
Abstract
Background:
Neurofibrillary pathology of abnormally hyperphosphorylated tau spreads along neuroanatomical connections, underlying the progression of Alzheimer’s disease (AD). The propagation of tau pathology to axonally connected brain regions inevitably involves trafficking of seeding-competent tau within the axonal compartment of the neuron.
Objective:
To determine the seeding activity of tau in cerebral gray and white matters of AD.
Methods:
Levels of total tau, hyperphosphorylation of tau, and SDS- and β-mercaptoethanol–resistant high molecular weight tau (HMW-tau) in crude extracts from gray and white matters of AD frontal lobes were analyzed by immuno-blots. Tau seeding activity was quantitatively assessed by measuring RIPA buffer–insoluble tau in HEK-293FT/tau151-391 cells treated with brain extracts.
Results:
We found a comparable level of soluble tau in gray matter versus white matter of control brains, but a higher level of soluble tau in gray matter than white matter of AD brains. In AD brains, tau is hyperphosphorylated in both gray and white matters, with a higher level in the former. The extracts of both gray and white matters of AD brains seeded tau aggregation in HEK-293FT/tau151–391 cells but the white matter showed less potency. Seeding activity of tau in brain extracts was positively correlated with the levels of tau hyperphosphorylation and HMW-tau. RIPA-insoluble tau, but not RIPA-soluble tau, was hyperphosphorylated tau at multiple sites.
Conclusion:
Both gray and white matters of AD brain contain seeding-competent tau that can template aggregation of hyperphosphorylated tau, but the seeding potency is markedly higher in gray matter than in white matter.
INTRODUCTION
Neurofibrillary tangles (NFTs) of abnormally hyperphosphorylated tau are a histopathological hallmark of Alzheimer’s disease (AD) [1–3]. The number of NFTs is positively correlated with cognitive impairment in patients with this disease [4–6]. Tau was initially described as a neuronal microtubule-associated protein that is enriched in the axon [7, 8]. Its presence within dendrites and in the post-synapse, both under physiological and pathological conditions, has been discovered recently [9]. The major known function of tau is to promote microtubule assembly and stabilize microtubule structure. Tau is a phospho-protein, and phosphorylation regulates its association with microtubules. However, in AD brain, tau is abnormally hyperphosphorylated, which is believed to be responsible for its loss of biological activity, its gain of toxic activity, and its aggregation into paired helical filaments (PHFs) in NFTs [10–12].
Tau lesion, in AD brain, is found sequentially in the transentorhinal–entorhinal areas, hippocampus, limbic areas, and, finally, in the associative and then primary neocortical areas [13, 14], which is associated with the progression of AD [14]. Recently, tau tracer retention measured by positron emission tomography also showed similar progression [15–17]. Thus, tau pathology in AD brain may spread along neuroanatomical connections. Injection of brain extract from tauP301S transgenic mice into the brain of wild-type human tau-expressing mice induced tau pathology not only at the injection sites, but also in the anatomically connected brain regions in a time-dependent manner [18], resulting in the concept of propagation of tau pathology. Following this study, the progressive propagation of tau pathology has been replicated in various animal models, including those using regional promoters, inoculation models, and viral models [19–25]. Tau pathology apparently radiates through the brain along synaptically connected pathways as the disease progresses. Thus, tau in dendritic and exonal processes appears to play a critical role in the propagation of tau pathology between brain regions. However, the seeding activity of tau in AD axons was not documented.
The cerebrum is made of an outer layer of gray matter and the core of white matter. Gray matter is distinguished from white matter in that it contains numerous cell bodies and relatively few myelinated axons, whereas white matter contains relatively few cell bodies and is composed chiefly of long-range my-elinated axons [26]. In the present study, we assessed the seeding activity of tau in the crude extracts of gray and white matters of AD and control frontal labes. We found accumulation of hyperphosphorylated tau in gray matter, but not in the white matter of AD cerebrum. Extracts of AD gray and whiter matters seeded aggregation of hyperphosphorylated tau in cultured cells, but gray matter extracts displayed more potent seeding activity than did white matter extracts. Seeding activity of tau was positively correlated with the level of hyperphosphorylated tau and SDS- and β-mercaptoethanol (ME)–resistant high-molecular weight tau (HMW-tau) in the cerebral extracts. These findings provide direct evidence of the presence of seeding-competent tau in both gray and white matters of AD brain, which templates tau aggregation, but is less potent in white matter.
MATERIALS AND METHODS
Human brain tissues and brain tissue processing
The frontal lobes of six AD and five age- and pos-tmortem delay-matched control brains used in this study were obtained without identification of the donors from the Netherland brain bank. All brain samples were histopathologically confirmed and stored at -80°C until use. The use of frozen human brain tissue was in accordance with the National Institutes of Health guidelines and was exempted by our institutional review committee because “the research does not involve intervention or interaction with the individuals” nor “is the information individually identifiable”.
Gray matter and white matter of frontal lobe were separated on ice as described previously [27] and homogenized with Teflon-glass homogenizer in 9×volumes of cold buffer (20 mM Tris-HCl, pH7.6, 0.32 M sucrose, 5 mM benzamidine, 10μM β-ME, 5 mM EGTA, 0.5 mM MgSO4, 50 mM glycerophosphate, 1 mM EDTA, 50 mM NaF, 1 mM Na3VO4, 0.6 mM PMSF, and 10μg/ml each of aprotinin, leu-peptin, and pepstatin). The homogenates were centrifuged at 10,000×g for 10 min, and the supernatant was collected as crude extract. The crude extracts containing soluble tau were stored at –80°C for further analysis.
