
Abstract
Background:
While extensive research on the brain has failed to identify effective therapies, using probiotics to target the gut microbiome has shown therapeutic potential in Alzheimer’s disease (AD). Genetically modified probiotics (GMP) are a promising strategy to deliver key therapeutic peptides with high efficacy and tissue specificity. Angiotensin (Ang)-(1-7) levels inversely correlate to AD severity, but its administration is challenging. Our group has successfully established a GMP-based method of Ang-(1-7) delivery.
Objective:
Since Drosophila represents an excellent model to study the effect of probiotics on complex disorders in a high throughput manner, we tested whether oral supplementation with Lactobacillus paracasei releasing Ang-(1-7) (LP-A) delays memory loss in a Drosophila AD model.
Methods:
Flies overexpressing the human amyloid-β protein precursor and its β-site cleaving enzyme in neurons were randomized to receive four 24-h doses of Lactobacillus paracasei alone (LP), LP-A or sucrose over 14 days. Memory was assessed via an aversive phototaxic suppression assay.
Results:
Optimal dilution,1:2, was determined based on palatability. LP-A improved memory in trained AD males but worsened cognition in AD females. LP-supplementation experiments confirmed that Ang-(1-7) conferred additional cognitive benefits in males and was responsible for the deleterious cognitive effects in females. Sex-specific differences in the levels of angiotensin peptides and differential activation of the kynurenine pathway of tryptophan metabolism in response to supplementation may underlie this male-only therapeutic response.
Conclusion:
In summary, LP-A ameliorated the memory deficits of a Drosophila AD model, but effects were sex-specific. Dosage optimization may be required to address this differential response.
INTRODUCTION
Alzheimer’s disease (AD) is the most common neurodegenerative disorder affecting close to 6 million adults aged 65 years or older in the US [1]. The pathophysiology behind AD is still under investigation, but hallmark signs include the formation of amyloid-β (Aβ) plaque deposits and tau-protein rich neurofibrillary tangles in the brain leading to neuronal death and loss of synaptic connections [2]. These pathological changes are suggested to set the basis for a progressive loss of memory; however, Aβ- and tau-targeting trials have not shown to reverse outcomes in AD patients [3, 4]. Indeed, most of the research has focused on the brain as the target organ but has failed to identify efficacious treatments which suggests the need to move away from a brain-centric approach for answers.
Recently, the role of the gut has been identified as an integral part of neural health and cognitive function [5–9]. The interaction between the gut and brain has been termed the “gut-brain axis” and communication between the gut and brain occur bidirectionally through a variety of mechanisms including production and expression of neurotransmitters, bacterial metabolites, immune regulation, and enteric sensory afferents [10–12]. Furthermore, aging is associated with disruptions in gut health with changes in gut microbial composition and populations [8, 13–15]. Therefore, understanding the relationship between the changing gut with age and the interactions of the gut-brain axis provide an important opportunity for targeting interventions for neurodegenerative disorders, including AD, in later life.
Angiotensin (1-7) [Ang-(1-7)] is a vasoprotective peptide in the counter-regulatory arm of the renin-angiotensin system (RAS) and is known to be abundant in the healthy gut [16]. Ang-(1-7) is formed as a result of the interaction of angiotensin-converting enzyme 2 (ACE2) and angiotensin I. This RAS axis stimulates several beneficial physiological effects including vasodilation and reduced inflammation [17]. More recent evidence suggests a benefit of Ang-(1-7) on AD pathobiology [5, 19]. Clinical and rodent studies suggest that the severity of AD is inversely correlated with circulating levels of Ang-(1-7) and ACE2 [18–20]. Unfortunately, a short half-life in plasma challenges the therapeutic potential of Ang-(1-7) [21]. Probiotics have been suggested as a safe and efficacious delivery method for therapeutic compounds, with a higher degree of site specificity, via genetically modified probiotics (GMPs) [22, 23]. They are one potential intervention to target the gut and thus, the gut-brain axis. Probiotics are live microorganisms which when properly administered can provide health benefits (i.e., antimicrobial activity, improved intestinal barrier function, immunomodulation) to the host [24, 25].
