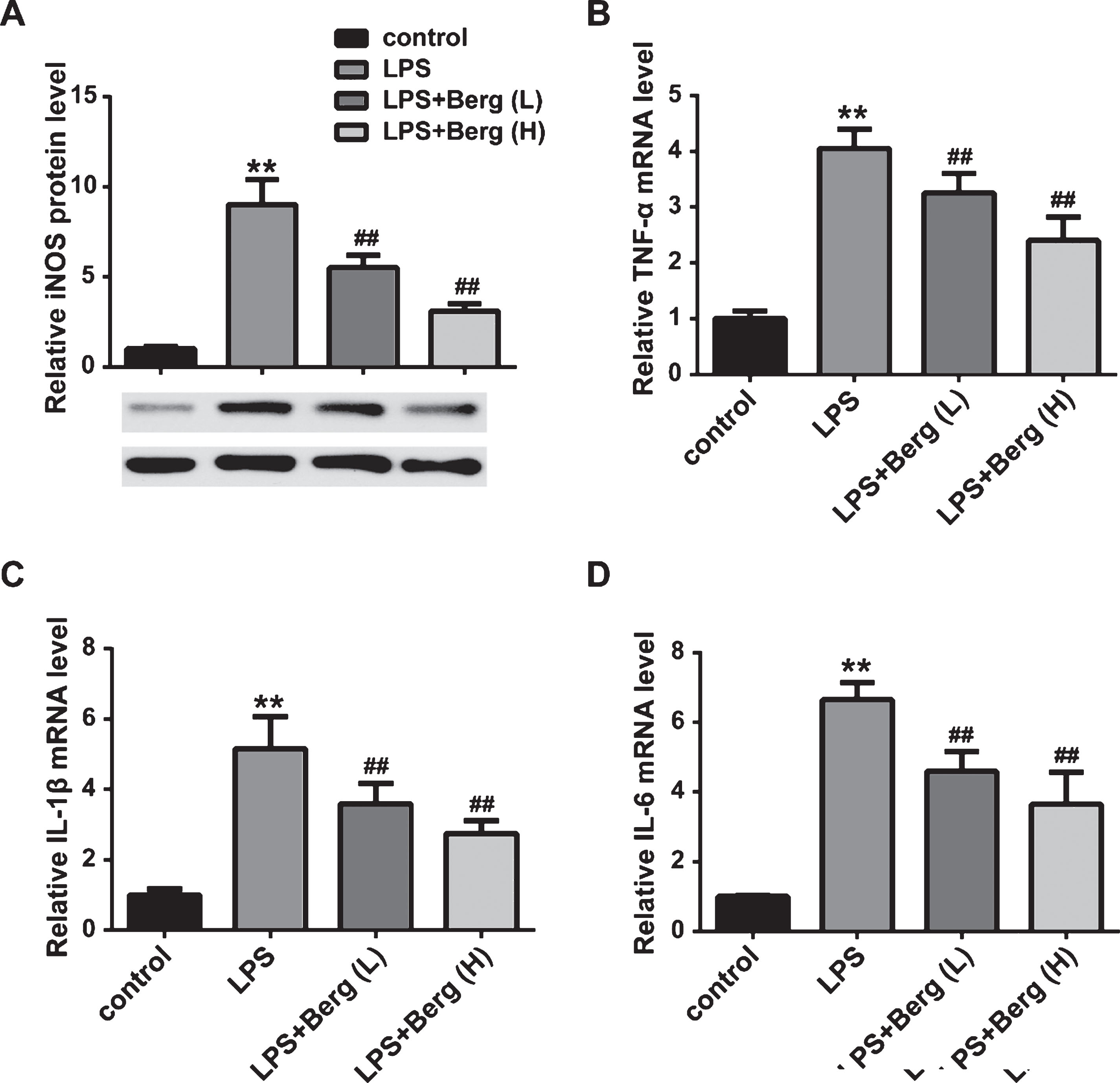
Berg inhibited inflammatory reactions in LPS-treated BV2 cells. A) The NO content in culture supernatant of BV2 cells was detected. The mRNA expression levels of TNF-α (B), IL-1β (C), and IL-6 (D) were measured by real-time PCR. **p < 0.01, compared with control group; # #p < 0.01, compared with MPTP group.