Cell culture and transfection
HEK-293FT cells (human embryonic kidney cell line) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Thermo Fisher Scientific, Wal-tham, MA, USA), supplemented with 10%fetal bovine serum (FBS) (Thermo Fisher Scientific), 100 U/ml penicillin, and 100μg/ml streptomycin, in a humidified atmosphere containing 5%CO2 at 37°C. Cells were seeded to culture plates, and all transfections were performed with FuGENE HD (Promega, Madison, WI, USA) according to the manufacturer’s instructions.
Western blots and immuno-dot blots
For western blots, the brain extracts and cell lysates were mixed with 4×Laemmli sample buffer (125 mM Tris-HCl, pH 6.8, 2%SDS, 10%glycerol, 10%β-ME, 0.004%bromphenol blue) containing 10μg/ml each of aprotinin, leupeptin, and pepstatin, followed by heating in boiling water for 5 min. Protein concentration of samples was assessed by using the Pierce™ 660 nm Protein Assay Kit (Thermo Fisher Scientific). Samples were subjected to SDS-PAGE and were electro-blotted onto polyvinylidene fluoride membrane (PVDF) (Millipore Sigma, Burlington, MA, USA). The membrane was subsequently blocked in 5%fat-free milk-TBS for 30 min, incubated with primary antibodies listed in Table 1 in 5%milk-TBS overnight, washed with TBST (50 mM Tris-HCl, 150 mM NaCl with 0.1%Tween20), incubated with HRP-conju-gated secondary antibody for 2 h, washed with TBST, and incubated with the ECL Western Blotting Substrate (Thermo Fisher Scientific) and exposed to HyBlot CL® autoradiography film (Denville Scientific, Inc., Holliston, MA, USA). Specific immunoreactive signal was quantified by using the Multi Gauge software V3.0 from Fuji Film (Minato, Tokyo, Japan).
Primary antibodies used in the present study
Mono-, monoclonal; p-, phosphorylated; Poly-, polyclonal; M, mouse; R, rabbit.
For immuno-dot blots, the brain crude extracts, adjusted to same protein concentration, were diluted serially with dilution buffer (0.2%BSA in TBS containing 50 mM NaF, 1 mM Na3VO4, and 2μg/ml each of aprotinin, leupeptin, and pepstatin) and app-lied onto nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA) at 5μl/grid (7×7 mm). AD O-tau from an AD brain (case 4222) prepared previously [28] was used as positive control. The blot was placed in a 37°C oven for 1 h to allow the protein to bind to the membrane and was processed as described above for western blots.
Seeding activity assay in cultured cells
Seeding activity in cultured cells was assayed as described previously [29]. Briefly, HEK-293FT cells seeded in 24-well plate were transfected with pCI/HA-tau151–391 with FuGENE HD. Crude ext-racts of gray or white matter from AD and control brains were filtered with 0.2μm filter (Millipore Si-gma). The level of tau in brain extracts was analyzed by immuno-dot blots developed with a mixture of two tau pan-antibodies, R134d and 92e, and were adjusted to the same tau levels with the homogeni-zing buffer. The crude extract was mixed with Li-pofectamine 2000 in Opti-MEM (Thermo Fisher Scientific) and incubated for 20 min at RT. The mix-ture was added into the cell cultures after 6 h transfection and cultured for 42 h. The AD O-tau from AD case 4222 [25, 30] was used as positive control. The cells were lysed in RIPA buffer (50 mM Tris-HCl, 150 mM NaCl, 1%NP–40, 0.5 %sodium deoxycholate, and 0.1%SDS) containing 50 mM NaF, 1 mM Na3VO4, 1.0 mM AEBSF, and 10μg/ml each of aprotinin, leupeptin, and pepstatin for 20 min on ice. The cell lysates were centrifuged at 100,000×g for 30 min, and the resulting pellet was washed twice with RIPA buffer. The supernatants were pooled together as RIPA-soluble fraction, and the pellet was the RIPA-insoluble fraction. Levels of RIPA-insoluble and -soluble tau were analyzed by western blots developed with anti-HA and site-specific and phosphorylation-dependent tau antibodies.
Statistical analysis
The GraphPad Prism 6 software was used for statistical analysis. Data were analyzed by two-way ANOVA followed by Sidak’s multiple comparisons and by Pearson correlation analysis.
RESULTS
Higher level of soluble tau in AD gray matter than in white matter
Tau is a cytosolic protein. In AD and related tauopathies’ brain, hyperphosphorylated tau aggregates into NFTs in cell bodies and neuropil threads in neuronal processes. Hyperphosphorylated cytosolic tau (AD P-tau) from AD brain, but not PHF-tau, sequesters normal tau in vitro [10]. To determine the level of soluble tau in gray and white matter, we prepared 10,000×g crude extract of gray and white matter from the frontal lobes of AD cases and age- and postmortem delay–matched control cases and analyzed tau by immuno-dot blots developed with a mixture of pan-tau antibodies, R134d and 92e (Fig. 1A). We found a similar level of tau in gray and white matter in control brains, but a higher tau level in gray than white matter in AD cases (Fig. 1A, B). The level of tau in AD gray matter was markedly elevated as compared with control gray matter, but tau level in AD white matter was not significantly different from that in control white matter (Fig. 1A,B). 2’,3’-cyclic-nucleotide 3’-phosphodiesterase (CNPase), a myelin-associated enzyme [31], was predominantly expressed in white matter (Fig. 1C). Thus, these data indicate accumulation of soluble tau in AD gray matter, which consists predominantly of neuronal bod-ies, but not in AD white matter made up mainly of axons.
