
Abstract
Background:
Alzheimer’s disease (AD) is a multifactorial disease, that involves neuroinflammatory processes in which microglial cells respond to “damage signals”. The latter includes oligomeric tau, iron, oxidative free radicals, and other molecules that promotes neuroinflammation in the brain, promoting neuronal death and cognitive impairment. Since AD is the first cause of dementia in the elderly, and its pharmacotherapy has limited efficacy, novel treatments are critical to improve the quality of life of AD patients. Multitarget therapy based on nutraceuticals has been proposed as a promising intervention based on evidence from clinical trials. Several studies have shown that epicatechin-derived polyphenols from tea improve cognitive performance; also, the polyphenol molecule N-acetylcysteine (NAC) promotes neuroprotection.
Objective:
To develop an approach for a rational design of leading compounds against AD, based on specific semisynthetic epicatechin and catechin derivatives.
Methods:
We evaluated tau aggregation in vitro and neuritogenesis by confocal microscopy in mouse neuroblastoma cells (N2a), after exposing cells to either epicatechin-pyrogallol (EPIC-PYR), catechin-pyrogallol (CAT-PYR), catechin-phloroglucinol (CAT-PhG), and NAC.
Results:
We found that EPIC-PYR, CAT-PYR, and CAT-PhG inhibit human tau aggregation and significantly increase neuritogenesis in a dose-dependent manner. Interestingly, modification with a phloroglucinol group yielded the most potent molecule of those evaluated, suggesting that the phloroglucinol group may enhance neuroprotective activity of the catechin-derived compounds. Also, as observed with cathechins, NAC promotes neuritogenesis and inhibits tau self-aggregation, possibly through a different pathway.
Conclusion:
EPIC-PYR, CAT-PYR, CAT-PhG, and NAC increased the number of neurites in Na2 cell line and inhibits tau-self aggregation in vitro.
Keywords
INTRODUCTION
Alzheimer’s disease (AD)
AD is the most frequent form of dementia among the elderly population [1]. This disease affects the processes of memory and learning, along with behavioral aspects and mood of the patients. The World Health Organization has estimated that around 50 million people suffer from AD. Recently, our laboratory has associated this pathology with early behavioral disorders that occur previous to cognitive decline [2]. This brain disorder is increasing at an alarming rate, constituting a major puzzle for the medical community and for society as well. According to the state of the art, two main etiological effectors have been reported: 1) Neurofibrillary tangles, derived from the progressive aggregation of the hyperphosphorylated protein tau, inside neurons and, that assembles into oligomeric structures named “paired helical filaments” [3 –6] and 2) senile plaques, composed by deposits of the amyloid-β (Aβ) peptide of 39 to 42 aminoacidic residues, generated by the proteolytic excision of the amyloid-β protein precursor (AβPP) as a result of the action of β- and γ-secretases. Senile plaques are accumulated in the extracellular space, promoting loss of synaptic processes and neuronal death [1, 7].
Innovative treatments are essential to improve the quality of life and ameliorate the symptoms of AD patients and those at risk of AD. However, the pharmaceutical industry has not yet provided effective pharmacotherapies to control or prevent AD. In this context, major attention has been given to natural bioactive compounds, such as polyphenols from plants [8]. Among them, epicatechin and catechin derived polyphenols from tea and other natural sources, have shown efficacy in preventing cognitive decline in elderly individuals in several clinical trials [9].