Berg activated the PI3K/Akt signaling pathway in LPS-treated BV2 cells
Similar to the results in vitro, the protein expression levels of p-PI3K and p-Akt were downregulated in LPS-treated BV2 cells, compared to the control group. In contrast, treatment with Berg, following MPTP injection, induced an increasement in p-PI3K and p-Akt protein levels compared to MPTP group. These results suggested that Berg could induce the activation of PI3K/Akt signaling pathway in vitro.
Berg attenuated the microglia-mediated cytotoxic effect in vitro
It is known that microglia play important roles in inflammation of neurodegenerative diseases, and we demonstrated that Berg alleviated inflammation in vivo and in vitro (Figs. 3 and 5). Thus, we investigated whether Berg attenuated microglia-mediated cytotoxicity to neurons. Culture supernatant (CS) gathered from BV2 cells treated with nothing, LPS, LPS + Berg (L), and LPS + Berg (H) were used to maintain PC12 cells. After 48 h incubation, we found that the LPS treatment reduced the viability and promoted the apoptosis of PC12 cells, while Berg administration mitigated the alterations. The results indicated that Berg could suppress the toxicity from LPS-activated BV2 cells on neuronal cells.
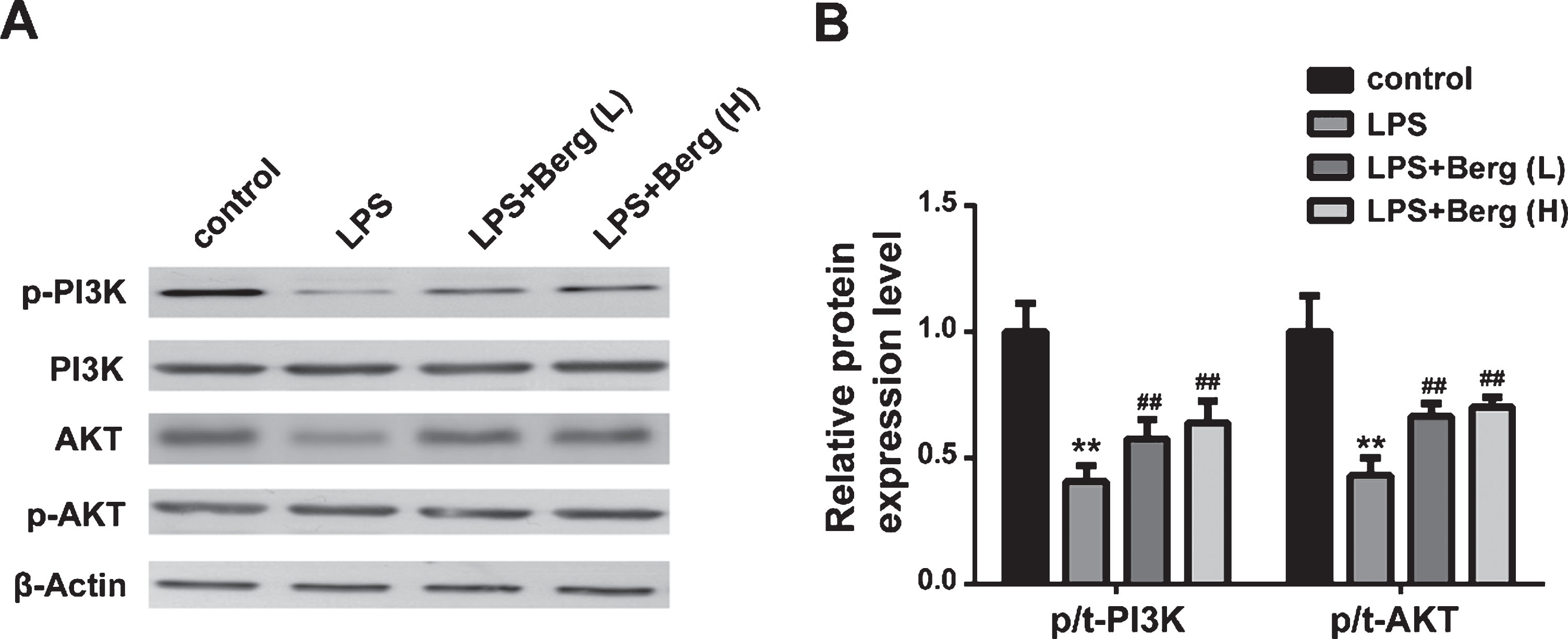
Berg induced the activation of PI3K/Akt signaling pathway in LPS-treated BV2 cells. A) The protein expression levels of p-PI3K, PI3K, p-Akt, Akt, and β-Actin were detected by western blot. B) Quantitative analysis of p-PI3K normalized to PI3K and p-Akt normalized to Akt. **p < 0.01, compared with control group; # #p < 0.01, compared with MPTP group.