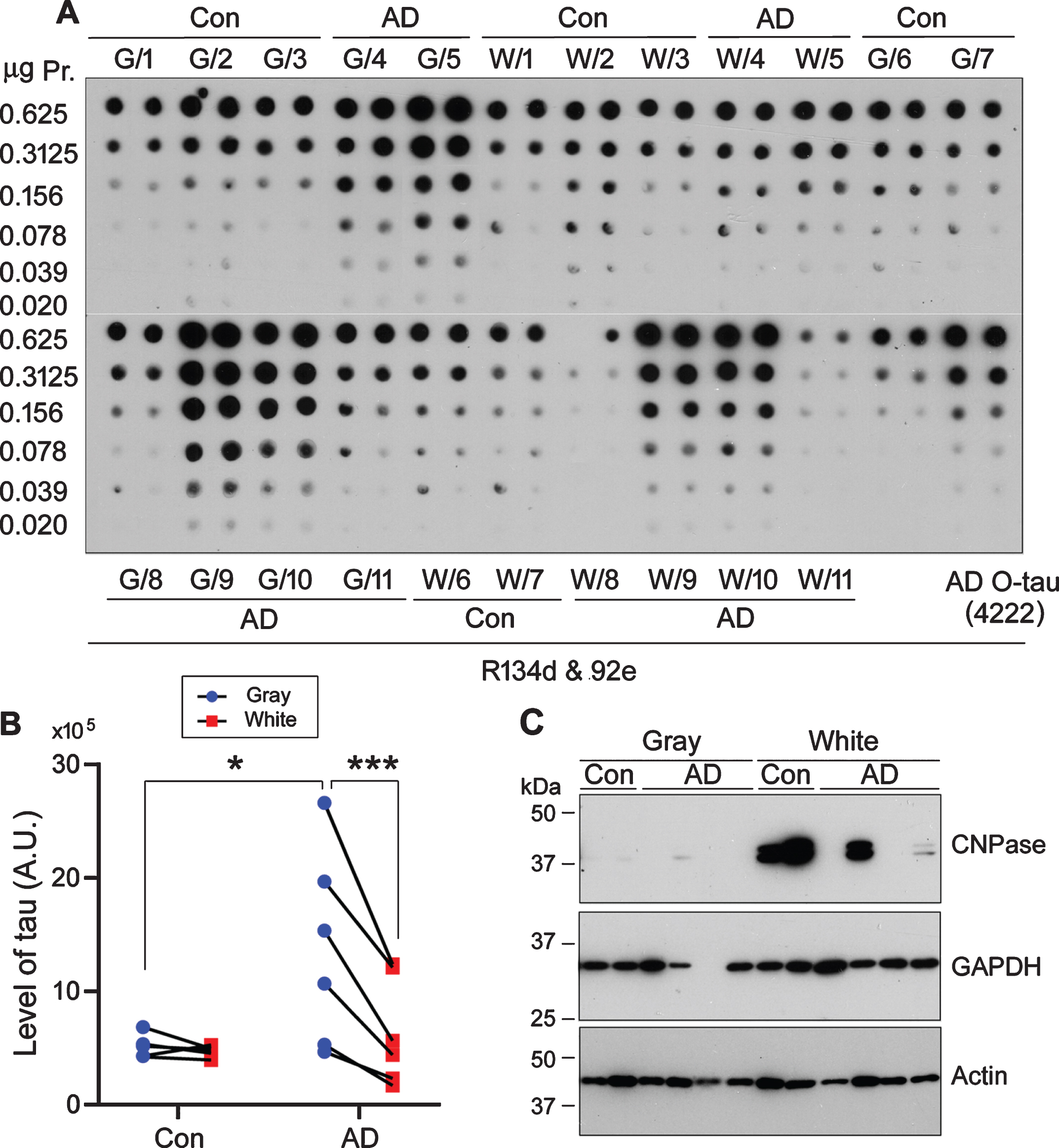
Level of soluble tau in gray and white matter of AD brain is higher than in corresponding fractions of control brains. A) 10%homogenates of gray matter (G) or white matter (W) of frontal lobe from 5 control (Con) and 6 AD cases were centrifuged at 10,000×g for 10 min. The supernatant saved as crude extract containing soluble tau was diluted serially and subjected for immuno-dot blot developed with a mixture of pan-tau antibodies R134d and 92e. B) The level of soluble tau in panel A was quantitated and analyzed with two-way ANOVA followed by Sidak’s multiple comparisons. The data are presented as relative levels of tau in gray and white matter from each case. *p < 0.05, ***p < 0.001. A.U., arbitrary units. C) Crude extracts of gray and white matters from 2 control (Con) and 4 AD cases were subjected for western blots developed with anti-CNPase, anti-GAPDH and anti-β-actin.
Higher levels of hyperphosphorylated tau and SDS- and β#x03B2;-ME–resistant high molecular weight tau (HMW-tau) in AD gray matter than in AD white matter
Tau is abnormally hyperphosphorylated in AD brain [1, 32]. To learn the level of hyperphosphorylation of tau in gray and white matter, we carried out western blots developed with site-specific and phosphorylation-dependent tau antibodies (Fig. 2A). We found that phosphorylated tau at Ser202/Thr205 (AT8), Thr217, Thr231, and Ser396/Ser404 (PHF-1) was undetectable in either control gray matter or white matter (Fig. 2A,B), whereas the level of hyp-erphosphorylation of tau was significantly higher in AD gray matter than in AD white matter and in control gray matter at all studied sites (Fig. 2A,B). Monoclonal tau antibody 77G7 recognizes microtu-bule-binding repeats and showed higher affinity to-ward tau in AD brain than in control brain [29]. We found that 77G7 immunoreactivity was markedly high in the AD gray matter (Fig. 2A,B). Thus, soluble tau was abnormally hyperphosphorylated in AD gray and white matter, but more severe in gray than white matter.