Here, we used a Lactobacillus paracasei-based GMP releasing the bioactive peptide Ang-(1-7) [LP-A]. Oral, but not parenteral, delivery of this peptide via LP-A has proved bioavailable and successful in inducing beneficial changes in the gut microbiome and eliciting circulating as well as brain-specific neuroprotective responses in older rats [5]. The metabolism of the essential amino acid tryptophan via the neuro-regulatory kynurenine pathway (KP) represents a critical link between the role of the Ang(1-7) axis in the gut to CNS function [26]. Recent data from human AD studies suggest that probiotics decrease systemic inflammation through the modulation of the KP of tryptophan metabolism [27]. This pathway establishes a key link between gut dysbiosis and neurodegenerative disorders and has shown to be differentially activated in conditions like AD [28, 29]. This pathway generates the metabolite 3-hydroxykynurenine (3-HK) and kynurenic acid (KA), two tryptophan-derived metabolites with well-described neurotoxic and neuroprotective roles in AD, respectively [30, 31]
To address whether an Ang(1-7)-releasing GMP confers neuroprotective effects in AD we used the common fruit fly Drosophila melanogaster. Drosophila represents an ideal model for this purpose due to its ability to model complex disorders, high throughput screening capabilities, low cost, and fast reproduction. Like vertebrates, enzymes regulating RAS, are present in Drosophila, including a fly angiotensin-converting enzyme (ACE), AnCE, which is inhibited by the same drugs that inhibit ACE in humans [32, 33]. The KP is also highly conserved in flies and like humans, this pathway has shown to modulate neuronal viability in various fly models of neurodegeneration including AD [34]. Additionally, the fly microbiome has proved amenable to intervention with key roles modulating neurodegeneration and aging [35]. Indeed, probiotic interventions have shown to simultaneously modulate age-associated elevations in oxidative stress and systemic inflammation, regulate metabolic homeostasis and enhance longevity in wild-type flies.
The present study aimed to determine whether oral treatment with LP-A delays the loss of memory function in a D. melanogaster model of AD. We report that the LP-A proved successful in postponing the memory decline in AD flies, but the effect was sex-specific. We used this model to further identify molecular targets mediating the benefits of LP-A on disease progression.
MATERIALS AND METHODS
Drosophila strains and rearing conditions
A Drosophila line (#33798) expressing two transgenes: the human amyloid-β protein precursor (hA βPP) and the human β-site APP-cleaving enzyme (hBACE) was acquired from the Bloomington Drosophila Stock Center (BSC). The transgenes were neuronally targeted using an elav-GAL4 driver (BSC#8760). This model expressing two human transgenes has shown to resemble the human phenotype [36, 37]. Co-expression of hA βPP and hBACE in this model favors the phenotypic expression of AD-associated traits [36, 37] and allows for the processing of hA βPP resulting in plaque formation [38]. Bloomington stock w1118 (BSC#5905) was used to generate driver-alone cohorts as the control group. All fly stocks and crosses were kept at a constant temperature of 25°C in a 12-h light:dark cycle at a 60-75% relative humidity, and fed a molasses-based food. To perform the experiments described below, newly eclosed elav-gal4/+(control) and hA βPP , hBACE/+; elav-gal4/+(AD) male and female virgins were collected in vials and raised at a constant temperature of 29°C in a 12-h light:dark cycle at a 60–75% relative humidity, and fed a molasses-based food. Flies were transferred to a new vial every 5–7 days to provide a fresh source of food.
Probiotic formulation and administration
The GMP releasing Ang-(1-7) was constructed using the plasmid pTRKH3-ldhGFP (Addgene, plasmid #27170) as the backbone of the expression vector for the angiotensin peptide as previously described [5, 40]. The Ang-(1-7) peptide was expressed as a secreted fusion protein with the chorea toxin binding protein subunit B (CTB), separated by a furin cleavage site which facilitates its transmucosal transport into circulation, followed by tissue uptake by GM1 receptor-mediated endocytosis. The resulting plasmids were electroporated into Lactobacillus paracasei (LP) as described by Welker et al. [41]. LP releasing Ang-(1-7) (LP-A) was cultured in MRS (deMan Rogosa Sharpe) broth (BD Difco, Houston TX, USA) media supplemented with 5μg/ml erythromycin at 37°C for 18 h. For administration, the bacteria were harvested by centrifugation at 5,000×g for 20 min and resuspended at a concentration of 1.2×1011 CFU/μl.
