
Abstract
Alzheimer’s disease (AD) is becoming a prevalent disease in the elderly population. Past decades have witnessed the development of drug therapies with varying targets. However, all drugs with a single molecular target fail to reverse or ameliorate AD progression, which ultimately results in cortical and subcortical network dysregulation. Deep brain stimulation (DBS) has been proven effective for the treatment of Parkinson’s disease, essential tremor, and other neurological diseases. As such, DBS has also been gradually acknowledged as a potential therapy for AD. The current review focuses on DBS of the nucleus basalis of Meynert (NBM). As a critical component of the cerebral cholinergic system and the Papez circuit in the basal ganglia, the NBM plays an indispensable role in the subcortical regulation of memory, attention, and arousal state, which makes the NBM a promising target for modulation of neural network dysfunction and AD treatment. We summarized the intricate projection relations and functionality of the NBM, current approaches for stereotactic localization and evaluation of the NBM, and the therapeutic effects of NBM-DBS both in patients and animal models. Furthermore, the current shortcomings of NBM-DBS, such as variations in cortical blood flow, increased temperature in the target area, and stimulation-related neural damage, were presented.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a neurodegenerative disease with increased prevalence in the older adult population and is the leading cause of dementia in elderly individuals. According to a WHO survey in 2019, approximately 50 million people globally suffered from AD, which presents a notable healthcare burden and poses a formidable threat to human longevity [1]. Clinically, AD manifests as progressive cognitive decline, behavioral disorders, and psychiatric syndromes, which are primarily related with its histopathological hallmarks: senile plaques and neurofibrillary tangles. The pathology of AD is mainly induced by tau hyperphosphorylation and oligomerization inside the neurons, as well as the deposition of neuritic Aβ plaques in the brain [2]. Moreover, the loss of cholinergic neurons and synapses is also believed to be responsible for cognitive decline [3].
Treatment of AD mainly relies on drugs, including cholinesterase inhibitors, which are recommended as the first-line drug by various guidelines [4–7], and glutamine NMDA receptor antagonist, which has been approved for application in moderate-to-severe AD [8, 9]. However, no drugs are currently available to reverse or even alleviate AD’s underlying neurodegeneration, although a group of drugs has been tested that prevents the deposition of Aβ or tau hyperphosphorylation, with negative results in phase III clinical trials [10–13]. The failure of single-target drugs promotes research into non-pharmacological strategies seeking to modulate the dysfunctional memory circuit across multiple neural networks in AD.
Many neuroradiology and electrophysiology studies have indicated that abnormalities in memory circuits may be involved in AD pathogenesis. Functional MRI (fMRI) research revealed hippocampal hyperactivity and reduced deactivation of the default-mode network (DMN) in patients presented mild cognitive impairment (MCI) at the prodromal stage of AD [14]. The dysfunction in DMN components can be seen in sporadic and familial AD patients decades before the diagnosis [15]. In PSEN1 mutation carriers, the preceding time can even be as far as 30 years [16]. Abnormal neuronal synchronized activities were also commonly seen in AD patients’ quantitative electroencephalography (qEEG), featured by a reduced neocortical alpha and gamma power [17, 18]. Moreover, the DMN where blood-oxygen-level-dependent fMRI showed decreased signals was found to coincide with the area appeared declined gamma power on qEEG in AD patients [19]. Accumulation of Aβ in the entorhinal cortex and the hippocampus has been observed in AD patients, which was closely related with the dysfunction of the memory-related Papez circuit [20]. Although AD is a heterogeneous disease with intricate mechanisms, those evidence has pointed to the circuitopathy nature of AD.
Therefore, physical stimulation therapy strategies to modify brain circuits and networks are currently hot areas, including transcranial direct current stimulation, repetitive transcranial magnetic stimulation, and deep brain stimulation (DBS). Although transcranial direct current stimulation and repetitive tra-nscranial magnetic stimulation have emerged as prospective non-invasive methods, both have a broad stimulation range and lack specific targets, and thus, they cannot be used to modulate specific neural circuits. DBS was demonstrated to be effective in engaging and modulating the specific neural circuits and networks involved in cognition, and emerged as a powerful strategy for AD treatment [20, 21].
The DBS device is composed of an electrode, whi-ch is stereotactically implanted to a certain nucleus or fiber tract, as well as a pulse generator embedded underneath the skin. The research conducted by Turnbull et al. in 1984 is believed to be the first clinical trial for DBS in AD treatment; the study found that DBS had no definite neurological effect, but preser-ved increased cortical glucose metabolic level in the nine-month follow-up [22]. Although DBS has not been formally listed as a technique used for AD tre-atment, it has received approval for treating movement disorders such as Parkinson’s disease (PD), essential tremor, and dystonia [23]. Accumulating clinical research also agreed with its robust therape-utic effect in diseases such as intractable epilepsy, obsessive-compulsive disorder, and depression [24–26]. For AD, a phase II trial of fornix DBS with enrollment of 42 mild AD patients has shown a sustained response in elderly patients [27] (Table 1). DBS can improve the glucose metabolism of the cortex and ameliorate the impairment of cognition in mild to moderate AD patients [28–30]. Mouse models of various AD transgenic strains have been elaborated and they provide primary evidence for different stimulation targets. Proper selection and accurate loc-alization of the target can increase the success of DBS. Clinical and animal studies of DBS for AD have targeted on the nucleus basalis of Meynert (NBM), as well as the fornix, entorhinal cortex, and hippocampus [22, 30–38] (details in Tables 1 and 2).
Summary of NBM deep brain stimulation clinical trials
AD, Alzheimer’s disease; ADAS-cog, Alzheimer’s disease Assessment Scale-Cognitive section; DBS, deep brain stimulation; GPi, globus pallidus interna; NA, not available; MDS-UPDRS, Movement Disorder Society-Unified Parkinson Disease Rating Scale; MMSE, Mini-Mental State Examination; NBM, Nucleus basalis of Meynert; PDD, Parkinson’s disease dementia; STN, subthalamic nucleus. Positions in trials with multiple target sites are presented as mean value (range). aNBM coordinates are presented aligning with the Talairach coordinates described in the atlas of Mai et al. [52]. x is the distance lateral to the third ventricle wall, y is the distance posterior to the anterior edge of the posterior commissure, and z is the distance ventral to the anterior commissure-posterior commissure line. bNBM coordinates are presented aligning with the midcommissural point coordinates. x, y, z are the distances lateral, posterior and ventral to the midcommissural point respectively. cThe research information is presented as “therapy duration, follow-up period”. When the therapy duration exceeded the follow-up period, only the latter time is shown here. *This patient received bilateral STN and NBM stimulation with 4 electrodes. **This patient received bilateral GPi and NBM stimulation with 2 electrodes. GPi stimulation started right after the operation, while NBM stimulation started 2 months later.