Current hypotheses on AD pathogenesis
We proposed that the onset of AD is a consequence of the chronic activation of microglial cells to different “damage signals”, including pathological tau oligomers, which triggers a neuro-inflammatory response. The latter promotes tau hyperphosphorylation, leading to a misfolding of this protein with the consequent oligomerization and structural alterations in the neuronal cytoskeleton [5 , 11]. In this context, we postulated the neuroimmunomodulation hypothesis of AD [12]. Innate immunity is directly involved in AD, thus neuroinflammation occurs as a result of damage signals which activate microglial cells to induce over production of cytokines that are finally responsible of the reactivation of protein kinases like CDK5 that determines tau hyperphosphorylation. This chain of brain reactions affects the neuroimmunomodulation patterns [12]. The alterations leading to tau phosphorylation followed by oligomerization of this protein constitutes a major paradigm of the etiopathogenesis of AD, in a similar fashion as observed in other protein misfolding disease that involve different protein systems: e.g., huntingtin in Huntington’s disease, α-synuclein in Parkinson’s disease, prion protein in spongiform encephalopathy, etc. It is likely that the neuroinflammatory response leads to an important loss of neurons, in contrast with the first action of damage signals [13]. In this context, neuroinflammation is a key element in triggering several different neuropathologies [14], including AD. Thus, AD is related to the chronic inflammatory condition associated with the progressive death of neurons and brain atrophy that involves the release of pathological forms of tau protein to the extracellular environment. Considering that tau oligomers are capable of activating microglia, they trigger a positive feedback mechanism, generating a constant damage to cells [10 , 16]. Therefore, therapies aimed to control tau oligomers and ameliorate neuroinflammation have an enormous value in AD treatment, particularly in view of the fact that high levels of inflammatory mediators are present in the vicinity of the paired helical filaments of AD patients [17].
Chronic metabolic diseases such as hypertension, diabetes, and clinical depression are considered silent contributors to chronic neuroinflammation [18]. Other risk factors causing functional impairment or even cellular death in the central nervous system (CNS) tissue are stroke, atherosclerosis, and the presence of senescent cells. Moreover, during normal aging, there is a natural chronic activation of pro-inflammatory signals in the same areas, contributing to an even higher vulnerability for neuropsychiatric disorders [19]. Pro-inflammatory agents, such as TNFα and others, correlate with clinical depression [18, 20].
Emerging therapeutic strategies to tackle AD
New drugs and vaccines are currently being proposed to effectively treat AD, most of them without clinical success. In this report, we propose a path to revisit the design of bioactive compounds against AD based on epicatechin-derived polyphenols and NAC in the same context. Several nutraceutical compounds have been analyzed with respect to their ability to control tau self-aggregation, a major culprit of AD pathogenesis, including the “Andean Compound” rich in fulvic acid and rosemary extracts [21, 22]. As was described previously, important evidence positions nitroxidative stress in aging and neurodegenerative diseases [23]. N-acetylcysteine (NAC) is a potent antioxidant, known as a glutathione (GSH) precursor, with the capacity to cross the blood-brain barrier [24, 25]. NAC has been mainly used against acetaminophen intoxication and contrast-induced nephropathy, besides to demonstrate prevention of cellular damage in different models and diseases, including Parkinson’s disease, Huntington’s disease, and others [26, 27]. Moreover, NAC has shown anti-inflammatory effect in CNS, promoting axonal sprouting, reducing microglial pro inflammatory activation, and increasing cell survival rate after spinal cord injury [28, 29].
As it was previously described, cognitive impairment and memory loss are hallmarks of AD. Recently, NAC treatment was found to prevent the cognitive impairment induced by streptozotocin (STZ), avoiding synaptic plasticity loss in a mouse model [30]. Also, it significantly improved memory retention and attenuation of oxidative damage after STZ injections in rats [31]. Similarly, NAC was able to reduce memory impairment caused by Aβ-induced damage in rats [32]. NAC can also improve the deficits in conditioned place preference consolidation after toxicity induced by methamphetamines in mice [33], and ameliorate memory impairment in SAMP8 aged mice [24]. Thus, NAC is a potential candidate molecule for treatment of AD. Nevertheless, the effects of NAC on tau protein have been poorly described, and will be evaluated in this work. Following the previous aim of searching for nutraceutical compounds for AD treatment, our group produced a series of semisynthetic catechin and epicatechin-derived adducts from natural proanthocyandins extracted from avocado peels and Peumus boldus leaves [34, 35]. Interestingly, these molecules decreased the secretion of pro-inflammatory cytokine IL-8 in AGS cells. Hence, these compounds could be used in other pathologies where inflammation is relevant. Here, we present a series of anti-inflammatory/antioxidant catechin-derived polyphenols and NAC as potential neuroprotective agents using a neuroblastoma N2a cells model system. These polyphenols were also tested as in their capacity to controlling human tau aggregation.