LP-A preparation was diluted in a 5% sucrose solution at 1:1 or 1:2 dilution. Experimental flies received the diluted preparation or sucrose alone starting on day 2 after eclosion following a 4-h fasting period. Probiotic preparation was administered through a capillary system resembling the Capillary Feeding (CAFÉ) assay [42] twice a week throughout the duration of the experiment. Two capillary tubes (10μl/capillary) containing the probiotic solution or sucrose alone were placed in each vial and replaced with new capillaries every 8 h. Flies were incubated under these conditions for 24 h after which they were transferred to fresh molasses-based food until the next dose. The number of flies in each vial and the amount of solution consumed were recorded per genotype, sex, and treatment group at each dose. A negative control vial for each treatment condition was included containing similar number of capillary tubes but no flies to account for evaporation under incubation conditions. Similar dilution and feeding conditions were used for cohorts of flies fed wild-type probiotic (LP).
Learning and memory assay
After two weeks of probiotic administration, control and AD male and female flies underwent an aversive phototaxic suppression assay. Flies, naturally drawn to light, were trained to prefer dark by pairing light to an aversive odor, 4-methylcyclo-hexanol (MCH). Within 48 h of the last probiotic administration, flies were quickly transferred to an opaque training chamber with an open and perforated section to control for light and odor exposure. The entire duration of the assay was conducted under red lighting as the wavelength is beyond the flies’ spectrum of vision. Temperature (∼25°C) and humidity (≥60%) were moderated for constancy. Replicates of the same cohort were run at the same time of the day. Sensory deficits were not detected in AD flies during preliminary studies (data not shown). After transferring to the training chamber, flies were acclimated for 2 min before being exposed to 1 min of light paired with MCH followed by 10 min of darkness with no odor for 10 cycles. An air pump was used to pull either the odor or air during each cycle.
Once training was completed, flies were immediately moved to a T-Maze followed by 2 min of acclimation. The T-maze then descended the flies to the decision tube, where they were given 2 min to choose between a light or dark chamber (Fig. 1). After the time was up, the T-maze was raised to prevent further decision making and the flies in each tube were then counted under carbon dioxide. This assay was completed in groups of at least 30 flies and at least four times for each sex, genotype, and treatment group. A separate cohort of untrained control and AD flies raised under control conditions were loaded in the T-maze and given the option to choose between dark and light chambers to confirm flies’ natural attraction to light. The proportion of flies choosing each chamber over the total number of flies in the experimental cohort was recorded.

Flies, naturally attracted to light, were trained to prefer dark by pairing light to an aversive stimulus. A) Once trained, flies were immediately loaded into a T-Maze followed by 2 min of acclimation. B) The T-maze then descended the flies to the decision tube for testing where they were given 2 min to choose between a light or dark chamber.
Measurement of tryptophan metabolites
Heads from 14-day-old flies were dissected and immediately frozen in liquid nitrogen and kept at –80°C until processing. We collected at least four independent replicates (30 heads/sample) per geno-type, sex, and treatment group. Fly heads were homogenized by sonication in 300μL of ultrapure water containing 0.01% BHT (butylated hydroxyto-luene). Fly homogenates and calibration standards were prepared in ultrapure water containing 0.01% BHT (100μL) with the appropriate amount of tryptophan (TRYP), kynurenic acid (KYNA), kynurenine (KYN), and 3-hydroxykynurenine (3-HK) to achieve concentrations ranging from 0.5–1,000 ng/mL homogenate. Additional QCs were prepared in control homogenate to ensure accuracy of extraction from the sample matrix. Standards, QCs, and samples were treated with deuterated internal standards (1μg/mL TRYP-d5, KYNA-d5, KYN-d4, and 3-HK 13C215N) and 10μl of 20% perchloric acid for protein precipitation. After centrifugation for 5 min at 21000×g, the supernatant was transferred to autosampler vials and analyzed in positive ion mode by LC/MS/MS consisting of a Shimadzu system (Columbia MD) equipped with LC20-AD dual HLPC pumps, an SIL20-AC HT autosampler, and a DGU-20A2 in-line degasser.