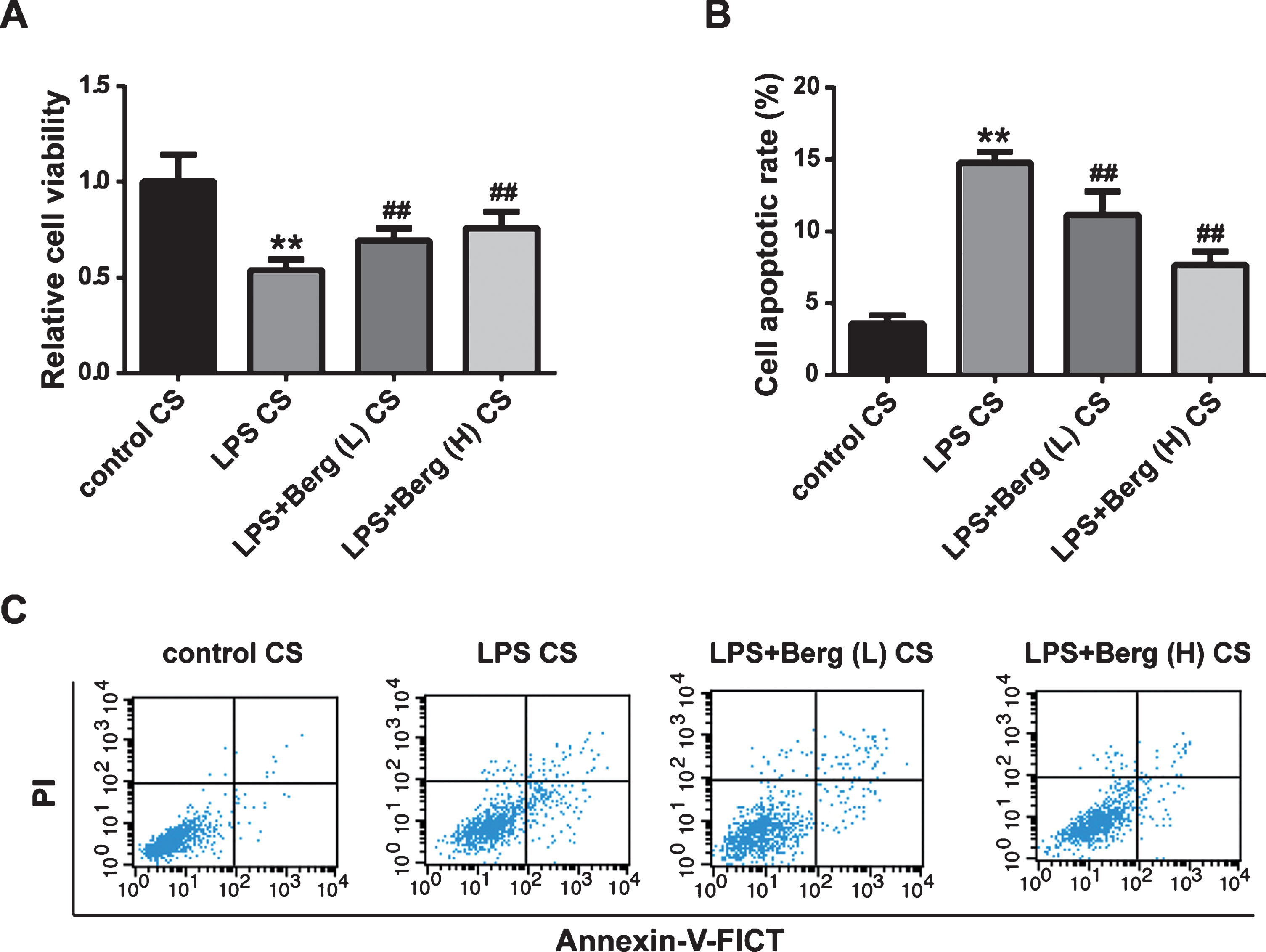
Berg attenuated the microglia-mediated cytotoxic effect on PC12 cells. A) The viability of PC12 cells was detected by MTT assay. B, C) The apoptotic rate of PC12 cells with different treatments were detected by flow cytometry. **p < 0.01, compared with control CS group; # #p < 0.01, compared with LPS CS group.