MATERIALS AND METHODS
Mouse neuroblastoma cells
Mouse neuroblastoma cells (N2a) were cultured in 10.0 cm plates. Each plate contained a final volume of 10.0 ml of DMEM Full medium (Gibco) containing 10% FBS (Biological Industries) and 1% penicillin/streptomycin (Biological Industries). Cells were maintained for 48 h incubated at 37°C in a humidified 5% CO2 incubator, obtaining 80–90% confluence.
Flavan-3-ol-derived compounds
Semisynthetic flavan-3-ols were obtained were obtained by nucleophilic attack with phloroglucinol and pyrogallol according to our previous work [34]. Using this procedure, epicatechin-derived adducts were synthetized from avocado peel procyanidins, while catechin-derived adducts were synthetized from P. boldus procyanidins [36]. All compounds were isolated using repeated centrifugal partition chromatography runs (CPC) until 99.8% purity. Identity and purity were assessed by HPLC-ESI-MS/MS. All adducts shown a pseudo-molecular ion at m/z 413 [M–H] – and MS2 ion fragments at m/z 287.3 (cleavage of pyrogallol or phloroglucinol moieties), an ion fragment at m/z 269.3 (–18 amu loss). Heterocyclic ring fission produced ion fragments at m/z 161.3 and m/z 125.2, both characteristic of phenolic adducts in LC–MS/MS negative polarity [37].
Viability assay
N2a cells were seeded in 96-well plates with a confluence of 7000 cells/well. On the next day, cells were treated with variable concentrations of the bioactive compounds (in the range 1–15μM) for 24 h, at 37°C and 5% CO2. Bioactive compounds used were: catechin pyrogallol (CAT), catechin phloroglucinol (FLG), epicatechin pyrogallol (EPI), and N-acetyl cysteine (NAC). Control groups were: 1) Cells without the treatment; and 2) a blank control, in which dish wells contained the medium but no cells. We used the MTT assay for cell toxicity. The medium was replaced by fresh DMEM medium without phenol red, 10μl of MTT [12μM] were added and incubated for 4 h at 37°C and 5% CO2. 100μl of 5% SDS–HCl solution was added and incubated in the same conditions for 4 h and finally recording the absorbance at 550 nm using Sunrise plate reader.
Immunocytochemistry
N2a cells were plated on glass coverslips (12 mm diameter; 10000 cells/well). Three independent experiments were performed with three technical replicates each. After being exposed to experimental conditions, cells were washed with PBS and fixed with paraformaldehyde-sucrose (4%) for 30 min at 37°C. Cells were blocked with 5% BSA in PBS and then incubated with mouse alpha-tubulin 1 : 1500 (Invitrogen), TO-PRO 1 : 500 (Life Technologies), Alexa Fluor-488 phalloidin at 1 : 300 (Invitrogen), and goat anti-mouse AlexaFluor-555-conjugated 1 : 400 (Invitrogen). Slides were mounted in FluorSave fluorescence mounting media (Calbiochem) and images were collected with a Carl-Zeiss Laser Scanning Confocal Microscopy. The photographs obtained were representative of the different samples.