Mass calibration, data acquisition, and quantitation were performed using Applied Biosystem Analyst 1.6.2 software (Applied Biosystems, Foster City, CA). We used a Phenomenex Luna Omega Polar C18, 100×2 mm 5μm particle column to separate the tryptophan metabolites and the internal standards from the homogenate matrix. The mobile phase was delivered at a flow rate of 400μL/min using a gradient elution profile consisting of DI water with 0.1% formic acid (A) and acetonitrile with 0.1% formic acid (B). A gradient elution profile was used in which mobile phase B was held at 3% for 2.5 min, then increased to 90% over 2.5 min, held at 90% for 1 min, returned to 3%, and equilibrated for 5 min. The analytes were detected using multiple reaction monitoring for the following transitions: TRYP (m/z 205.0⟶188.0), KYNA (m/z 190.1⟶144.0), KYN (m/z 209.2⟶192.0), 3-HK (m/z 225.0⟶110.0). The internal standard transitions were as follows: TRYP-d5 (m/z 210.0⟶192.0), KYNA -d5 (m/z 195.1⟶149.0), KYN-d4 (m/z 213.0⟶196.0), 3-HK 13C215N (m/z 228.0⟶110.0).
Statistical analysis
Data were analyzed using two-way analysis of variance models with genotype (control or AD), treatment (untrained or trained/trained plus LP or LP-A for memory assays; sucrose, LP or LP-A for the other dependent variables), and genotype-by-treatment interaction included in the model. Repeated measures ANOVA was used to compare the level of intake per dose over the course of two weeks of intervention for each genotype and treatment groups in females and males. A two-way analysis of covariance (ANCOVA), with TRYP levels included as covariate in the model, was used to analyze variation for TRYP-derived metabolites. If data did not meet normality, a log10 transformation was applied. Post-hoc pairwise comparisons via a Tukey’s test were used to determine significant differences between groups. Statistical analyses were performed with SAS version 9.4 (SAS Institute, Cary, NC, USA). A significant level of 0.05 was used throughout the study.
RESULTS
Establishment of the optimal oral dose of LP-A in AD flies
Drosophila melanogaster is an excellent in vivo model to assess the effect of probiotics on complex disorders, like AD, in a high throughput manner [43]. Our group has recently shown that an orally delivered LP-A is bioavailable and can successfully target the gut-brain axis modifying the gut microbiome and displaying significant neuroprotective effects in 24-month-old male Fisher344/Brown Norway (F344/BN) rats [5]. Before testing the effect of LP-A in our fly model of AD, we needed to establish the optimal dose. The optimal dose was determined based on palatability in a cohort of AD male and female flies. As previously described in Materials and Methods, flies were fed LP-A at a ratio of 1:1 or 5% sucrose alone. Two-way ANOVA revealed no significant effect of sex (F1,26 = 2.23; p = 0.15), but a significant effect of LP-A supplementation (F1,26 = 61.30; p < 0.0001) on the average intake per dose per fly. As shown in Fig. 2A, AD cohorts supplemented with LP-A had a ∼33% lower average consumption per dose per fly compared to those flies fed 5% sucrose alone (0.52±0.02 versus 0.35±0.02μL/dose/fly).
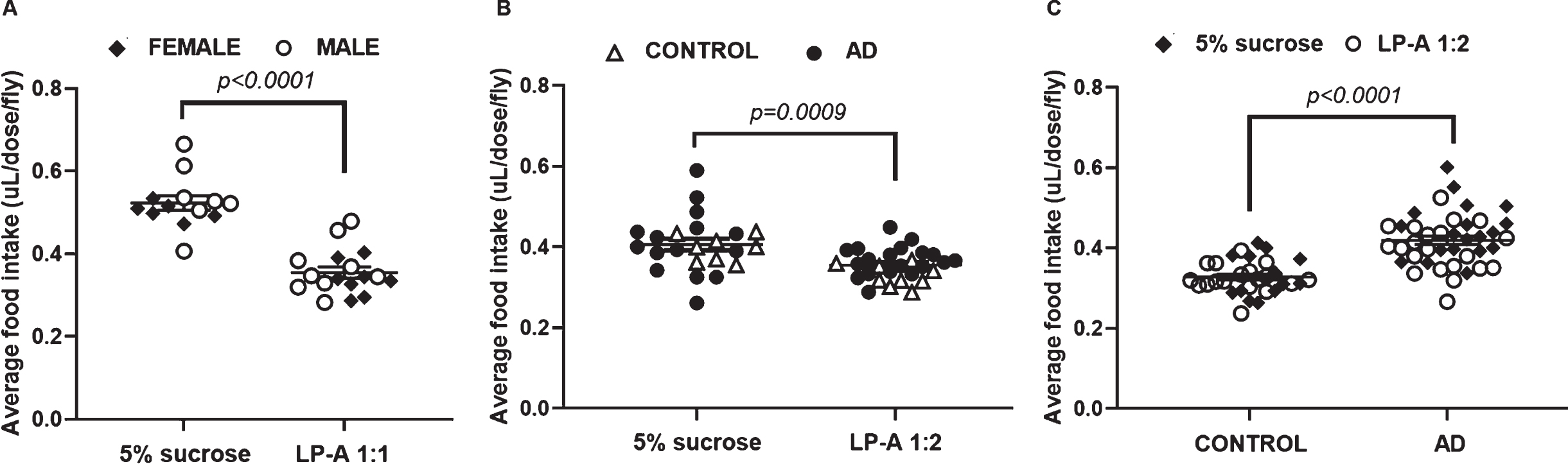