Summary of studies of NBM deep brain stimulation in AD animal models
AD, Alzheimer’s disease; MBM, modified Barnes maze; MWM, Morris water maze; OFT, open field test. APP/PS1 mouse refers to the amyloid-β precursor protein/presenilin1 (HuAPP695swe, PSEN1-dE9) transgenic mouse. TgF344 -AD rat refers to the mutant human amyloid precursor protein (APPsw) and presenilin1 (PS1ΔE9) transgenic rat. NBM coordinates are presented aligning with the mouse or rat coordinates. AP, ML and DV are the distances anterior, lateral and inferior to the bregma respectively.
The NBM is a key area in the cholinergic system and consists of the neurons that provide the major source of acetylcholine projection to the neocortex, which supports normal cognitive function. Tau pathology in the NBM was observed in AD during disease progression [39]. Therefore, the NBM stands out as a promising target. Thus far, two phase I clinical trials on NBM-DBS in AD treatment have been completed, revealing ameliorated Alzheimer’s Disease Assessment Scale-Cognitive Subscale (ADAS-Cog) decline and improved cortical metabolism [30, 32]. In animal studies, NBM-DBS showed a neuroprotect-ive effect on APP/PS1 mice, with reduction of Aβ deposition and neuronal apoptosis as well as better performance in spatial memory tasks [37]. In the present review, we summarized the current research about NBM-DBS in dementia treatment in order to emphasize the NBM’s cognitive network connection, assessment methods to determine the NBM’s location, effects and tolerance of NBM-DBS in clinical practice, and its main shortcomings and future development prospects.
THE NBM, ITS ANATOMY AND FUNCTIONAL PROJECTION, ELECTROPHYSIOLOGICAL PROPERTY, AND ROLE IN COGNITIVE DYSFUNCTION
The NBM (Ch4) is located in the posterior domain of the basal forebrain. In humans, the NBM stretches horizontally from the olfactory tubercle to the uncal gyrus of the hippocampus anteroposteriorly along the midline. The anterior part of the NBM is circumscribed by the horizontal limb of the nucleus of the diagonal band of Broca’s area inferiorly, the ventral globus pallidus superomedially, and the anterior commissure superolaterally. The posterior part of the NBM adjoins the ansa lenticularis superiorly, with the amygdala on the inferior side, the putamen on the lateral side, and the optic tract on the medial side [40, 41] (Fig. 1). The main nucleus can be subdivided by the populations of cholinergic neurons it contains, consisting the anteromedial portion (Ch4am), the anterolateral portion (Ch4al), the intermediate portion (Ch4i), and the posterior portion (Ch4p) (Fig. 2). Notice that the bridging part between the anterior and intermediate NBM was recognized as an anterointermediate portion (Ch4ai) in some studies [40]. The cholinergic cell group which was named nucleus subputaminalis and was superolateral to the NBM main nucleus was also acknowledged as a part of NBM [42].
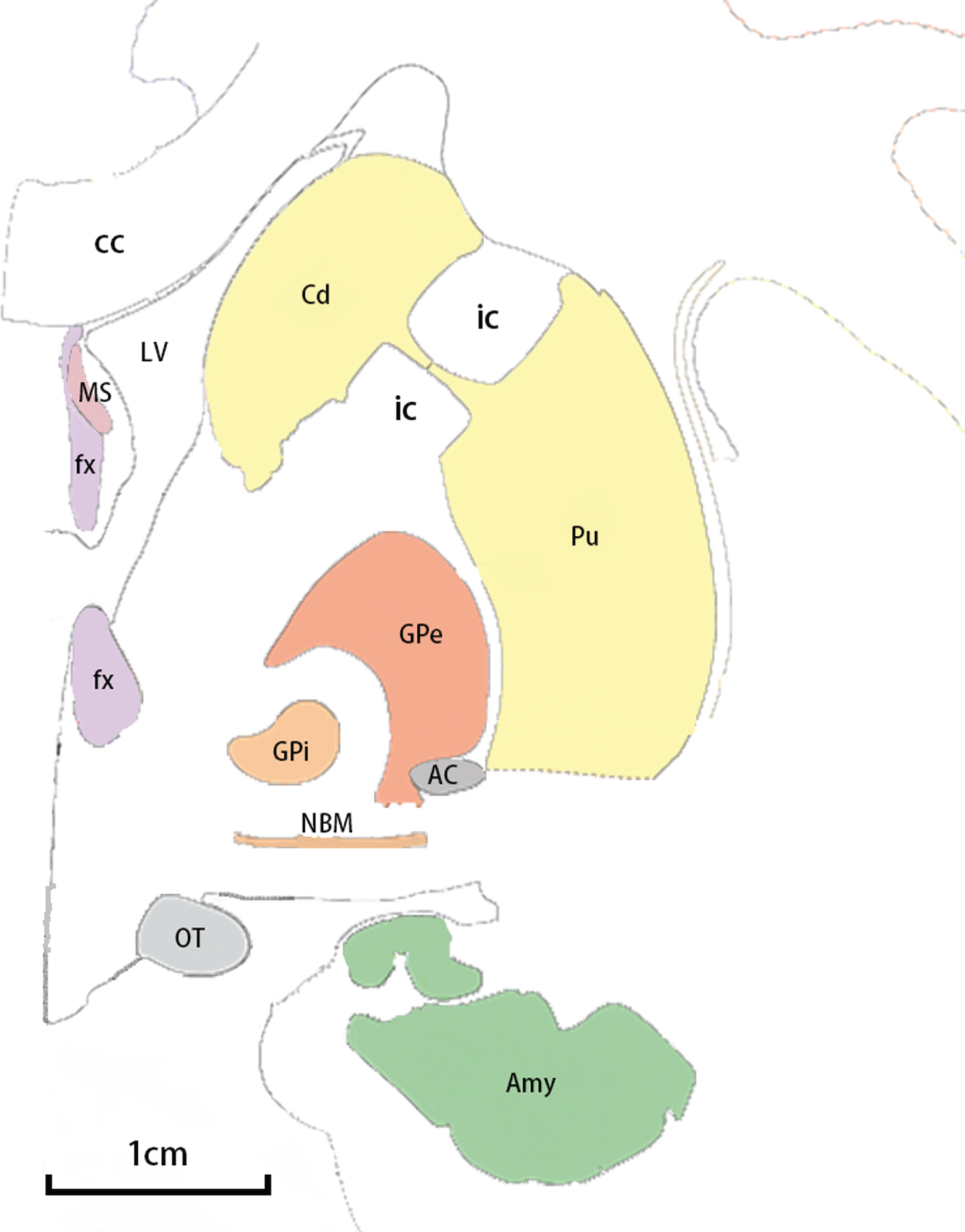
Adjacent structures of NBM in a coronal view. Major adjacent structures of NBM are shown in an anterior-posterior coronal view with an 1 cm scale. The illustration is based on Atlas of the Human Brain, third ed. [87]. AC, anterior commissure; Amy, amygdala; cc, corpus callosum; Cd, caudate nucleus; fx, fornix; GPe, globus pallidus externa; GPi, globus pallidus interna; ic, internal capsule; LV, lateral ventricle; MS, medial septum; NBM, nucleus basalis of Meynert; OT, optic tract; Pu, putamen.