Tau protein purification
Recombinant tau protein purification was carried out as described [38, 39]. The sequence of htau 40 (longest isoform) was donated by Dr. Eckard Mandelkow (Hamburg, Germany). The tau protein fragment was cloned into pET-28ª vector to yield a His-tagged protein, while and htau 40 was expressed in Escherichia coli strain BL21(DE3). LB medium containing kanamycin was inoculated with a stationary overnight culture. The culture was grown at 37°C to OD600 of 0.5–0.6 and protein expression was induced by addition of 1 mM IPTG for 4 h. The cells were pelleted and sonicated. Recombinant tau was purified via a succession of Ni-Sepharose chromatography (equilibrated in 20 mM NaH2PO4, 500 mM NaCl, and 20 mM imidazole, pH 7.4, elution with buffer 200 mM imidazole) and side exclusion chromatography coupled to HPLC in a Proteema 100 column (PSS, Germany) with buffer 50 mM NaH2PO4, 300 mM NaCl, pH 6.5. The purity of the protein was verified on a Coomassie Brilliant Blue stained SDS-polyacrylamide gel (12%) and analyzed through densitometry by using software ImageJ [40]. The protein was concentrated and stored at –20°C until use. The concentration of purified tau was determined using the extinction coefficient at 280 nm (7700 M– 1cm– 1).
Tau aggregation assays
Aggregation of htau40 was performed by incubating htau40 in the concentration of 50μM in a final volume of 1 ml at 37°C in buffer PBS, pH 7.4 with the anionic cofactor heparin (molar ratio of tau to heparin = 4 : 1) for 10 days, with continuous shaking. In the case of control, the protein was incubated at the same concentration without the aggregating agent (Heparin). At day three of tau aggregation, in separate experiments the four natural compounds were added independently to a final concentration of 10μM, and 5.0 mM for NAC. In control experiments compounds were also added at zero time.
ThS fluorescence assay