Food intake was differentially affected by genotype, sex, and LP-A supplementation. A) Regardless of sex, LP-A at 1:1 dilution dramatically decreased (∼30%) the average intake per dose per fly among AD cohorts. B) LP-A supplementation 1:2 significantly decreased food intake in females independent of genotype. C) Intake among males was unaffected by LP-A at 1:2 dilution. Regardless of supplementation, AD males ate significantly more than control males. Error bars represent S.E.M. p-value obtained from Tukey-Kramer post hoc tests for multiple comparisons.
Since our findings revealed that LP-A at a 1:1 dilution was considerably less palatable than sucrose alone which could impact the overall outcome of the experiment, we proceeded to test a 1:2 dilution instead. Two-way ANOVA showed a significant effect of sex (F1,70 = 4.49; p = 0.04) and LP-A supplementation (F1,70 = 8.84; p = 0.004) on the average intake per dose per fly. While AD males consumed significantly more than AD females (0.42±0.01 versus 0.39±0.01μL/dose/fly), LP-A supplemented AD flies ate significantly less than unsupplemented AD cohorts. However, the intake difference induced by LP-A at 1:2 dilution was less striking than the one described for the 1:1 dilution. Intake was ∼12% lower in LP-A supplemented flies compared to their sucrose-fed counterparts (0.38±0.01 versus 0.43±0.01μL/dose/fly). Considering these findings, we chose 1:2 as the optimal dilution based on palatability for the remaining experiments.
Food intake is affected by LP-A supplementation in a sex-specific manner
Quantification of food intake is of value to assess the impact of LP-A supplementation among control and AD cohorts. With 1:2 identified as the optimal dilution based on palatability, we proceeded to supplement control and AD female and male flies with LP-A over two weeks. Among females, there was no effect of genotype (F1,47 = 2.54; p = 0.12), but a significant effect of LP-A supplementation (F1,47 = 12.47; p = 0.0009) on the average food consumption per dose per fly. As shown in Fig. 2B, supplemented females ate significantly less than unsupplemented females independent of genotype. On the other hand, two-way ANOVA revealed no effect of LP-A, but a significant effect of genotype (F1,67 = 47.28; p < 0.0001) on the average intake per dose per fly among males. As reported in Fig. 2C, AD males had a considerably greater average consumption than their control counterparts independent of LP-A treatment.
Since LP-A was supplemented twice a week for two weeks, we analyzed the intake per dose over the four 24-h doses in female and male cohorts. Two-way repeated-measures ANOVA in females revealed that the intake significantly changed across the different doses (F3,141 = 14.96; p < 0.0001) with consumption at dose 1 (0.45±0.02μL/dose/fly) being significantly greater than consumption at dose 2 (0.36±0.01μL/dose/fly), 3 (0.34±0.01μL/dose/fly) and 4 (0.36±0.01μL/dose/fly). Additionally, this analysis showed a significant interaction between dose and treatment (F3,141 = 2.75; p = 0.05), indicating that the intake change across doses depended on LP-A supplementation. As shown in Fig. 3A, females fed LP-A, independent of genotype, had a significantly lower intake at dose 2 (0.32±0.02 versus 0.40±0.02μL/dose/fly) and dose 4 (0.31±0.02 versus 0.41±0.02μL/dose/fly) compared to their sucrose-fed counterparts. Two-way repeated-measures ANOVA in males showed a significant interaction between dose and genotype (F3,201 = 2.73; p = 0.04), indicating that intake was changing across doses, but this change depended on genotype. Overall, AD males consumed larger amounts of food at all doses compared to control males (Fig. 3B).