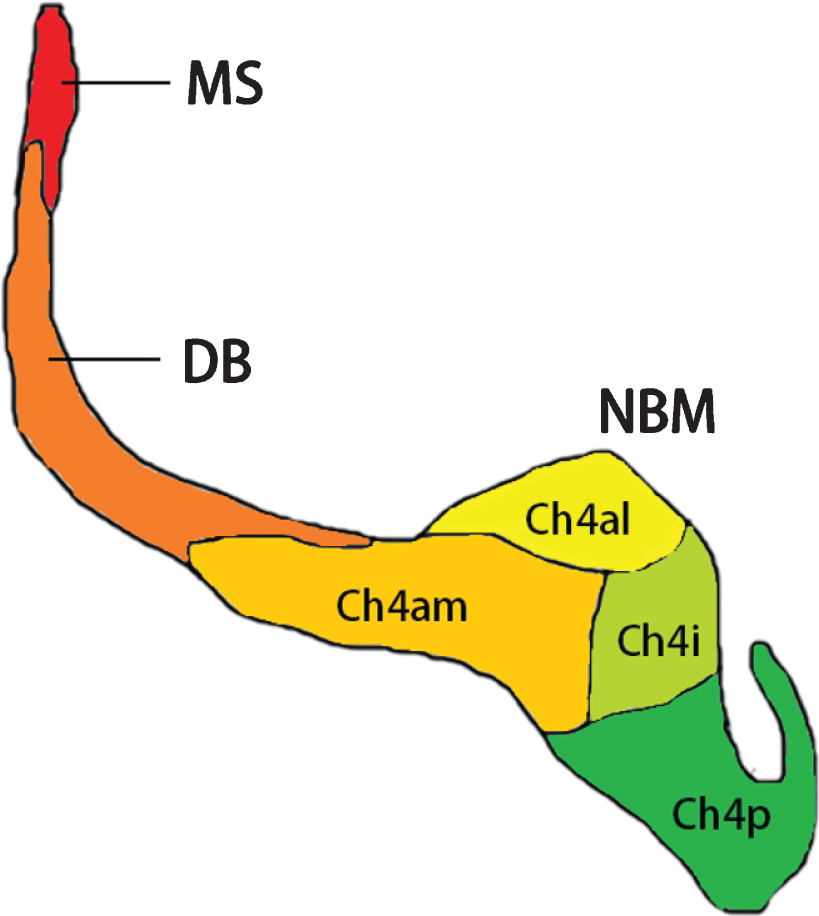
Diagram for subsectors of NBM. Subsectors of NBM are shown in a posterior-anterior view. Adjoined structures of the cholinergic system, including the medial septum (Ch1) and the diagonal band (Ch2 and Ch3) are also presented. The illustration is based on Mesulam and Geula [40]. Ch4al, anterolateral portion of NBM; Ch4am, anteromedial portion of NBM; Ch4i, intermediate portion of NBM; Ch4p, posterior portion of NBM; DB, diagonal band; MS, medial septum; NBM, nucleus basalis of Meynert.
Efferent fibers from the NBM constitute the largest cholinergic innervation to the neocortex. All of these efferent fibers converge to two major bundles: medial and lateral. The medial bundle leaves the anterior NBM and curves around the corpus callosum. After entering the cingulum, the bundle travels toward the posterior part and finally merges with the fibers from the lateral bundle in the occipital lobe [43]. The lateral bundle has two branches. The capsular branch travels in the external capsule and sends the fiber to the amygdala and the temporal cortex. The perisylvian branch first goes into the white matter around the claustrum and then radiates out to innervate the frontoparietal cortices, superior temporal gyrus, and the insula [44] (Fig. 3).
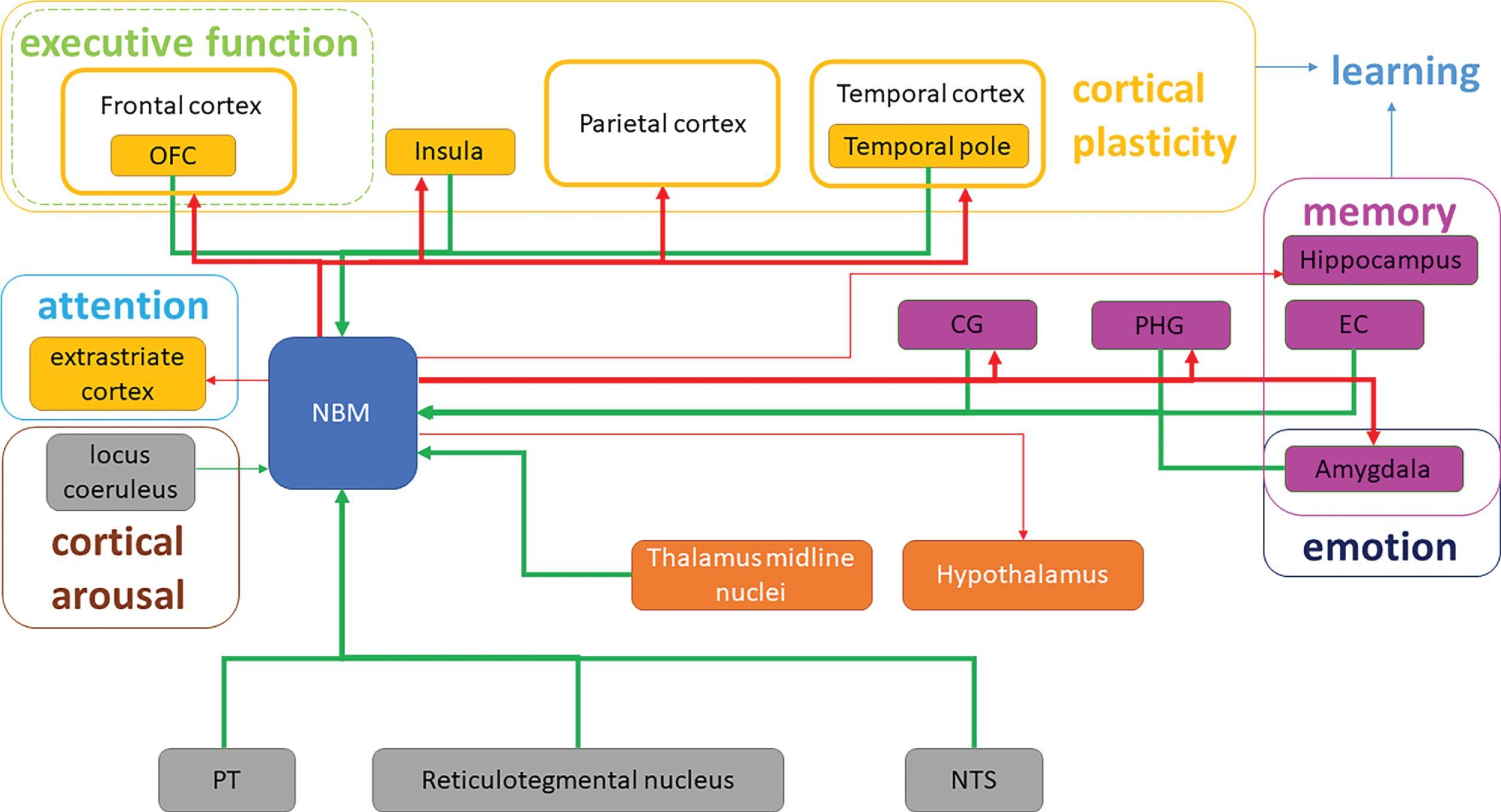
Schematic diagram of NBM fiber projection and cognitive domains modification in the whole brain. Efferent and afferent cholinergic fiber projections of NBM are shown. Red arrows represent the efferent fibers, while green arrows represent afferent fibers. Thick and fine arrows represent the major and minor projections, respectively. CG, cingulate gyrus; EC, entorhinal cortex; NST, nucleus of solitary tract; OFC, orbitofrontal cortex; PHG, parahippocampal gyrus; PT, pontomesencephalic tegmentum.
The NBM draws input from areas mostly limited to the limbic system, of which the central amygdala accounts for a large proportion [40]. As for cortical sources, the orbitofrontal, insular, temporopolar, parahippocampal, entorhinal, and cingulate regions contribute to the afferents [45]. Furthermore, certain neurons in the thalamus (midline thalamic nuclei) and the brainstem (pontomesencep-halic tegmentum, reticular formation, and the nucleus of the solitary tract) also project to the NBM [46, 47] (Table 3, Fig. 3).
Summary of NBM fiber projection to other anatomical structures
CG, cingulate gyrus; EC, entorhinal cortex; IPL, inferior parietal lobule; ITG, inferior temporal gyrus; NST, nucleus of solitary tract; OFC, orbitofrontal cortex; PHG, parahippocampal gyrus; PT, pontomesencephalic tegmentum; STG, superior temporal gyrus.
The basic electrophysiological properties of NBM cholinergic neurons have been confirmed by abundant pharmacological studies. The study using the human fetal NBM indicated that NBM cholinergic cells express voltage-gated Na+channels which are featured with rapid activation and inactivation and K + channels which experience fast activating and de-layed rectification. NBM cholinergic neurons can be excited by the acetylcholine via both nicotine and muscarinic receptors [48]. Apart from the acetylcho-line, NBM cholinergic neurons are also excited by the serotonin via the 5-HT1A receptors [49], by the noradrenalin via the α1 receptors with the participation of β receptors [50] and by the histamine via the H1 and H2 receptors concomitantly [51]. The primary inhibitory neurotransmitter acting on NBM cholinergic neurons is γ-aminobutyric acid (GABA). Animal study has revealed that the inhibitory effect