
Abstract
We aimed to evaluate the association of PLCG2 rs72824905 variant with Alzheimer’s disease (AD) and multiple sclerosis (MS) using large-scale genetic association study datasets. We selected 50,024 AD cases and 467,330 controls, and 32,367 MS cases and 36,012 controls. We found moderate heterogeneity of rs72824905 in different studies. We found significant association between rs72824905 G allele and reduced AD risk (OR = 0.66, 95% CI 0.59–0.74, p = 5.91E-14). Importantly, rs72824905 G allele could also significantly reduce the risk of MS with OR = 0.94, p = 3.63E-05. Hence, the effects of rs72824905 on AD and MS are consistent.
INTRODUCTION
PLCG2 encodes an enzyme called phospholipase C gamma 2 (PLCγ2) [1, 2]. PLCG2 is expressed in myeloid lineage cells, microglia, and granule cells of the dentate gyrus [1, 2]. It has been reported that PLCγ2 could regulate osteoclastogenesis via its interaction with ITAM proteins and GAB2 [3]. Until now, multiple gain-of-function mutations have been identified to be associated with auto-inflammation and PLCγ2-associated antibody deficiency and immune dysregulation (APLAID) syndrome and pediatric common variable immunodeficiency (CVID) [4, 5]. The potential mechanism is that gain-of-function mutation in the murine Plcg2 gene could cause hyperreactive external calcium entry in B cells and expansion of innate inflammatory cells, and finally lead to severe spontaneous inflammation and autoimmunity [6].
Alzheimer’s disease (AD) is a complex neuro-degenerative disease, and large-scale genetic association studies have been widely conducted to identify common and rare genetic variants [7–10]. In 2017, a protective and rare coding variant rs72824905 in PLCG2 was first reported to be significantly associated with AD risk with p = 5.38E-10 and odds ratio (OR)=0.68 for G allele [11]. In 2019, van der Lee and colleagues conducted a candidate gene study using participants from European descent [12]. They aimed to evaluate the effect of the rs72824905 variant G allele on multiple neurodegenerative diseases including AD, Parkinson’s disease (PD), dementia with Lewy bodies, frontotemporal dementia, progressive supranuclear palsy, amyotrophic lateral sclerosis (ALS), and multiple sclerosis (MS), as well as longevity [12]. Interestingly, van der Lee and colleagues found that rs72824905 variant G allele could significantly reduce the risk of AD (p = 6.30E-04), dementia with Lewy bodies (p = 0.045), and frontotemporal dementia (p = 0.011), but identified no significant association of rs72824905 variant G allele with the risk of PD, ALS, MS, or progressive supranuclear palsy [12].
There are still two main concerns, although these are important and interesting findings. First, independent genetic association studies have reported inconsistent findings about the association between rs72824905 variant and AD risk in European population even with the almost equivalent AD sample size. In power analysis, van der Lee et al. concluded that the AD dataset (4,985 cases and 9,238 controls) had adequate statistical power (>0.8) to detect a protective association (p = 0.05) with an OR around 0.68, which was reported in the original study [11]. In 2019, Moreno-Grau et al. conducted a genome-wide association study (GWAS) using 7,409 unrelated individuals including 4,120 AD cases and 3,289 control individuals [13]. However, Moreno-Grau et al. reported no significant association between rs72824905 variant and AD risk with p = 0.3496 [13]. Until now, it remains unclear what causes the inconsistent findings.
Second, genetic association studies with large sample sizes are still required to evaluate the association between rs72824905 variant and MS risk. In power analysis, van der Lee et al. concluded that the available MS dataset (4,476 cases and 5,714 controls) had adequate statistical power (>0.8) to detect the protective association (p = 0.05) with an OR around 0.68 [12]. In discussion, van der Lee and colleagues ensured that the available sample sizes provided adequate statistical power and larger meta-analysis will unlikely observe an association of rs7282490 variant with MS, PD, and ALS [12]. In fact, the available MS sample size (4,476 cases and 5,714 controls) is relatively smaller than PD (27,595 cases and 106,951 controls) and ALS (10,953 cases and 20,673 controls) [12]. Importantly, large-scale MS GWAS datasets are available, which provides strong support to further investigate this association [14, 15].