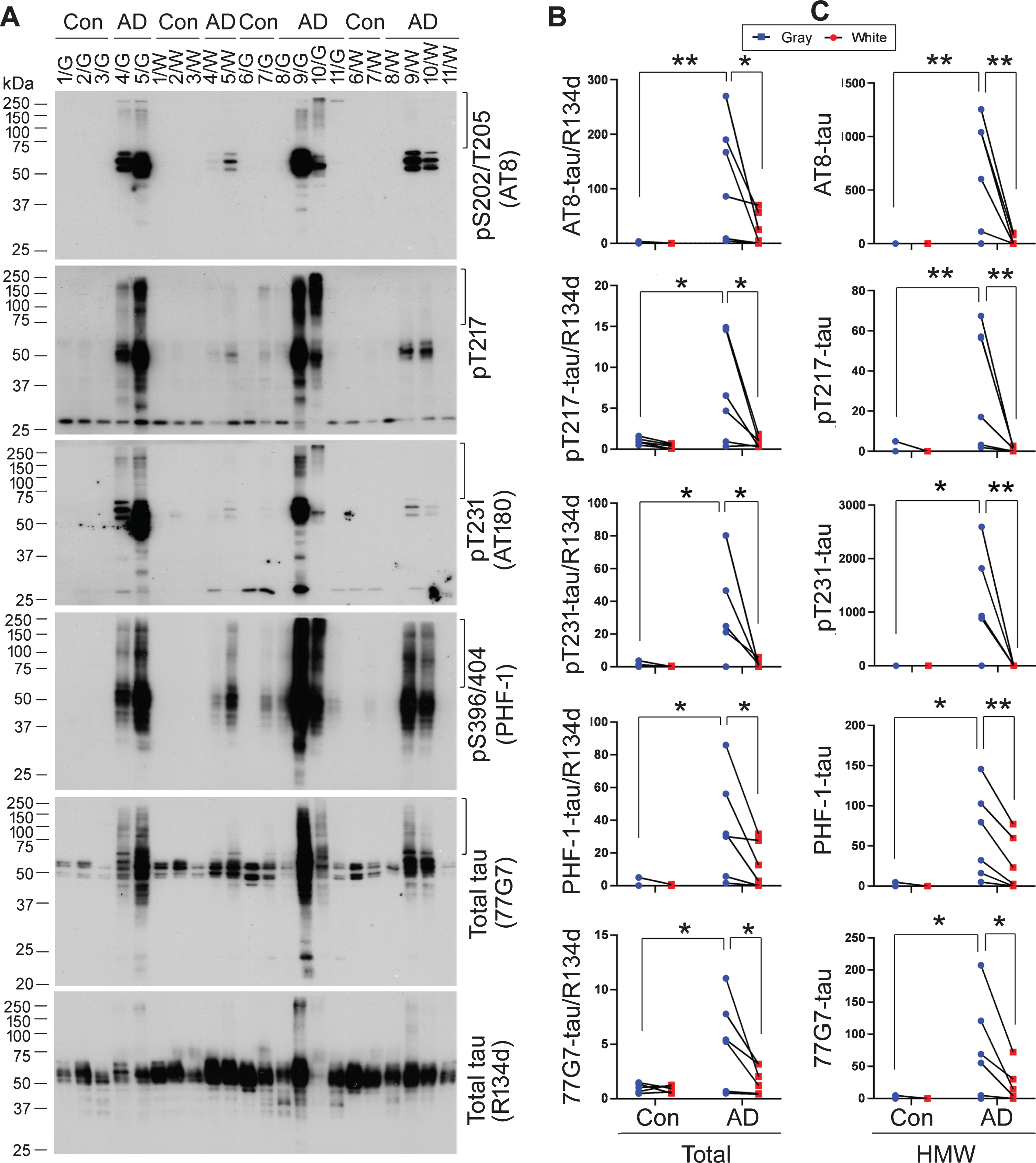
Level of hyperphosphorylated tau in AD gray and white matter is higher than in corresponding fractions of control brains. A) Hyperphosphorylation of tau in gray (G) and white (W) matters from five control (Con) and six AD brains was analyzed by western blots developed with site-specific and phosphorylation-dependent tau antibodies, monoclonal tau antibody 77G7 against the microtubule-binding repeats, and a pan-tau antibody, R134d. B,C) Levels of 77G7-tau and hyperphosphorylation of tau in brain extract normalized with R134d and in SDS- and β-ME–resistant high molecular weight (HMW), labeled with vertical mark at the right side of each blot in panel A, were quantitated by densitometry. Data are presented as relative level in gray and white matter from each case. The data were analyzed with two-way ANOVA followed by Sidak’s multiple comparisons. *p < 0.05, **p < 0.01.
In addition to abnormal hyperphosphorylation of tau, we found SDS- and β-ME–resistant high molecular weight tau (HMW-tau), a common feature of AD and related tauopathies [29, 33], in gray and/or white matter of most AD cases and only in one control case (Fig. 2A). The HMW-tau was found to immunoreact with all anti-phospho-tau antibodies (Fig. 2A), suggesting that it is hyperphosphorylated at multiple sites. Levels of hyperphosphorylated HMW-tau at Ser202/Thr205, Thr217, Thr231, and Ser396/404 were significantly higher in AD gray than white matter and in control gray matter (Fig. 2C). HMW-tau level in AD white matter was not significant but showed a trend to be higher than in control white matter at all studied phospho-sites. 77G7 immunoreactive HMW-tau also was elevated in AD cases and higher in AD gray matter than in white matter. These results suggest accumulation of hyperphosphorylated and SDS- and β-ME–resistant HMW-tau in AD brain, especially in the gray matter.