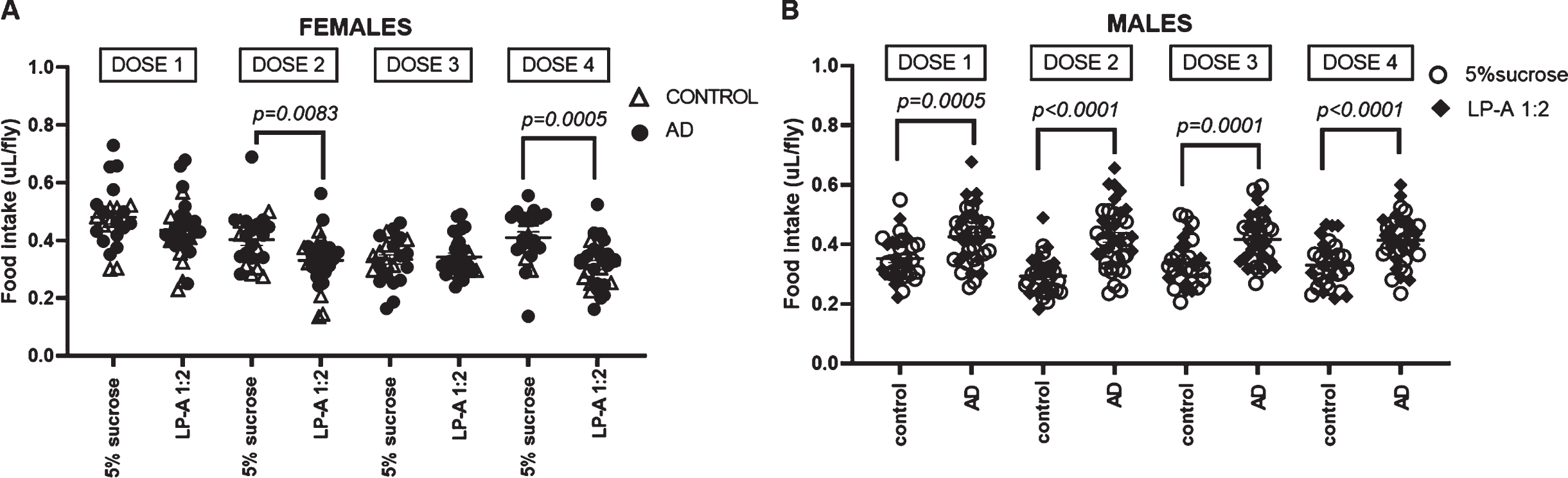
Food intake per dose was differentially affected by genotype and LP-A supplementation. A) LP-A supplementation significantly decreased intake at dose 2 and dose 4 among females, independent of genotype. B) Regardless of LP-A supplementation, AD males consumed larger amounts of food at all doses compared to control males. Error bars represent S.E.M. p-value obtained from Tukey-Kramer post hoc tests for multiple comparisons.
Overall, the outcomes we reported here are of interest since LP-A-mediated effects on cognition may be impacted by the intake per dose, particularly the last one before testing. Our findings reflect genotype- and sex-specific changes in feeding behavior which may mediate differential effects derived from LP-A supplementation.
LP-A supplementation ameliorated memory deficits in AD males but not in females
Since Ang-(1-7) has shown therapeutic potential in AD [18–20], we proceeded to test whether 2-week LP-A supplementation impacted learning and memory in AD flies. First, we compared the memory performance of untreated AD flies versus age-matched control flies via an aversive phototaxic suppression assay. A training regimen of 10 cycles of light exposure paired to an aversive odor, MCH, followed by darkness with no odor was used to modify flies’ natural preference to light. Task was learned if the fly reversed its ethological preference and chose the dark chamber. Two-way ANOVA among unsupplemented females revealed a significant effect of training (F1,12 = 18.62; p = 0.001) on preference towards dark, independent of genotype (F1,12 = 0.68; p = 0.43). As shown in Fig. 4A, both trained control and AD females displayed a significantly greater dark preference than their untrained counterparts. On the other hand, two-way ANOVA among unsupplemented males showed a significant effect of genotype (F1,14 = 5.23; p = 0.04), training (F1,14 = 6.77; p = 0.02) and a significant genotype-by-training interaction effect (F1,14 = 5.40; p = 0.04) on dark preference. As shown in Fig. 4D, while a significantly higher proportion of control males moved towards dark after training, we found no difference between untrained and trained AD males on dark preference. Thus, AD males but not females were memory impaired.