Regarding the first concern, we consider that the inconsistent findings may be caused by the small sample size from the candidate study, or the significant genetic heterogeneity of rs72824905 variant in different populations or studies, as described in our previous studies [16, 17]. Here, we performed an updated analysis of 517,354 subjects including 50,024 AD cases and 467,330 controls from recent GWAS [11, 18], and candidate gene study datasets [12, 20]. Regarding the second concern, we consider that large sample size may contribute to identify significant association between rs72824905 variant and MS risk. Here, we performed a candidate study using the large-scale MS GWAS meta-analysis dataset [15].
MATERIALS AND METHODS
AD genetic datasets
The first dataset is from the International Geno-mics of Alzheimer’s Project (IGAP) [11]. The IGAP included 85,133 subjects (37,022 AD cases and 48,402 controls) from the Genetic and Environmental Risk for Alzheimer’s Disease (GERAD)/Defining Genetic, Polygenic and Environmental Risk for Alzheimer’s Disease (PERADES) Consortium, the Alzheimer’s Disease Genetic Consortium (ADGC), the Cohorts for Heart and Aging Research in Genomic Epidemiology (CHARGE), and the European Alzheimer’s disease Initiative (EADI) [11]. The second dataset is from the Genome Research at Fundacio ACE (GR@ACE) Consortium including 7,409 unrelated individuals (4,120 AD cases and 3,289 control individuals) [13]. The AD cases are from the Fundació ACE, Institut Català de Neurociències Aplicades (Catalonia, Spain). The controls are from Fundació ACE (Barcelona, Spain), Valme University Hospital (Seville, Spain), and the Spanish National DNA Bank Carlos III (University of Salamanca, Spain) [13]. The third dataset is from the UK Biobank including 404 AD cases and 402,383 controls (http://pheweb.sph.umich.edu/SAIGE-UKB/pheno/290.11) [18]. Meanwhile, we also selected three candidate gene study datasets [12, 20]. Of these candidate gene studies, Conway et al. conducted a meta-analysis in Caucasian and African American populations, and the population-specific results are not publicly available. Hence, this meta-analysis was defined a single candidate gene study. Finally, we selected a total of 517,354 subjects (50,024 AD cases and 467,330 controls), as provided in Table 1.
Sample information in AD GWAS and AD genetic association studies
Heterogeneity test
We first extract the corresponding summary statistics of rs72824905 variant in these above datasets. We then determine the heterogeneity of rs72824905 variant among these datasets using Cochran’s Q test and I2 = (Q -- (k --1)) / - Q × 100 %. The Q statistic approximately follows a χ2 distribution with k-1 degrees of freedom (k is the number of studies for analysis, here, k = 6). I2 ranges from 0 to 100%. I2 = 0–25%, 25–50%, 50–75%, and 75–100% indicates low, moderate, large, and extreme heterogeneity, respectively [16, 17].
Meta-analysis
A meta-analysis was used to evaluate the association between rs72824905 variant and AD using R Package (meta: General Package for Meta-Analysis) [17]. The overall OR is calculated by the fixed effect model (Mantel-Haenszel) or random-effect model (DerSimonian-Laird) [17]. The significance of OR was determined using the Z test. The statistical significance is 0.05, as described in recent studies [16, 17].
MS genetic dataset
The MS genetic dataset is from a large-scale meta-analysis of MS GWAS datasets from the International Multiple Sclerosis Genetics Consortium (IMSGC) [15]. This dataset consisted of 32,367 MS cases and 36,012 controls drawn across the IMSGC centers in Australia, 10 European countries, and multiple US states [15], as provided in Supplementary Table 1. A total of 120,991 low-frequency coding variants were genotyped by the Illumina Human Exome Beadchip or on a custom array [15]. The statistical significance is 0.05.
RESULTS
Heterogeneity test
Using all these six selected studies, the heterogeneity test showed no significant genetic heterogeneity of rs72824905 variant with Cochran’s Q test p = 0.5121 and I2 = 0 with 95% CI [0.0%; 70.2%]. We further conducted a sensitivity analysis by excluding IGAP study and found evidence of low genetic heterogeneity with Cochran’s Q test p = 0.3721 and I2 = 6.1%. Hence, we test the genetic heterogeneity between IGAP study and other five studies. The results showed that moderate heterogeneity between IGAP and GR@ACE with I2 = 48.9%. However, there is no evidence of heterogeneity between IGAP study and other four studies, as provided in Table 2. By excluding the GR@ACE, we found that the I2 and 95% CI changed from 0.0% [0.0%; 70.2%] to 0.0% [0.0%; 50.4%].