Higher seeding activity of soluble tau in AD gray matter
To determine the seeding activity of tau in brain extract, we employed a new method, in which HA-tau151–391 aggregation was induced by tau seeds in cultured cells and analyzed by immuno-blots in RIPA –insoluble fraction [29]. We overexpressed HA-tau151–391 in HEK-293FT cells and treated them with extracts of gray and white matter, with the same amount of tau as determined by immuno-dot blots, after 6 h transfection. After 42 h culture, the cells were lysed in RIPA buffer. The RIPA-soluble and -insoluble fractions were separated by centrifugation of cell lysates at 100,000×g for 30 min. Aggregated tau yielded in the RIPA-insoluble fraction, which reflects seeding activity of tau, was analyzed by western blots developed with anti-HA (Fig. 3A). We found a similar level of RIPA-soluble tau (Fig. 3A,B), but a markedly different level of RIPA-insoluble tau in the cells treated with the above brain extracts (Fig. 3A,C). Interestingly, tau in the RIPA-soluble fraction showed one band, but tau in the RIPA-insoluble fraction showed multiple bands in the blots (Fig. 3A). Very little or no RIPA-insoluble tau was found in the cells treated with extracts from control gray matter or white matter (Fig. 3A,C), and markedly high levels were found in cells treated with extracts of AD gray matter or white matter (Fig. 3A,C). Compared with control brain extract treatment, AD gray matter extract significantly increased the level of RIPA-insoluble tau (Fig. 3C), but AD white matter extract did not significantly increase RIPA-insoluble tau (Fig. 3C). Moreover, the level of RIPA-insoluble tau in cells treated with the extracts of AD gray matter was significantly higher than that treated with the corresponding AD white matter (Fig. 3A,C). These results suggest that AD gray and white matter contains seeding-competent tau to template tau aggregation, but soluble tau in AD gray matter displays much stronger seeding activity than in the white matter from the same area of the brain.
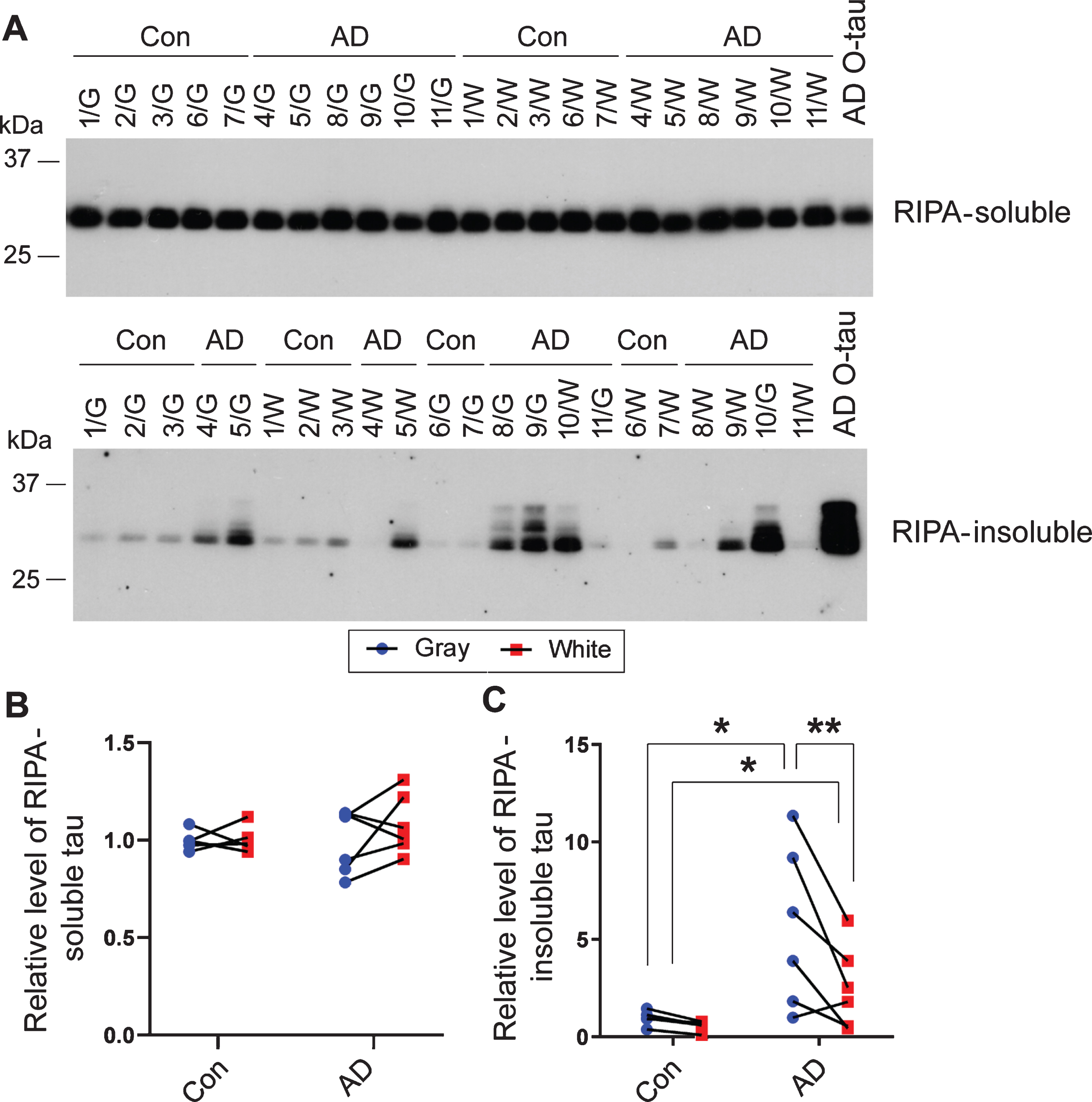
Soluble tau in AD gray matter and white matter seeds tau aggregation in cultured cells. A) HA-tau151–391 was expressed in HEK-293FT cells for 6 h. The cells were treated with the extracts of gray (G) or white (W) matter from AD and control (Con) brains with the same amount of tau for 42 h and were lysed in RIPA buffer. RIPA buffer-soluble and -insoluble fractions were separated by centrifugation at 100,000×g for 30 min and analyzed by western blots developed with anti-HA. AD O-tau was used as positive control. B, C) Relative levels of RIPA-soluble tau (B) and -insoluble tau (C) in panel A were quantified by densitometric analysis. Data are presented as relative levels in gray and white matter from each case and were analyzed with two-way ANOVA followed by Sidak’s multiple comparisons. *p < 0.05; **p < 0.01.