DISCUSSION
PD is a common neurodegenerative disease characterized by disabling dyskinesias and the loss of DA neurons in the substantia nigra. In PD research, several chemicals were commonly used to establish in vivo and in vitro PD models, among which MPTP was one of the most chemical used to introduce PD symptoms in vivo [26]. In vitro, LPS is a toxin that commonly used to stimulate cellular PD events [27]. LPS administration of BV2 cells could introduce neurodegeneration, microglia activation, and neuroinflammation [28]. In this study, we used MPTP-induced mice and LPS-induced BV2 cells to establish PD models.
Although the cause of PD remains elusive, it is considered that microglial-derived inflammation contributed to the pathophysiology of PD, as well as AD [29–31]. Physiologically, microglia could defend against microorganism invasion and thus are beneficial to neuronal function [32]. However, under pathological conditions, microglia are over-activated and release pro-inflammatory cytokines, such as iNOS, TNF-α, IL-1β, and IL-6, leading to neuronal cell death and neurodegeneration [33]. Also, suppression of microglia activation could attenuate neuronal inflammation [34]. In our study, MPTP treatment activated Iba-1-positive microglia in mice, and Berg alleviated this alteration. Moreover, the expression levels of pro-inflammatory cytokines were upregulated in MPTP-induced PD mice or LPS-induced cells, and Berg treatment attenuated these alterations. Therefore, we speculated that Berg might mitigate the loss of DA neurons by inhibiting the activation of microglia and the expressions of pro-inflammatory cytokines.
Berg, a colorless crystalline isocoumarin primarily obtained from Bergenia species, was reported to possess a series of health benefits, including hepatoprotective, anti-tumor, anti-inflammation, and neuroprotective properties in particular [35]. For instance, Gao et al. showed that Berg exerted anti-inflammatory roles via inhibiting MAPK and NF-κB signaling pathways in a mouse model of LPS-induced mastitis [36]. Shi et al. demonstrated that Berg could induce cell apoptosis, cell cycle arrest and suppress STAT3 signaling pathway in cervical cancer [18]. Of note, Berg was reported to exhibit neuroprotective activity by attenuating oxidative stress and restoring hippocampal cytoarchitecture in AD rat models [20]. Berg compounds suppressed NO generation, thus exerted anti-inflammatory activity on mice model [21]. Berg derivatives exhibited strong antioxidant effects and prevented neuronal death in vitro [17]. However, to the best of our knowledge, the role of Berg in PD animal models has not been explored before. Therefore, we investigated the function of Berg on PD. Our data demonstrated that Berg could alleviate the loss of DA neurons and inflammation in vivo and in vitro.
The PI3K/AKT signaling pathway was reported to play important roles in cell growth, proliferation, and survival under physiological conditions and could initiate inflammation [37]. For example, vitexin protected dopaminergic neurons by activating PI3K/Akt pathway in a MPTP-induced PD model [38]. Acacetin activated PI3K/AKT signaling pathway and attenuated neuronal cell apoptosis in a PD model of SH-SY5Y cells [39]. Zhao et al. reported that glutamine served as protective roles in a PD cell model via suppressing he PI3K/Akt signaling pathway [37]. Zhu et al. demonstrated that downregulation of DEC1 promoted MPP+-induced neurotoxicity via inhibiting PI3K/Akt/GSK3β pathway in PD mice model [40]. Additionally, Jia et al. demonstrated that PI3K/AKT/FoxO3a signaling pathway exerted neuroprotective effects in in vitro and in vivo models of PD [41]. These previous reports indicated that PI3K/Akt signaling pathway contributed to the neuroprotection in PD. In our study, to further explore the underlying mechanism of the neuroprotective effect of Berg on PD, we examined the PI3K/Akt signaling pathway. The results showed that the PI3K/Akt signaling was inhibited in MPTP-induced PD mice and LPS-induced BV2 cells. However, Berg could attenuate the alterations, indicating that the PI3K/Akt signaling pathway was involved in the mechanism.