The htau40 aggregation was monitored by fluorescence Thioflavin S (ThS) as described by Friedhoff [41] and Barghorn [42]. ThS to 6.25μM was added to the htau40 and heparin solution and incubated for 1 h at 37°C in the dark and stirring to allow binding of the fluorescent probe with tau beta-sheet structures formed. Fluorescence was measured on the computer Biotek Synergy 2 excitation at 440 nm and emission at 508 nm (excitation filter 440/30 and 508/20 emission filter), minus the basal fluorescence of blank incubated in the same conditions.
RESULTS
The morphology of N2a cells presents alterations after treatment with the bioactive compounds
While the different treatments present diverse effects, mostly all of them promote morphological changes in N2a cells, such as an increase in the number and length of neuritic processes and reorganization of the neuronal cytoskeleton (Fig. 1B). Figure 1A shows the fluorescence images for tubulin, actin, the nucleus, and merge for the control condition.
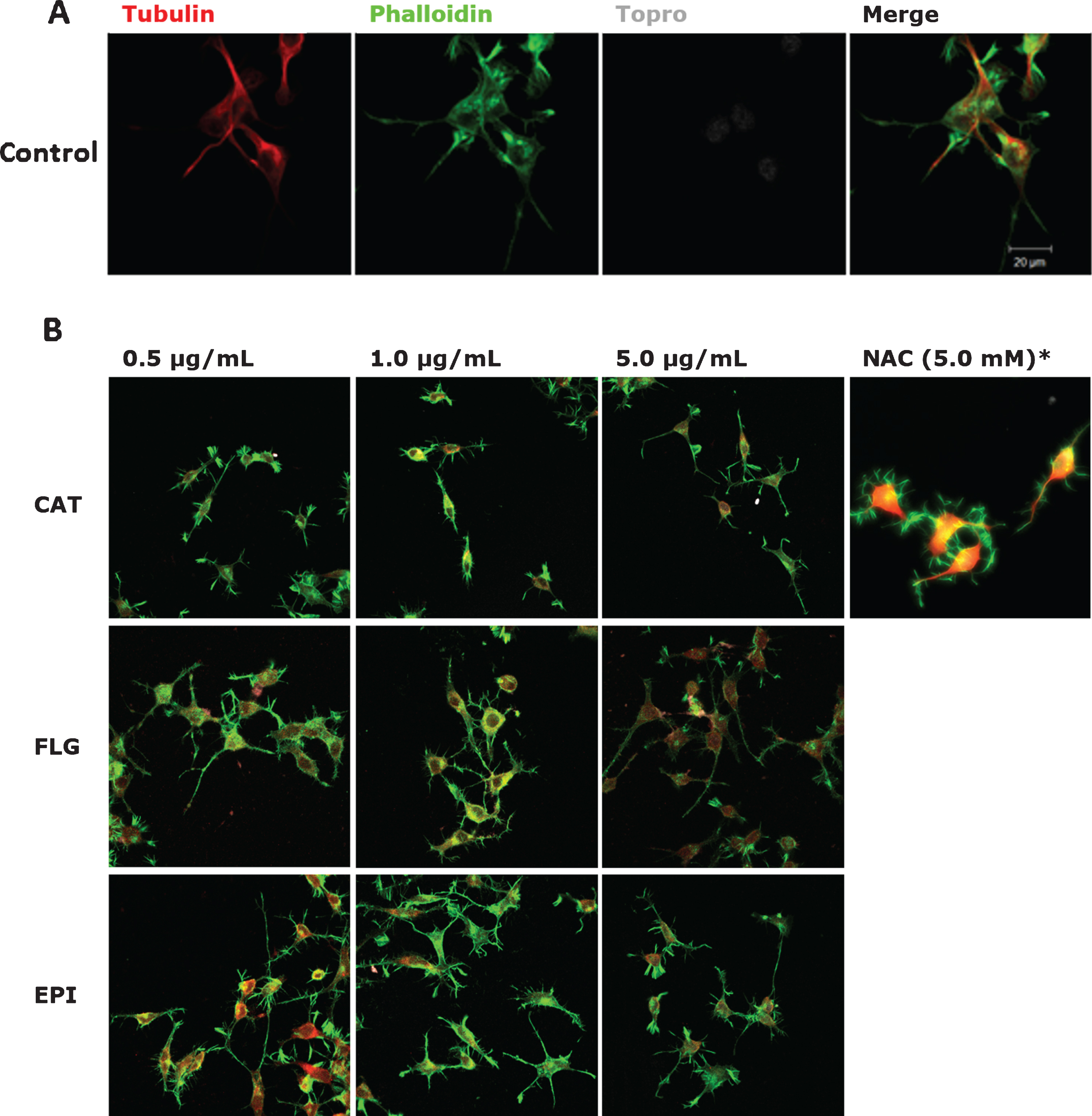
Bioactive compounds modify N2A cells morphology. The representative images after treatment with different concentrations of the bioactive compounds are shown. Immunofluorescence assays representing different assay conditions. A) Control condition. Protein markers used in the experiment are shown as follow; α-Tubulin, microtubule-associated protein marker; Phalloidin, β-actin protein marker; Topro, cell core and merge of all the channels involved. B) The representative images of N2a cells present their morphological changes after being treated with 0.5μg/mL; 1.0μg/mL, 5.0μg/mL, and 5.0 mM of the bioactive compounds catechin-pyrogallol (CAT), catechin-pholoroglucinol (FLG), epicatechinpyrogallol (EPI), and N-acetyl cysteine (NAC) for 24 h. N = 3.
Cell viability of N2a is not affected after treatments with the different bioactive compounds
At a global level, we found that under different conditions of bioactive compounds (Fig. 2A), cells did not present serious impairment in viability (Fig. 2B). In specific cases, such as NAC, higher concentrations over 25.0 mM of NAC were slightly toxic to cells. In general, all polyphenols investigated in the concentrations range indicated did not decrease viability as compared with the control condition.