Positive correlation between seeding activity of tau and tau hyperphosphorylation in gray and whiter matter
To learn the association of seeding activity of tau with its hyperphosphorylation, we performed Pearson correlation analysis. We found that the level of RIPA-insoluble tau reflecting seeding activity was positively correlated with the levels of hyperphosphorylation of tau at Ser202/Thr205 (AT8), Thr217, and Ser396/404 (Fig. 4A), suggesting that in the brain, hyperphosphorylation of tau is positively associated with its seeding activity.
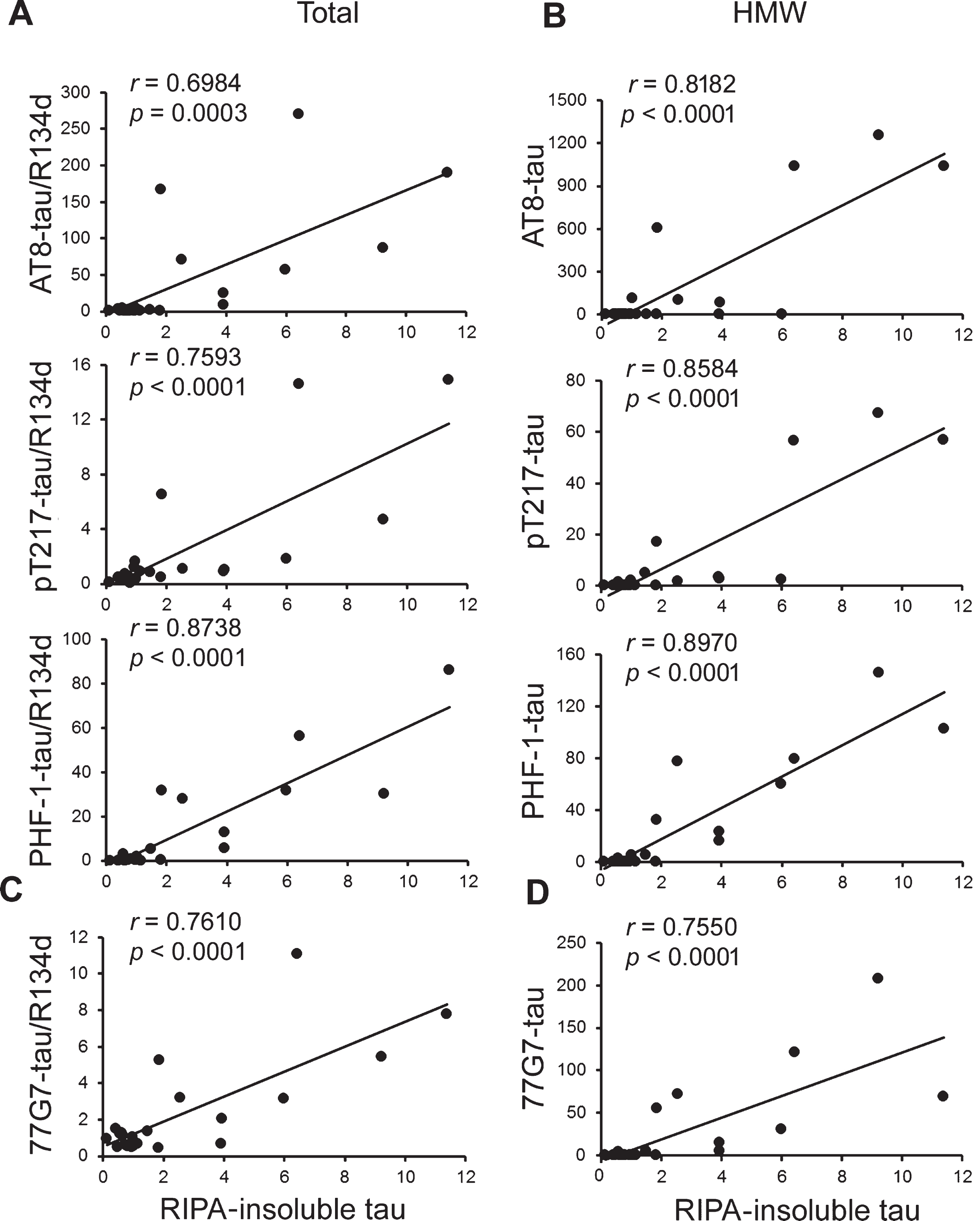
The seeding activity of tau positively correlates with the levels of tau phosphorylation and HMW-tau in brain extracts. A, B) The levels of phosphorylated tau normalized with R134d (A) or phosphorylated HMW-tau (B) in the above extracts determined in Fig. 2 and the level of RIPA-insoluble tau shown in Fig. 3 were subjected to Pearson correlation analysis. C, D) Pearson correlation analysis was performed between the levels of total and HMW tau determined by 77G7 in Fig. 2 with the level of RIPA-insoluble tau shown in Fig. 3.