Heterogeneity between IGAP study and other five studies
Association between rs72824905 and AD
The meta-analysis showed significant association between rs72824905 variant G allele and AD risk using the fixed effect model (OR = 0.68, 95% CI 0.61–0.75, p = 1.58E-13) and the random-effect model (OR = 0.68, 95% CI 0.61–0.75, p = 1.58E-13), as provide in Fig. 1. Interestingly, meta-analysis by excluding GR@ACE further even showed more significant association between rs72824905 variant G allele and reduced AD risk using the fixed effect model (OR = 0.66, 95% CI 0.59–0.74, p = 5.91E-14) and the random-effect model (OR = 0.66, 95% CI 0.59–0.74, p = 5.91E-14).
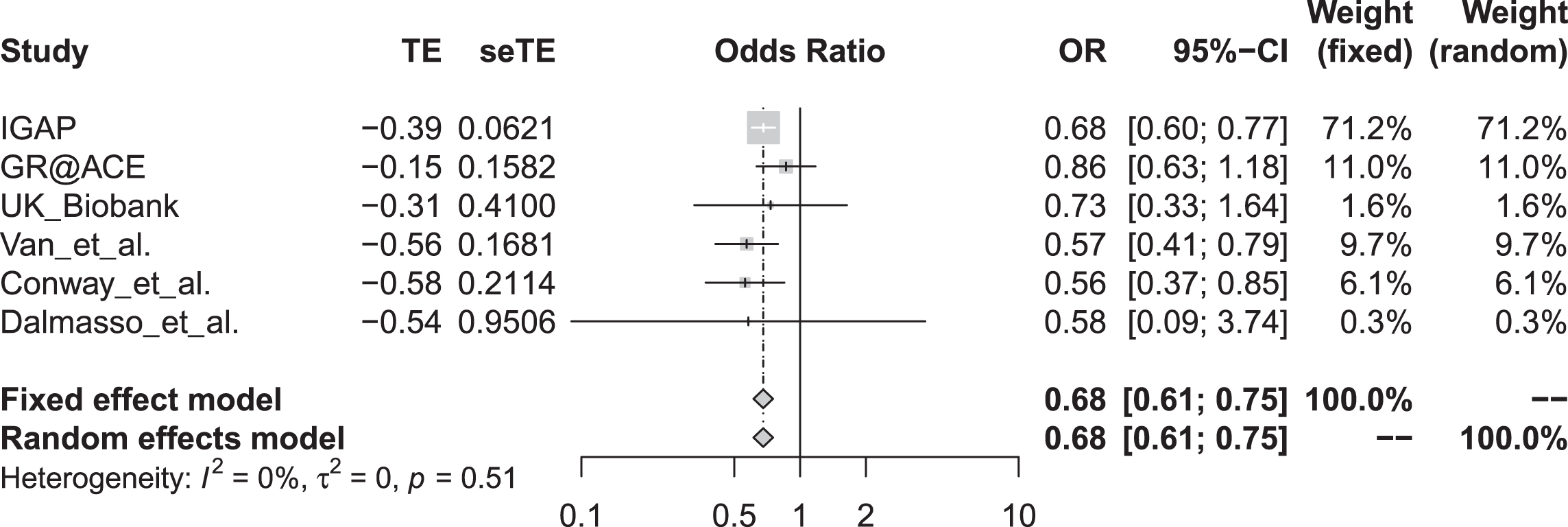
Forest plot for the meta-analysis of rs72824905 variant with AD risk. M-H, Mantel-Haenszel; Fixed, fixed effect model; CI, confidence interval.
Association between rs72824905 and MS
Interestingly, the results showed that rs72824905 variant G allele could significantly reduce the risk of MS with OR = 0.94, p = 3.63E-05. Hence, the effects of rs72824905 variant G allele on AD and MS are consistent with each other.
DISCUSSION
The protective and rare coding variant rs72824905 in PLCG2 was originally identified to be significantly associated with AD risk [11]. However, replication studies have reported inconsistent findings about the association between rs72824905 and AD [12, 18–20]. Here, we consider that the inconsistent findings may be caused by the small sample size from the following GWAS and candidate gene studies, or the genetic heterogeneity of rs72824905 variant in different populations [16, 17].
In order to confirm this view, we conducted this updated analysis of rs72824905 variant using 517,354 subjects including 50,024 AD cases and 467,330 controls from recent GWAS [11, 18], and candidate gene study datasets [12, 20]. We first evaluated the heterogeneity of rs72824905 variant in all these six selected studies and found no evidence of significant genetic heterogeneity. However, sensitivity analysis suggested that there was moderate heterogeneity between IGAP and GR@ACE with I2 = 48.9%, but no evidence of heterogeneity between IGAP study and other four studies. Importantly, meta-analysis by excluding GR@ACE even showed more significant association between rs72824905 variant G allele and reduced AD risk than using all these six selected studies. This may explain why van der Lee et al. (4,985 cases and 9,238 controls) and Moreno-Grau et al. (4,120 AD cases and 3,289 control individuals) used the almost equivalent AD sample size but got different findings [11, 13].