Conclusion
Taken together, our findings revealed that Berg alleviated motor dysfunctions and neurodegeneration in MPTP-induced PD mice, and attenuated the inflammation in vivo and in vitro (Fig. 8). The neuroprotective effects of PD might be via the PI3K/Akt signaling pathway. Our study provided Berg as a therapeutic drug candidate for PD treatment.
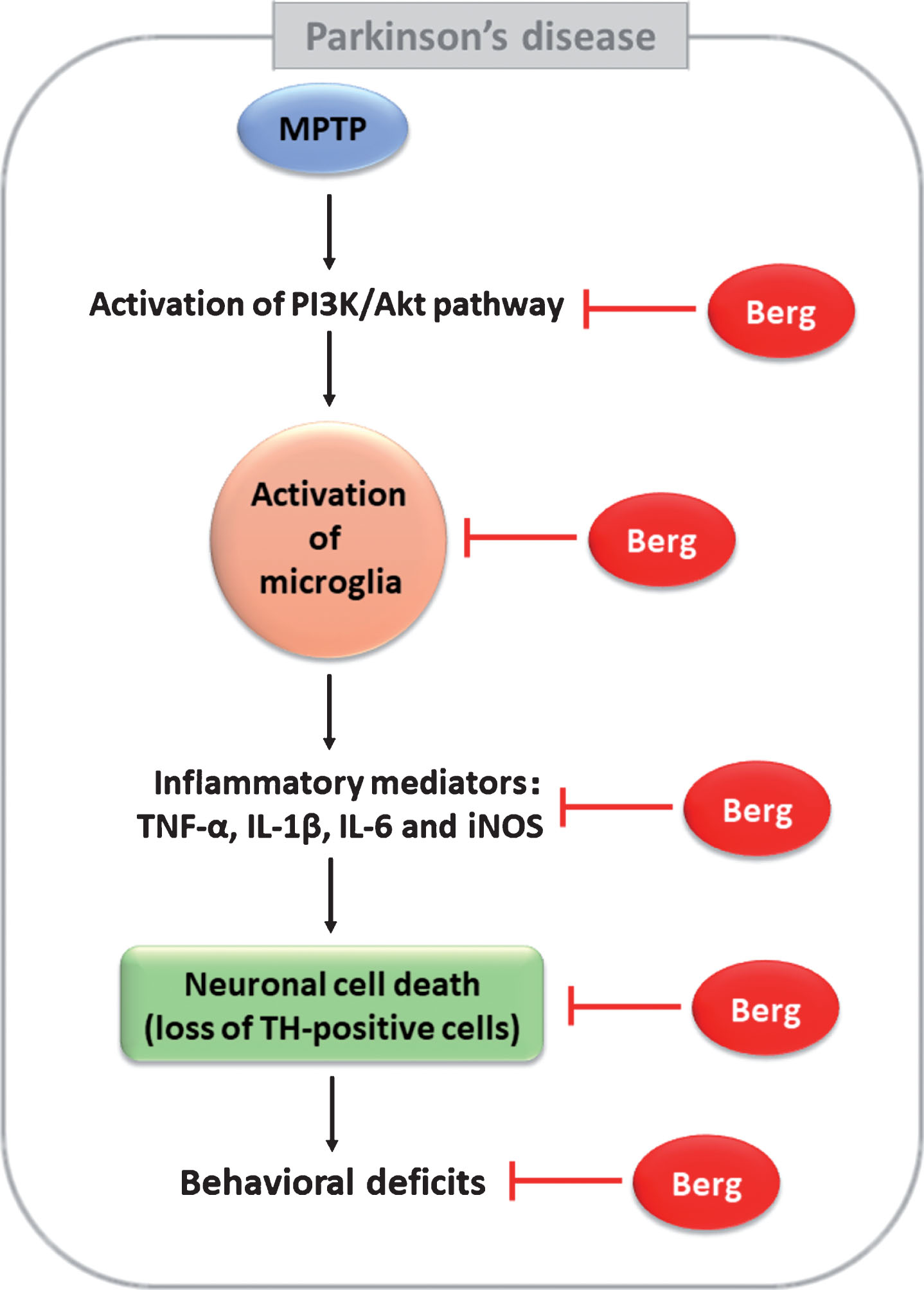
A diagram showing the proposed mechanisms of Berg in MPTP-induced PD. Berg could protect neuronal death induced by MPTP intoxication via regulation of PI3K/Akt signaling pathway and anti-inflammatory activities.