Phosphorylated HMW-tau is known to be up-taken more effectively by neurons and to seed tau aggregation [34]. We analyzed the relationship between the seeding activity and the level of SDS- and β-ME–resistant hyperphosphorylated HMW-tau. We found that the level of RIPA-insoluble tau in cells treated with the above brain extracts was positively correlated to the level of HMW-tau hyperphosphorylated at Ser202/Thr205, Thr217, and Ser396/404 (Fig. 4B). Thus, hyperphosphorylated HMW-tau is positively associated with tau seeding activity.
We previously found that antibody 77G7 to total tau had higher affinity to tau from AD brain than control brain [29]. We found that the level of RIPA-insoluble tau was positively correlated with 77G7 immunoreactive tau and HMW-tau (Fig. 4C,D).
Moreover, we found that the Pearson coefficient r values were larger with phospho-HMW-tau than with phospho-tau (Fig. 4), suggesting a slightly stronger association of tau seeding activity with HMW-tau than total phospho-tau.
Hyperphosphorylation of RIPA-insoluble aggregated tau induced by AD brain extract
NFTs are made up of abnormally hyperphos-phorylated tau [32]. Phosphorylation alters tau migration in SDS-PAGE [35]. RIPA-soluble tau and RIPA-insoluble tau displayed one band and multiple bands, respectively (Fig. 3A), suggesting that tau is present in various phosphorylation statuses. Thus, we analyzed hyperphosphorylation of RIPA-soluble and -insoluble taus by western blots developed with site-specific and phosphorylation-dependent tau antibodies. We found that the level of hyperphosphorylated tau in the RIPA-soluble fractions was similar or slightly altered in cells treated with gray and white matter extracts from AD and control brains (Fig. 5A,B). In contrast, in RIPA-insoluble fractions of AD extract-treated cells, the levels of phosphorylated tau at Thr181, Ser199, Ser202/Thr205, Ser214, Thr217, and Thr231 were increased (Fig. 5C,D). The increased hyperphosphorylated RIPA-insoluble tau was more in AD gray matter than in AD white matter–treated cells (Fig. 5D). These results indicate that tau in the aggregates, but not in soluble fraction, is hyperphosphorylated.

Soluble tau in AD cerebral gray and white matter induces aggregation of hyperphosphorylated tau. A, B) HA-tau151–391 was expressed in HEK-293FT cells for 6 h, and then the cells were treated with extracts of gray (G) or white (W) matter from control (Con) or AD brains for 42 hr. RIPA buffer–soluble (A) and -insoluble (C) fractions were subjected to western blots developed with site-specific and phosphorylation-dependent tau antibodies. B, D) Relative levels of hyperphosphorylated tau were quantified and are presented in gray and white matter from each case. E) HEK-293FT/HA-tau151–391 cells were treated with extracts of gray or white matter from AD or control (Con) brains as in panels A and B and lysed with Laemmli buffer. Hyperphosphorylation of tau was analyzed by western blots, and relative levels of tau phosphorylation at individual sites are presented in gray and white matter from each case. Data were analyzed with two-way ANOVA followed by Sidak’s multiple comparisons. *p < 0.05.
The study above showed that RIPA-insoluble ag-gregated tau was hyperphosphorylated at multiple sites. To investigate the effect of AD brain extract treatment on tau phosphorylation in cultured cells, we treated HEK-293FT/HA-tau151–391 cells with the above crude extracts and lysed cells in Laemmli sample buffer. We found that tau phosphorylation in the cell lysates was not significantly changed in tau phosphorylation at the sites studied, including Thr181, Ser199, Ser202/Thr205, Thr212, Thr217, and Thr231 (Fig. 5E), suggesting pathogenic tau in AD brain may seed aggregation of hyperphosphorylated tau in the cultured cells.
DISCUSSION
Propagation of tau pathology following synaptic connected brain regions underlies the progression of AD. The present study showed a marked accumulation of hyperphosphorylated tau in AD gray matter. Extracts of gray and white matter from AD brain, but not control brain, induced tau151–391 aggregation in cultured cells, whereas AD gray matter extract displayed more potent seeding activity than the white matter. The seeding activity was correlated positi-vely with the levels of hyperphosphorylation of tau and SDS- and β-ME–resistant HMW-tau. AD brain extract induced the aggregation of hyperphosphorylated tau in cultured cells. These findings provide direct evidence of seeding-competent tau in both gray and white matter of AD brain and thus, a molecular basis of tau spread through axons between brain regions with tau pathology in AD.
In the present study, we compared the level of soluble tau in the frontal gray matter, which represents the somatodendritic compartment of the neuron, with that in the frontal white matter (the axonal compartment) by using semi-quantitative immuno-dot blots probed with a mixture of two pan-tau antibodies, R134d and 92e, which we have extensively characterized previously [29]. We found that when compared to the white matter, the corresponding gray matter has a similar level of total tau in normal aged human brains, which is consistent with our previous study [27]. As a microtubule-associated protein functioning primarily to stimulate microtubule assembly and to help maintain their structure, tau was initially thought to be an axonal protein [36] and has thus long been used as a marker of axons [37, 38], despite later recognition of its distribution in dendrites and the neuronal perikaryon [39, 40]. These early studies on the compartmental distribution of tau relied on immunohistochemistry, in which visibility of tau appeared to be affected by the way of tissue fixation [40]. To overcome this drawback, Khatoon et al. [27] employed radioimmunoslot blots to compare the tau level in frontal gray matter to that in frontal white matter in postmortem human brains with the Tau-1 antibody, and detected no significant difference in the levels of total tau between gray matter and white matter; the level of total tau was detected by Tau-1 after pre-treatment of blots with alkaline phosphatase. These findings together suggest that the somatodendritic compartment of the mature neuron has a similar level of total tau as the axonal compartment, at least in normal aged human brains.