of GABAergic inputs was mediated by GABAA rec-eptors specially and evoked depolarization yet inhibition of NBM cholinergic neurons [52]. Animal model with intraventricular administration of Aβ25–35 has been used in the research on the effect of high-frequency stimulation of NBM. In terms of spike activity, higher background spike train frequency was found in the hippocampus. The AD group showed relatively enhanced excitatory response (tetanic potentiation-post-tetanic potentiation, TP-PTP) of the hippocampus and relatively enhanced inhibitory response (tetanic depression-post-tetanic depression, TD-PTD) of the amygdala to high-frequency stimulation of NBM. These results indicated that normal cholinergic activity of NBM was responsible for the maintenance of the balance between excitation and inhibition of neurons in the hippocampus and the amygdala [53].
NBM is an important part of the cholinergic system, which provides the most widespread cholinergic innervation of the neocortex in the four cholinergic groups of the basal forebrain including the medial septum (MS, Ch1), the vertical limb of the dia-gonal band (vDB, Ch2), the horizontal limb of the diagonal band (hDB, Ch3), and NBM (Ch4). The fun-ctional connectivity of NBM revealed by resting-state fMRI has been indicated as a predictor for treatment response of cholinesterase inhibitors in MCI stage of AD [54]. The basal forebrain, especially NBM, presents strong connectivity with other portions of the cholinergic system. The basal forebrain receives cholinergic innervation from the olfactory system and is engaged in the olfactory memory retrieval of the rodent [55]. Particularly, cholinergic neurons of NBM are mainly innervated by the basolateral and the central amygdala, and send reciprocal projections back [56]. NBM also sends tremendous cholinergic projections to the limbic area. These projections take an active part in the maintenance of attention and cortical arousal which are further discussed below. A group of NBM neurons indirectly communicates with the hippocampal formation especially the entorhinal cortex and the hippocampus via the fornix and the cingulate gyrus. Loss of cholinergic transmission from the basal forebrain in AD patients can cause degeneration of entorhinal cortex [57]. Volume loss of the NBM detected by T1-weighted MRI is found to be associated with atrophy of hippocampus and amygdala in MCI patients, and 10 mg/day donepezil treatment reduced the progression of NBM volume loss on MRI in the 12 months of follow-up [58, 59]. Furthermore, early studies have shown that NBM was also related with cholinergic neuron groups in the brainstem such as the pedunculopontine nucleus (Ch5) and laterodorsal tegmental nucleus (Ch6) through common thalamus innervation, although these nuclei are relatively spared in AD progression [60]. For other neural circuits, although NBM is generally not regarded as a core member of the Papez’s Circuit and the major projections of NBM do not pass through the fornix directly, it is reported that the whole basal forebrain receives precommissural fibers from the hippocampal pyramidal cells via the fornix [41]. A diffusion tensor imaging (DTI) study by Ray et al. showed that the white matter in the fornix suffers atrophy in both AD and MCI patients, which may correlates with the dysfunction of the cholinergic system of basal forebrain including NBM [61]. This fiber tract may also contain opposite projections. Besides, NBM also receives input from the entorhinal cortex which contributes to memory consolidation [62, 63].