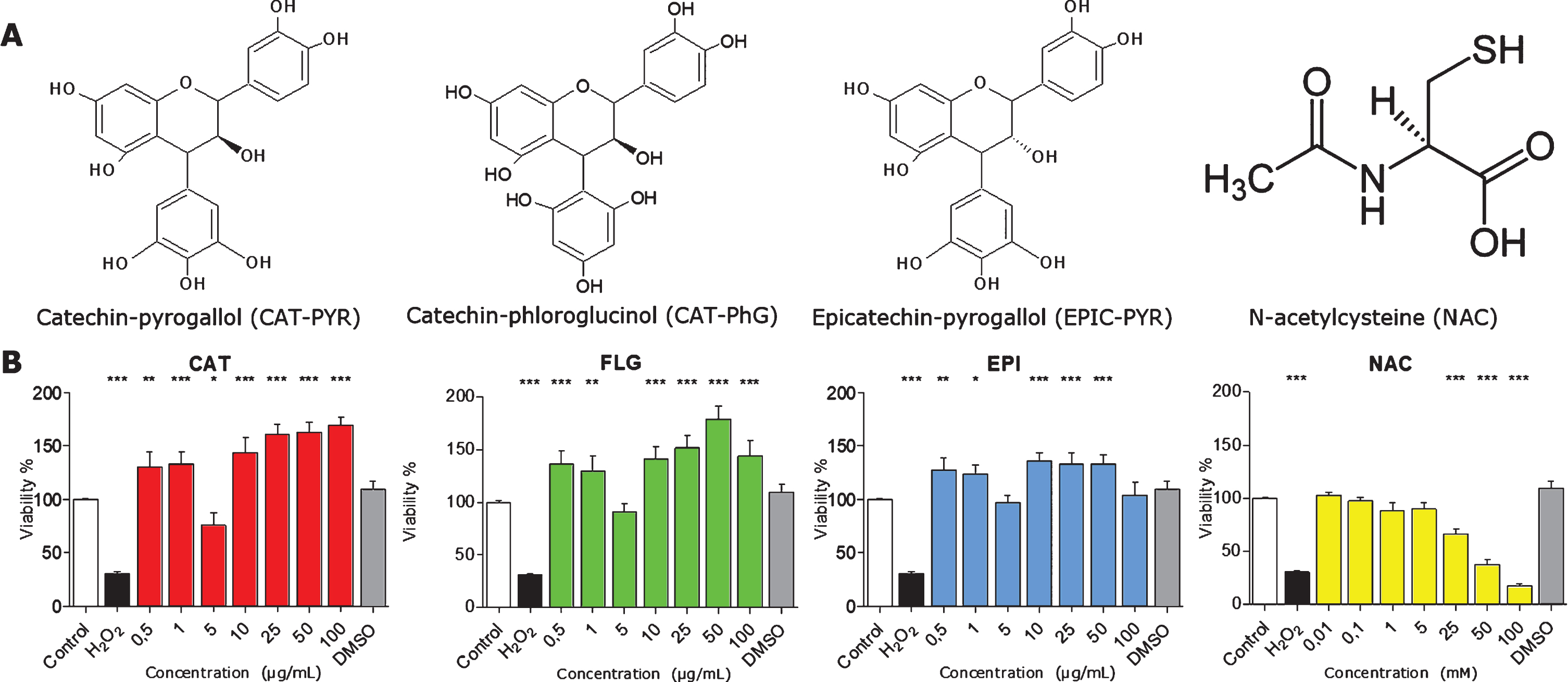
Viability of N2a cells in the presence of the different polyphenols. A) Structures for each bioactive molecule assessed CAT, FLG, EPI, and NAC. B) Viability assays by MTT. Concentrations from 0.5μg/mL to 100.0μg/mL of each polyphenol, or 0.01 mM to 100.0 mM for NAC, were tested in N2a cells. Only high concentrations (25.0, 50.0, and 100.0 mM) of NAC were slightly toxic to cells. N2A are viable and not morphologically affected in a wide range of concentrations of the bioactive compounds. Data represent mean ± s.e.m. *P < 0.05, **P < 0.01.
In vitro assays showed that the bioactive compounds control human tau self-aggregation
All bioactive compounds were tested for their potential anti-aggregative effect on human tau protein. All of them showed that were able to differentially inhibit tau self-aggregation (Fig. 3).

Anti-aggregation assay on the effects of polyphenols on in vitro recombinant tau self-aggregation. For 10 days, purified hTAU40 samples, with or without heparin, were subjected to self-aggregation in the presence of either of the bioactive polyphenolic compounds. 5.0μg/mL of the polyphenol molecules and 5.0 mM for NAC were used. Tracking curve of hTAU40 protein aggregates, after 10 days of registration. Each point corresponds to the mean of three different experiments. Standard errors are indicated for some determinations, errors in determinations were between 1–3% of the fluorescence measurement of each point. It is observed that 5.0μg/mL of each compound was able to prevent the formation of these tau aggregates.