The function of tau in stimulating microtubule assembly and stabilizing the microtubule network is inhibited by hyperphosphorylation [1, 41]. In AD brain, tau is hyperphosphorylated at multiple sites, and hyperphosphorylation leads to tau dissociation from microtubules and accumulation in the cell bodies [1]. Our previous study showed that the level of normal tau (tau unphosphorylated at Ser198/199/202) was reduced 40%, but abnormally phosphorylated tau at Ser198/199/202 is increased in 100,000×g supernatant from both gray and white matters in AD by subtracting normal tau from total tau determined by Tau-1 antibody with and without dephosphorylation [27]. Here, we found hyperphosphorylation and accumulation of tau in 10,000 x g supernatant in AD gray matter, whereas no significant elevation in AD white matter by using western blots developed with site-specific and phosphorylation-dependent tau antibodies, which is consistent with findings from previous study [42].
Oligomeric tau from AD brain (AD O-tau) seeds tau aggregation in vitro and in vivo [25, 29]. In addition to AD O-tau, monomeric tau with a specific conformation also seeded tau aggregation. The 10,000×g or 23,000×g extract contains oligomeric and monomeric tau, which were used to analyze tau seeding activity [34, 44]. Here we found that extract from gray matter or white matter of AD brain seeded tau aggregation, but much more tau aggregation was seeded by AD gray matter extract. No or very little tau aggregation was seeded by the extract from control gray or white matter. Thus, AD gray and white matter contains seeding-competent oligomeric and monomeric tau, but it predominantly presents in gray matter, where NFTs are formed.
Of particular note, we found here that gray matter exhibited a markedly higher level of soluble tau, a higher level of hyperphosphorylated tau, and higher seeding activity than white matter, even adjusted to the same level of total tau. This is consistent with a recent study by DeVos et al. [42], which reported a higher level of tau seeding in frontal gray than wh-ite matter by using the Forster resonance energy transfer (FRET)–based tau biosensors [45]. Interestingly, a more recent study by Thierry et al. [46] demonstrated that the seeding-competent tau species in axons in the pillar of fornix appear to be hyperphosphorylated and aggregated, but non-fibrillary, as no paired helical filaments were observed in the axons by immuno-electron microscopy. It was also reported that polymerization inhibits the seeding competency of aggregated tau [47], which may in turn enable the integration of NFT-burdened neurons in functioning cortical circuits in the mouse brain [48]. Collectively these findings strongly lend support to the notion that hyperphosphorylated oligomeric tau accounts for the seeding-competent tau species in AD brain, which was first shown in vitro by our group in 1996 [12] and confirmed by studies from our group and others recently [25, 49–52].
In addition to AD brain extracts, we found that some control brain extracts were also able to seed tau aggregation, even if slightly, in cultured cells, sugg-esting that some control aged brains may also have a trace amount of seeding-competent tau that can, but weakly, induce tau aggregation. In control brains, there is no tau pathology in the frontal lobe, which supports the hypothesis that seeding-competent tau may precede tau pathology [43].
Here we found that tau is hyperphosphorylated in AD gray and white matter, but more so in gray matter than in white matter. HMW-tau species, which are predominantly present in 3,000×g and 10,000×g brain extracts from tau-transgenic rTg4510 mice (expressing P301L tau) [53] and human sporadic AD cases, are efficient in uptake by neurons and in seeding tau aggregation [34]. HMW-tau is taken up, axonally transported, and passed on to synaptically connected neurons [34]. Phosphorylation affects HMW-tau uptake by neuron and determines the spread of AD O-tau seeded tau pathology [25, 34]. We also found that SDS- and β-ME–resistant hyperphosphorylated HMW-tau is present in both AD gray and white matter but is abundant in the gray matter. Levels of hyperphosphorylation of tau and hyperphosphorylated HMW-tau were positively correlated with seeding activity of the extracts, supporting the critical role of hyperphosphorylation and HMW-tau in the propagation of tau pathology.
In addition to the seeding-competent tau in AD gray and whiter matter, RIPA-insoluble aggregated tau templated by extracts of AD gray and whiter matter was hyperphosphorylated at multiple sites, and tau in RIPA-soluble fraction was similarly hyperphosphorylated at these sites. Interestingly, AD brain extracts did not alter the phosphorylation of tau in total cell lysates in parallel experiments. Thus, we conclude that hyperphosphorylated tau may be easier to be recruited by seeding-competent tau from AD extracts, and/or aggregated tau may be resistant to phosphatase [30].
Tau RD P301S biosensor assay was widely used to assess tau seeding activity in biological samples [54]. HEK-293 biosensor cells express a tau repeat domain (RD) sequence containing the P301S mutation tagged to either cyan fluorescent protein (CFP) or yellow fluorescent protein (YFP). At baseline, this tau protein is maintained in a soluble, monomeric form and is FRET negative. Upon tau aggregation introduced by adding tau seeds to the biosensor cells, a FRET response can be sensitively and quantitatively measured by using flow cytometry [45]. In the present study, we expressed tau151–391, truncated tau of the longest isoform tau441, tagged with HA at the N-terminus without disease-related mutation in HEK-293FT cells. Sedimentation of cell lysate in RIPA buffer yielded aggregated tau introduced by tau seeds, which was quantified by immuno-blots. In addition, small HA tag may disturb less tau aggregation. In summary, AD gray matter and white matter contain seeding-competent tau, which recruits and templates aggregation of hyperphosphorylated tau. Hyperphosphorylated and HMW-tau are associated with the seeding activity. These findings provide direct evidence that both gray matter (cell bodies and dendrites) and white matter (axons) can participate in the propagation of tau pathology.
Footnotes
ACKNOWLEDGMENTS
We are thankful to Maureen Marlow for copyediting the manuscript. This work was supported in part by a grant from the U.S. Alzheimer’s Association (DSAD-15-363172) and by funds from the New York State Office for People With Developmental Disabilities and Nantong University.