The NBM plays multiple regulatory roles in several cognitive domains. Typically, the NBM modulates the formation and configuration of memory [64, 65]. NBM cholinergic neurons that project to the basolateral amygdala are critical to the generation and the extinction of contextual fear memory [66]. Moreover, pharmacological study indicates that cholinergic projections from the NBM to the hippocampus participate in and modulate hippocampal function in spatial memory and learning [65]. Theta oscillation of the hippocampus takes place at active memory state, which is related with information transfer and encoding [67, 68]. During rapid eyes movement (REM) sleep, theta rhythm also appears in the hippocampus where memory replay and consolidation are believed to be undertaken [69]. Classic theory model established that the cholinergic and GABAergic systems in the basal forebrain were the pacemaker of the theta rhythm of the hippocampus through their projections to the interneurons and the pyramidal cells in CA1. In the model, the medial septum (MS) and the diagonal band were more focused as the generators [70]. However, accumulating evidence has shown that integrity of NBM also takes a critical role in the formation of hippocampal theta rhythm and working memory. Research on event-related oscillations suggested that the NBM cholinergic system may modify the synchronization/phase resetting of different frequencies within a certain brain area, which was different from the inter-area modulation of the MS system [71]. Damage to the NBM can induce suppressed theta oscillation in the hippocampus, which causes worse performance in learning and memory tasks. The suppressed theta oscillation here is restorable by Huperzine A, a selective inhibitor of acetylcholinesterase, suggesting that NBM may interact with the hippocampus through cholinergic innervations [72]. Apart from the hippocampal projections, fibers from the NBM to the cortex also engage in memory regulation. Evidence has shown that basal forebrain cholinergic neurons are involved in olfaction-related memory in rodents, as the entire olfactory region including the olfactory bulb and the piriform cortex has established reciprocal interaction with the NBM [55]. Additionally, cholinergic fibers from the basal forebrain, particularly the NBM, to the prefrontal cortex can also mediate Pavlovian and conditioned fear learning and extinction [73].
Research has also revealed the critical function that NBM is involved in modulating cortical plasticity, which is of vital importance in learning and memory formation and consolidation. Excitation of the NBM cholinergic fibers was found to facilitate the cortical plasticity. Electrical stimulation of NBM coupling with an auditory cue reorganized the representative map of the primary auditory cortex in rats [74]. Further study indicated that innervation from NBM assigned the significance of a stimulus and improved learning. This reorganization was soon normalized and was not necessary for good performance in a learning task [75]. According to this principle, Engineer et al. applied vagus nerve stimulation as a less invasive method with tone pairing to treat tinnitus model rats, remodeling the cortical map and normalizing the map distortion of tinnitus [76]. Similar to the auditory cortex, NBM cholinergic projections enhanced the response of the insular cortex in novel taste recognition as well as gustatory memory retrieval, which can be inhibited by GABAergic projections also from NBM [77]. Lesion of NBM by 192 IgG-saporin also induced dendritic atrophy in the frontal cortex, which was related with cholinergic innervation and was more severe in aged rats than young rats [78].
Another function of the NBM is the maintenance of attention. Cholinergic innervation from the NBM in the medial bundle boosts the activity of the extrastriate cortex and facilitates visual attention [79]. Cholinesterase inhibitors can promote the enhancement of visual attention led by sensory stimulus detection and encoding in the extrastriate cortex, which is regulated by the NBM [80]. NBM lesions caused by stereotaxic injection of 192 IgG-saporin (which selectively inhibits ribosomes in the cholinergic neurons to cease protein composition) in rats resulted in increased omissions of light stimulus presentation for the reward and decreased correct responses in the five-choice serial reaction time task (5-CSRT), which indicated impaired visuospatial attention [81].
The NBM is also responsible for cortical arousal. It receives noradrenergic projections from the locus coeruleus, which is believed to be critical to the ascending activation system of the cerebral cortex [31, 82]. In generally anesthetized mice, neocortical activity was dominated by 0.5–2 Hz synchronous deflections. Optogenetic selective stimulation of cholinergic neurons in the NBM, as well as their axons in the neocortex, has been proven to evoke gamma oscillation desynchronization in the neocortical local field potential (LFP), just as a tail pinch does, on anesthetized mice, suggesting that the NBM has a promoting effect on the awake state [83]. Earlier studies revealed that the electroencephalograph (EEG) of AD patients presented neocortical generalized slow waves, and that impeded cholinergic-monoaminergic interactions likely contribute to EEG slowing in dementia [84, 85]. EEG slowing suggests the inability of cholinergic innervation to maintain arousal, which may consequently inhibit attention, task learning, and memory formation in AD.
IMPLANTATION OPERATION AND TARGET EVALUATION IN NBM-DBS SURGERY
Implant parameters
For clinical practice, human brain atlases such as the Schaltenbrand and Wahren’s Atlas for Stereotaxy of the Human Brain and Mai JK.’s Atlas of the Human Brain are commonly used in the registration of DBS surgery with MRI [86, 87]. The coordinates applied in clinical trials, according to the Atlas of the Human Brain, located the NBM at x = 12.5 mm lateral to the wall of the third ventricle, y = 4 mm posterior to the anterior border of the posterior commissure, and z = 5 mm ventral to the anterior-posterior commissure line [30, 36].