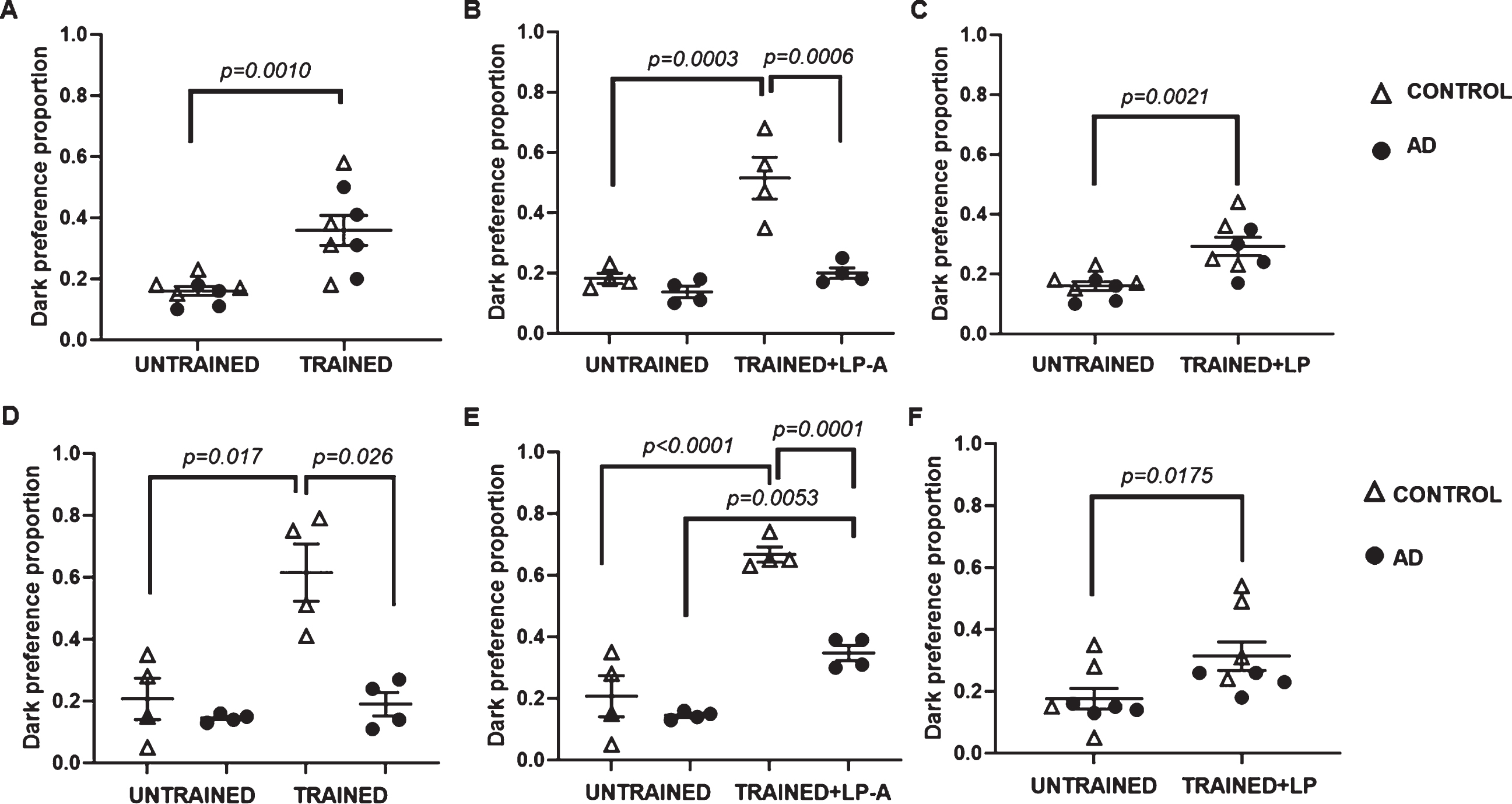
LP-A supplementation ameliorated memory deficits in a genotype and sex-specific manner. A) A higher proportion of females chose a dark chamber after training, independent of genotype. B) A significantly higher proportion of LP-supplemented females, independent of genotype, preferred dark after training compared to their untrained counterparts. C) LP-A supplemented control but not AD females prefer moving significantly more towards a dark chamber than a light chamber after training. Unlike AD females, AD males were memory impaired. D) Among unsupplemented cohorts, a higher proportion of control but not AD males chose a dark chamber after training. LP- (E), and particularly LP-A (F) supplemented males, significantly increased their dark preference after training, independent of genotype. Error bars represent S.E.M. p-value obtained from Tukey-Kramer post hoc tests for multiple comparisons.