DISCUSSION
NAC has the capacity to reduce the aggregation of htau to 60% of control, while EPI reduces to 30% human tau aggregation. As it has been mentioned before, nitroxidative stress promotes neurodegeneration and cellular death, mainly by the effect of tau phosphorylation [43]. Although research focused on tau protein and NAC is limited, some publications support this effect, as will be discussed next. Treatment of 300 mg/kg of NAC combined with other antioxidants, namely 2,4-disulfonyl α-phenyl tertiary butyl nitrone (HPN-07), reduce the accumulation of hyperphosphorylated tau protein after blast-induced injury in rats [44]. In human neuroblastoma cells, 30 mM NAC was capable of reversing the oxidative stress and toxicity induced by diverse damage agents, such as UV light or Aβ1–42, and decreased markedly tau phosphorylation [45]. All these studies are consistent with the effects observed in the present report, and that it is noteworthy that the action of NAC in inhibiting the process of tau self-aggregation, which is of major significance in the context of prevention and possible therapeutic intervention in neurodegenerative disorders such as AD and tauopathies.
In addition, diverse data relate NAC with different participants of the neuroimmunomodulation pathway, as described below. NAC promoted a decrease in cell death and the levels of cleaved caspase 3 after an HIV-1 (human immunodeficiency virus type1) Tat and gp120 treatment in N2a cells. Data correlated with studies on a transgenic rat model of HIV-1 infection, that exhibited hyperphosphorylated tau, increased CDK5 and GSK-3β expression levels, and proinflammatory cytokines as TNF-α, IL-6, and MCP-1, suggesting a role of NAC in the prevention of neurodegenerative process that affect to HIV-1 patients [43, 46]. In a similar way, NAC prevent social isolation-induced accelerated impairment of contextual fear memory, through the normalization of p35/Cdk5 activity, associated with the onset of early AD, in a transgenic mouse model [47]. In addition, NAC is capable to suppress NF-kappa-B activation triggered by TNFα in a cellular line [48]. After exposure to neurotoxic compound, NAC could prevent cell death and astrogliosis in rats’ cerebral cortex [49]. Altogether, these data supports the potential application of NAC and catechin derivatives in pharmacological intervention of brain disorders and AD, thus focusing on tau’s point of view, according to the molecular pathways of neuroimmunomodulation [10], which besides is sustained by the connection between oxidative stress and inflammation [50].
Conclusion
Our results indicate that certain semisynthetic polyphenols investigated in this study, namely CAT, FLG, EPI, and the gluthatione percursor NAC, are able to modulate a possible functional effect on a brain-derived cell line, promoting neuritogenic effects. This was evidenced by increase in length and number of neuronal processes. As well, they are able to promote an anti-aggregation effect preventing anomalous formation of tau oligomers. These studies, along with the current clinical data on natural epicatechin-derivatives and dementia, suggest the potential role of these compounds in controlling the neurodegenerative events in AD. These studies are of a major interest since other anti-oxidative compounds, such as the Andean Compound rich in fulvic acid [8, 22] and extracts from the rosemary plant [32], efficiently control tau oligomerization. New experiments and approximations are required in order to understand the exact molecular and cellular mechanisms of these polyphenols. These data support the need for promoting the development of further investigation of natural compounds as preventive agents of the onset and/or progression of neurodegenerative disorders.
Footnotes
ACKNOWLEDGMENTS
This study was supported by Innova, High Technology Project CORFO 14IEAT-28658 grant to R.M.B., the International Center for Biomedicine (ICC) and the Ricardo B. Maccioni Foundation for the financial support to carry out this research. We acknowledge Constanza A. Maccioni and Romina Rodriguez for their assistance and data organization in this research. E.P was funded by FONDEQUIP EQM150025.