Stimulating electrode accuracy evaluated by MRI imaging
The location and atrophy of the NBM in AD patients can be effectively recognized with 1.5-Tesla or 3.0-Tesla MRI [88, 89]. During the early stage of DBS surgery, intraoperative CT combined with postoperative MRI was widely used for target confirmation [90]. In recent years, intraoperative MRI has been welcomed as a feasible method, as it allows implantation under general anesthesia with safety and accuracy [91, 92], which is usually utilized to calibrate indicators in the brain during DBS registration. This technique can also offer surveillance on acute changes, such as hemorrhage and brain shift caused by respiratory movement, loss of cerebrospinal fluid, and other factors during the operation [93]. For centers equipped with MRI systems of 7.0-Tesla or higher field strength, direct visualization preoperatively or postoperatively can assist in target discrimination and evaluation [94]. In clinical practice, various sequences are applied, including regular T1-weighted, T2-weighted, and T2-FLAIR sequences, and especially specific sequences to facilitate the display of the target nucleus. Kerl et al. showed that the T2*-FLASH2D sequence on the 3.0-Tesla MRI provides optimal delineation of the subthalamic nucleus for PD-DBS [95]. These increasingly advanced MRI techniques are illuminating for NBM-DBS in AD treatment. Application of DTI may also help to avoid critical tracts during implantation of the DBS electrode. In STN-DBS, DTI may assist to avoid the pyramidal tract and approach the dentatorubrothalamic tract so as to reduce the incidence of tremor and dysarthria. The distance between the electrode contacts and the center of dentatorubrothalamic tract was negatively correlated with the improvement of tremor symptom evaluated by UPDRS (r = –0.459, p < 0.001). On the contrary, contacts inducing dysarthria was found to be 0.64 mm in average closer to the center of pyramidal tract [96]. DTI can detect fiber tracts in the white matter and outline the macroscopic axonal organization in the brain parenchyma, which provides a clear display for alterations in the projection network in AD patients, and also helps to determine the proper stimulator implantation route [97]. Teipel et al. found that atrophy of the NBM in AD patients can cause detectable damage on the integrity of fiber tracts, which mainly appear in the corpus callosum, cingulate, superior longitudinal fasciculus, and other cortical tracts; this can influence for target visualization and follow-up in NBM-DBS therapy [88]. As a sensitive technique for brain blood oxygen metabolism, fMRI can be used to play a role in presurgical evaluation for NBM neu-romodulation. The NBM has been successfully segregated from adjacent nuclei in the basal forebrain with resting-state fMRI, which can improve detection of metabolic dysfunction in the NBM in AD patients, and can also predict the effect of DBS surgery by examining functional connectivity [98].
Target evaluation by intraoperative microelectrode recording
Considering that an atlas alone cannot cover ind-ividual variations in anatomic structures, during the DBS surgery, real-time monitoring can be provided by microelectrode recording (MER) before implanting the stimulation electrode. While the microelectrode advances along the scheduled route, MER identifies featured electrical activity from the target and helps to avoid implanting the electrode in the wrong structures. MER detects the extracellular pot-ential from cell bodies, dendrites, and axons of the target with fine microelectrodes, and identifies specific discharge patterns. Parameters often used in MER include single-unit action potential, firing rates, and background activity level. Burst index is another important parameter for firing patterns, which is def-ined as the ratio of mean and modal interspike intervals [99]. A burst index between 1 and 1.5 is recognized as a regular firing interval, between 1.6 and 9.9 is an irregular firing interval, and over 10 indicates a bursting pattern. Lee et al. reported a specific recording pattern of MER in their study of NBM-DBS targeting for PD patients [100]. According to the case report, the trajectory of the microelectrode passed through the globus pallidus interna (GPi), border cells (neurons at the boundary between the GPi and globus pallidus externa), and the NBM in sequence. Neurons in the GPi presented with a higher firing rate (mean firing rate 92.6±46.1 Hz) with a regular firing pattern (based on the burst index), while border cells fired at a lower rate (mean firing rate 34±21 Hz) with a regular pattern. When the microelectrode advanced to the NBM, firing signals of lower rate (mean firing rate 13±10 Hz) and irregular pattern (significantly greater burst index) were recorded, which provides a meaningful MER reference for NBM localization in DBS.
Target evaluation by quantitative EEG
Raw data can be easily acquired from simultaneous EEG and LFP recording during the operation for further quantitative EEG (qEEG) analysis of DBS target recognition. In AD studies, qEEG has been frequently used to analyze pathological alternation in the power spectrum or the brain functional connectivity during disease progression. Compared with the normal population, AD patients show a power decrease in the alpha band and an increase in the delta and theta bands in the cortex, especially in the frontal lobe [18]. Decreased coherence in the interhemispheric occipital alpha band or decreased prefrontal theta-gamma coupling can also distinguish AD recordings [101, 102]. For target evaluation, one study about NBM-DBS for a patient with PD dementia (PDD) revealed differences in the power spectrum and the coherence profile between situations when the electrode was positioned outside/within or near (<2 mm) the NBM. Recorded LFP presented significantly higher peak power and higher temporal coherence in the delta band, which makes qEEG a potential target detection method during surgery in clinical practice [103].
Is selection of the NBM subdivision possible with more effective AD treatment?
Because the NBM consists of groups of neurons with diverse projection patterns, various subdivisi-ons of the NBM dominantly innervate different areas of the corresponding cortex and modulate different cognitive fields. The intermediate sector of the NBM (Ch4 intermedius, Ch4i) represents the NBM’s lar-gest neuron group, which is easily located on a structural MRI by its proximity to the anterior commissure and the optic tract [31], and is often selected as tar-get for DBS. The research led by Freund et al., wh-ich localizing the tip of the electrode at Ch4i for PDD therapy, confirmed a prominent putative therapeutic effect by DBS. A study from Gratwicke’s team also admitted the advantage of Ch4i stimulation for PDD. As the largest component of the NBM, Ch4i provided the highest probability of successful electrode implantation and the most widespread cortical projections. Additionally, specific approaches to Ch4i avoid important structures near the basal forebrain, such as the optic tract [35]. Another study on NBM-DBS in AD tried to choose target areas extending from Ch4i medially to Ch4p (the posterior part of the NBM) in the subputaminal region. Unfortunately, it was pointed out that the insertion of electrodes into the preselected target may be difficult as a con-sequence of vascular alterations or pathological conditions such as hemorrhage, sublenticular cysts, or perivascular lacunae [30]. In spite of the difficulty in accurate localization to every subarea in the NBM, the intermediate portion connects with the ventrolateral orbital, insular, inferior temporal, and parahippocampal areas, as well as the inferior parietal lobule, making it a highly effective target in dementia treatment [33, 104].
TREATMENT EFFECTS OF NBM STIMULATION IN AD
NBM stimulation modifies neocortical excitability and reduces AD pathology burden
Stimulation of cholinergic fibers of the NBM can induce enhancement of neocortical gamma oscillation [83]. Regulation of the excitability of these fibers through DBS may also present certain topographical correspondence, as proven in fibers projecting to the dorsal frontal cortex. Sequential electrical stimulation applied to the NBM in an anteroposterior manner induces excitation in fibers arranged from the medial to lateral dorsal frontal cortex marked with voltage-sensitive dyes [105]. However, NBM lesions can result in an increase in event-related oscillation energy in delta and theta bands in the frontal cortex, and a decrease in gamma event-related oscillation energy in the frontal cortex and amygdala, similar to what has been recorded in AD patients [71]. In this case, the value of the NBM as a neuromodulation target for neocortical excitability speaks for itself. Gamma oscillation is credited with the electrophysiological index of cortical activity in learning and memory, the magnitude of which is modulated by cortical or subcortical slower rhythms [106]. Modulation of the cortex’s oscillation pattern by DBS to match the resonant frequency can strengthen the coupling of proper oscillation and imposed stimulation [107]. These studies indicate that stimulation of the NBM may benefit from boosted cortical gamma oscillation induced with low-frequency rhythms, as well as from coupling between the NBM and the neocortex.