Meanwhile, multiple independent genetic association studies have been conducted to evaluate the association between rs72824905 and other neurodegenerative diseases such as MS [12]. In a recent study, van der Lee et al. found no association between rs72824905 and MS, and concluded that the available MS dataset (4,476 cases and 5,714 controls) had adequate statistical power and larger meta-analysis will unlikely observe an association of rs7282490 variant with MS [12]. Here, we conducted a candidate variant study using 32,367 MS cases and 36,012 controls, and revealed rs72824905 variant G allele to be significantly associated with the reduced risk of MS.
Interestingly, this finding is consistent with recent findings that rs72824905 variant and PLCG2 promotes immune functions [2, 21]. A knock-in (KI) mouse model using CRISPR/Cas9 gene editing reveals that rs72824905 could potentiate the primary function of Plcγ2, improve survival and increase acute inflammatory response of the KI macrophages, enhance microglial functions [2]. A genetic association study indicated that rs72824905 G allele associates with lower Braak stage and reduced quantitative tau pathology [22]. In COS7 and HEK293T cells, Magno et al. found that rs72824905 could weakly increases enzyme function [1]. Meanwhile, rs72824905 could mitigate tau pathology in the presence of amyloid pathology, reduce AD progression and maintains cognitive function [23, 24]. PLCG2 connected APOE and TREM2 in co-expression protein network [24], and PLCγ2 activity could regulate divergent microglial functions dependent and independent of TREM2 signaling [21].
In summary, we conducted an updated analysis of the rs72824905 variant with the risk of AD and MS. Using 50,024 AD cases and 467,330 controls, we demonstrated that the genetic heterogeneity may cause different findings about the roles of rs72824905 variant in different studies. Large sample size may contribute to identify significant findings without significant genetic heterogeneity. Using 32,367 MS cases and 36,012 controls, we conducted a candidate variant study and revealed rs72824905 variant G allele to be significantly associated with the reduced risk of MS. Hence, we not only confirmed the association between rs72824905 and AD, as reported by van der Lee and colleagues, but also highlighted the protective role of rs72824905 in MS, which was not identified by van der Lee and colleagues.
Footnotes
ACKNOWLEDGMENTS
We thank the International Genomics of Alzhei-mer’s Project (IGAP). The investigators within IGAP contributed to the design and implementation of IGAP and/or provided data but did not participate in analysis or writing of this report. IGAP was made possible by the generous participation of the control subjects, the patients, and their families. The i-Select chips was funded by the French National Foundation on AD and related disorders. EADI was supported by the LABEX (laboratory of excellence program investment for the future) DISTALZ grant, Inserm, Institut Pasteur de Lille, Université de Lille 2 and the Lille University Hospital. GERAD was supported by the Medical Research Council (Grant n° 503480), Alzheimer’s Research UK (Grant n° 503176), the Wellcome Trust (Grant n° 082604/2/07/Z) and German Federal Ministry of Education and Research (BMBF): Competence Network Dementia (CND) grant n° 01GI0102, 01GI0711, 01GI0420. CHARGE was partly supported by the NIH/NIA grant R01 AG033193 and the NIA AG081220 and AGES contract N01-AG-12100, the NHLBI grant R01 HL105756, the Icelandic Heart Association, and the Erasmus Medical Center and Erasmus University. ADGC was supported by the NIH/NIA grants: U01 AG032984, U24 AG021886, U01 AG016976, and the Alzheimer’s Association grant ADGC-10-196728. We thank the International Multiple Sclerosis Genetics Consortium (IMSGC) for providing the large-scale meta-analysis dataset about 120,991 low-frequency coding variants from 32,367 MS cases and 36,012 controls drawn across the IMSGC centers. We also thank the Genome Research at Fundacio ACE (GR@ACE) Consortium, UK Biobank, and Conway et al. and Van der Lee et al. for the genetic association study datasets. This work was supported by funding from the National Natural Science Foundation of China (Grant No. 82071212, 6180010993, and 81901181), and the Mathematical Tianyuan Fund of the National Natural Science Foundation of China (Grant No. 12026414).