In terms of long-term effect, NBM-DBS promotes an increase in cortical thickness that is observable on T1-weighted MRI, and presents positive correlation with preserved Mini-Mental State Examination (MMSE) score in AD patients when comparing the baseline to six and 12 months after surgery [34]. In another study, three out of four AD patients exhibited improved parieto-temporal glucose metabolism on 18F-FDG-PET one year after NBM-DBS [30]. Moreover, NBM-DBS stimulation was proven to reduce the AD pathology burden in one APP/PS1 mouse study by Huang et al. Levels of soluble Aβ40 and Aβ42 in the hippocampus and cortex were suppressed significantly after 21 days of stimulation. Additionally, more neuron survival and downregulation of the expression of pro-apoptosis proteins, such as caspase-3 and BH3-interacting domain death agonist (Bid), were also confirmed [37].
Regenerative effects of NBM stimulation
Cholinotrophic defects and circuitry dysfunction play essential roles in AD development and are partly mediated by the dysregulation of nerve growth factor (NGF) function. In the cortex, NGF binds to the tropomyosin-related kinase A (TrkA) receptor and the low-affinity neurotrophin receptor P75NTR (p75NTR) receptor, which conduct survival and apoptosis signals, respectively. Compared with normal individuals, in MCI and AD patients, the TrkA receptor is downregulated in the neurons projecting from the NBM to the neocortex [108]. Clinical trials also supported NGF-targeted therapy in AD treatment. Researchers attempted through different methods, such as drug delivery of recombinant human NGF and ex vivo or in vivo gene transfer to specific neurons, to increase the level of NGF in the cortex of AD patients [109, 110]. Hotta et al. first provided evidence that stimulation of the NBM in the adult rat can affect the neocortical extracellular environment by promoting NGF secretion [111]. Further study has shown that basal forebrain stimulation can induce NGF secretion in the parietal cortex by cholinergic innervation in adult rats [112]. In essence, the NBM and the neocortex reciprocally support the functionality and survival of each other, which demonstrates the regenerative function of the brain as a whole [107]. Apart from NGF released by the neurons, astrocytes in the local cortical area are also involved in the regenerative effect of DBS by modulating cortical plasticity via calcium signaling [113].
Role of NBM stimulation in improving or delaying the progression of cognitive dysfunction
In the early days, experiments on normal rats showed that NBM stimulation may facilitate the early stage of memory formation [114, 115]. Recently, Koulousakis and colleagues, who investigated the stimulation pattern and effect, have found that bilateral intermittent NBM-DBS can elicit supernormal performance in transgenic AD rats in a novel object recognition task, and a modified Barnes maze task [38]. These results provide primary evidence for the therapeutic effect of NBM-DBS in the mitigation of cognitive impairment in AD.
Until the present, several preclinical trials with li-mited cohort scales on NBM-DBS for dementia patients had been undertaken. In their research conducted in 2009, Freund et al. treated a 71-year-old PDD patient with bilateral NBM-DBS, and they also placed two electrodes in the subthalamic nuc-leus (STN) and found notable improvement in the patient’s performance on the clock drawing task, trail making test, auditory verbal learning and memory test, and verbal fluency. Also noteworthy is their finding that after 24 h with the stimulation temporarily turned off, cognitive improvement was canceled [31]. Moreover, in Kuhn’s study, six patients with mild-to-moderate AD received bilateral low-frequency DBS of the NBM with 12 months of follow-up, resulting in insignificant increase of 3 points in the ADAS-Cog score, a decrease in MMSE score by 0.5 points, comparing to a previous research which shown that natural progression of AD can lead to the decline of scores in MMSE by 1.84 per year according to the regression model [116]. The study also revealed an unchanged clinical dementia rating of 0.83±0.3 on average [30]. The research led by Gratwicke and colleagues on six patients with PDD reported positive result that the total score of Neuropsychiatric Inventory decreased by 5.00 at median with significance, confirming the treatment effect of DBS on psychiatric symptoms in AD. However, no significant differences were found in cognitive scales including MMSE and MDS-UPDRS between NBM-DBS-on and off state [35]. Taken together, these inconsistent preliminary results indicate that NBM-DBS may prevent the deterioration of the cognitive scores and maintain a relatively stable cognitive state. However, a definite conclusion for feasible effective stimulation protocol and specific treatment mechanisms require further study. Treatment effects of NBM-DBS in psychiatric symptoms of AD are also noteworthy as suggested.
SHORTCOMINGS AND FUTURE DIRECTIONS OF NBM-DBS THERAPY IN AD
Despite the progress in selecting a definite target, developing a feasible operation procedure, and findings of a preliminary therapeutic effect, DBS as a therapy for AD has inherent shortcomings that can mainly be attributed to the invasive nature and specific projections of the NBM. As an invasive method, DBS electrode implantation causes tissue damage and radiofrequency (RF) heating on the metal-tissue interface in the MRI environment. The NBM contains complex and widespread projection to the frontal, insular, and temporal lobes, and thus NBM stimulation is prone to induce side effects. Stimulation on one of the NBM’s four adjacent subdivisions with distinct projection patterns also requires clear discrimination to ensure a surgery benefit. The current review focuses on the shortcomings, which may influence the therapeutic results of NBM-DBS, and may be soluble with revised designs to the equipment and the procedure.
Variations on regional cortical blood flow
NBM stimulation can induce various side effects, one of which is variation in regional cortical blood flow. Activation of the NBM’s cholinergic neurons was demonstrated to elevate regional cortical blood flow, which occurred independent of metabolic changes [117]. The vasodilative activity was then ascribed to the possibility of NBM projection directly to the microvessel, or to the astrocytes circumscribing them, in the cortex [118, 119]. NBM dysfunction may explain the abnormal cortical circulation in AD, and animal experiments have demonstrated the protective effect of NBM stimulation after transient ischemia. However, NBM stimulation can also alter the perfusion condition in affected areas abruptly [120]. Further investigation is required to determine whether hemodynamic changes induce subsequent impairment and compromise the therapeutic effect.
Focal temperature increase in stimulation target area
Attention was also paid to the focal heat damage caused by electric stimulation to the brain tissue in previous studies. During the stimulation, Joule heat is generated and the target temperature increases. The temperature fluctuation depends on conditions including the waveform, lead selection, conductivity of the brain tissue, blood perfusion, and metabolic state [121]. As the temperature increases higher than normal, influence on the brain tissue and the network occurs. Compared with the normal state, a temperature raised by 0.5°C can cause changes in cell excitability, while a temperature raised by 1°C can induce alteration in synaptic transmission as well as blood-brain barrier function. Temperatures higher than 39°C lasting for hours can probably damage circuitry function [122, 123]. However, in a computer simulation with finite element models, this thermal effect induces temperature rise by 0.8°C at most under the clinical DBS protocol [121, 124], which indicated that the regular clinical DBS is safe and will not cause damage with clinical meaning.
Apart from the direct electrical heat, the DBS device, including the electrode, the wire, and the battery, can become heated during the MRI due to the effect of RF heating. A NaCl solution-filled model has substantiated the finding that common MRI sequences might be potentially hazardous to individuals with DBS devices [125]. In an earlier study, 10 patients with implanted DBS devices received 3-Tesla MRI scanning with T2-weighted sequence and presented no heating or distinguishable neurological adverse effects [126]. In spite of the significant artifact and the small cohort, this study provided preliminary evidence for the safety of MRI scanning with DBS devices. A further phantom study suggested that among different sequences, T2-weighted sequences may cause higher heating due to a higher specific absorption ratio. Utilizing the Medtronic DBS lead model 3387, temperature increase measured in 3-Tesla MRI was no more than 2°C with different sequences [127]. Therefore, the researcher suggested avoiding the application of T2 sequences in patients with DBS devices. To reduce RF heating, recently, a research group experimented on the reconfigurable MRI coil (coil with changeable orientation) for patients with bilateral leads, and achieved a reduction of 65%average RF heat for bilateral leads (and 80%for the unilateral) [128], indicated that this novel technology hopefully can assist in the optimization of MRI scanning for DBS patients.
Therefore, although the conventional parameter setting for DBS stimulation shows enough safety in terms of thermal effect, surveillance of electrode temperature may serve as an improved measure to guarantee intraoperative and postoperative security, especially for centers carried on MRI evaluation during or after DBS regularly.
Stimulation-related tissue damage
Tissue damage inevitably occurs during DBS operations. This damage can include hemorrhage and microelectrode penetration. In the period following stimulation, infection, lead migration, breakage, or erosion may occur [129]. Furthermore, electrical stimulation induces a tissue response that can cause extra damage around the electrode. Faradic electrochemical reaction occurs on the interface between the electrode and brain tissue. The strength of these oxidization and reduction reactions depends on multiple factors. The material of the electrode is one of the major factors. Stainless steel electrodes can induce tissue damage even when following the same design and stimulation parameters as their platinum-iridium counterparts. Therefore, platinum-iridium alloy coated with polyurethane jacket is gradually accepted as a safer DBS electrode [130, 131]. Charge density of the electrode is another factor that affects stimulation safety. Relative in vitro research has recommended 50μC/cm2 as a threshold. In experiments exceeding this threshold, destruction of the blood-brain barrier has been observed. Additionally, charge density over 30μC/cm2 has been found to clearly increase the risk of tissue damage [132, 133]. Apart from proper electrode material and charge density control, safe stimulation also requires appropriate stimulation waveform and interval [129, 133]. For tissue damage due to long-term stimulation, Gaddum et al. reviewed postmortem anatomic studies of DBS for PD treatment, which revealed that fibrosis and gliosis surrounding the electrode could appear in almost all patients. Furthermore, among the 57 cases collected, capsule formation was found in 19 cases, while microvasculature was found in 5 cases [134]. In another early postmortem case for thalamus stimulation, neuronal degeneration and vacuolization were also verified around the electrode site [135].
Neural complications related to NBM stimulation
Considering the complicate widespread projections from the NBM and the individual variability of basal forebrain nuclei, stimulation of this target can hardly avoid side effects caused by activation of cholinergic or even non-cholinergic fibers. Thus far, several clinical trials of NBM-DBS have been launched to treat different diseases, and some have reported on the neural complications related to stimulating this target.
In Kuhn’s study of NBM-DBS in six AD patients, one subject complained about inner restlessness at higher stimulation intensities (>5 V) [30]. In fact, Wolz’s research about DBS targeting the STN to treat non-motor PD symptoms also reported inner restlessness in 53%of 34 subjects. However, when stimulation began, this proportion decreased to 31%(p = 0.008) and the severity of restlessness reduced from 28.8%(±30.8%) to 11.5%(±21.2%) (p = 0.001) (severity of this complication was scaled with a visual analogue scale ranging from 0%[no symptoms] to 100%[most severe symptoms]) [136]. Additionally, this symptom should be differentiated from akathisia or restless leg syndrome, although these manifestations are rarely seen in AD patients. The patient’s symptom of inner restless in Kuhn’s study improved after downregulation of the voltage, although the relation between high voltage stimulation and inner restlessness is unclear.
In Gratwicke’s study of PDD patients treated by NBM-DBS, one of six subjects reported visual hallucinations after stimulation [35]. The projection of NBM fibers may indicate the source of visual hallucination. As the NBM lies under the lentiform nucleus stretching horizontally, the tip of the electrode, which advances obliquely, may protrude into the part of the NBM posterior to Ch4i, namely, Ch4p. Ch4p sends cholinergic projections to the superior temporal gyrus and the temporal pole, where stimulation of Ch4p can induce hallucinations and delusions [137].
CONCLUSIONS
The current review focuses on NBM-DBS as a feasible surgical neuromodulation therapy for AD with great promise. Featuring widespread projections, the NBM is involved in the regulation of memory, attention, and arousal state. As AD progresses, the NBM suffers from neuronal loss as a critical component of the cholinergic system. Using a stereotactic system, an MRI with multiple sequences, and microelectrode recording, growing accuracy has been realized in target localization. Although preclinical trials on NBM-DBS have not been conclusive due to their limited samples, the treatment effect of NBM-DBS on cognition is worth investigation. There are also important issues waiting to be addressed. Patient selection, or indication for NBM-DBS, should be fur-ther considered to improve efficacy and obtain a fav-orable prognosis. Furthermore, the projection relationships of the NBM are highly complicated, which requires target refinement as well as individual parameter optimization. Moreover, studies designed to improve the stimulation instrument and focal neural damage control during NBM-DBS treatment for AD should improve the application of this surgical neuromodulation therapy in future clinical practice.
Footnotes
ACKNOWLEDGMENTS
This work was funded by the National Natural Science Foundation of China, grant number 81974279, China association against epilepsy fund for epilepsy research-UCB fund, grant number 2020020A, Grants for returned overseas doctors of Tongji Hospital, Grants for teaching research of Huazhong University of Science and Technology and the Fundamental Research Funds for the Central Universities, grant number 2019kfyXKJC069.