Next, we proceeded to test the effect of LP-A supplementation on the memory performance of AD flies. There was a statistically significant effect of genotype (F1,12 = 27.26; p = 0.0002), training-plus-supplementation (F1,12 = 36.11; p < 0.0001) and a significant genotype-by-training-plus-supp-lementation interaction effect (F1,12 = 7.10; p = 0.02) on preference towards dark among females. While a considerably greater proportion of supplemented control females moved towards dark after training as shown in Fig. 4B, LP-A supplemented AD fe-males did not increase their dark preference. To confirm whether the effect was due to the Ang(1-7) peptide, we proceeded to test the effect of wild-type probiotic (LP) supplementation on the memory performance of AD females. Unlike LP-A supplemented AD females, two-way ANOVA showed a significant effect of training plus LP-supplementation (F1,12 = 15.31; p = 0.0021) on dark preference among females independent of genotype. Both LP-supplemented control and AD females moved significantly more towards dark after training compared to their untrained counterparts (Fig. 4C).
Findings were different among LP-A supplemen-ted males. There was a statistically significant effect of genotype (F1,12 = 29.83; p = 0.0001), training-plus-supplementation (F1,12 = 101.39; p < 0.0001) and a significant genotype-by-training-plus-supp-lementation interaction effect (F1,12 = 16.40; p =0.0016) on preference towards dark among males. Both LP-A treated control (untrained = 0.19±0.03 versus trained-plus-supplementation = 0.67±0.03; p < 0.0001) and AD males (untrained = 0.15±0.03 versus trained-plus-supplementation = 0.35±0.03; p = 0.0053) moved significantly more towards dark after training compared to their untrained counter-parts. However, while phototaxis was undistinguishable among untrained control and AD males, dark preference after training was significantly greater in supplemented controls compared to supplemented AD males (0.67±0.03 versus 0.35±0.03; p =0.0001).
To confirm whether the effect was due to the Ang(1-7) peptide, we proceeded to test the effect of LP supplementation on the memory performance of AD males. Two-way ANOVA showed a significant effect of training-plus-supplementation on dark preference among males for those supplemented with LP (F1,12 = 9.58; p = 0.0093). LP-supplemented males also moved significantly more towards dark after training than their untrained counterparts (Fig. 4E, F), but we detected no difference between genotypes. Of note, the proportion of flies preferring dark was greater with LP-A than with LP-supplementation (Fig. 4E, F), particularly for control males. In summary, our findings suggest that LP and LP-A supplementation ameliorated the memory deficits in our fly model of AD, but these benefits were not homogeneous between males and females.
Differential activation of the Kynurenine pathway (KP) of tryptophan metabolism in response to LP and LP-A supplementation
The KP of tryptophan metabolism represents a critical link between the role of the Ang-(1-7) axis in the gut to CNS function [44] and metabolites derived from this pathway have been associated to neurodegeneration and neuroprotection in humans [31, 46], in rodents [5], and fly models of neurodegeneration [34, 47]. Here, we proceeded to measure the levels of TRYP, KYN, KYNA, and 3-HK in dissected heads from 14-day-old flies supplemented or not with LP or LP-A. While there was no effect of genotype (females: F1,24 = 1.21; p = 0.28; males: F1,24 = 3.24; p = 0.08), supplementation had a significant effect on TRYP levels in both females (F2,24 = 20.95; p < 0.0001) and males (F2,24 = 67.46; p < 0.0001). As shown in Fig. 5A, TRYP was significantly higher in LP-supplemented females compared to their sucrose- and LP-A fed counterparts (p = 0.0247). LP- (p < 0.0001) and LP-A (p = 0.0247) supplemented males, on the other hand, displayed greater TRYP levels compared to untreated males, independent of genotype.
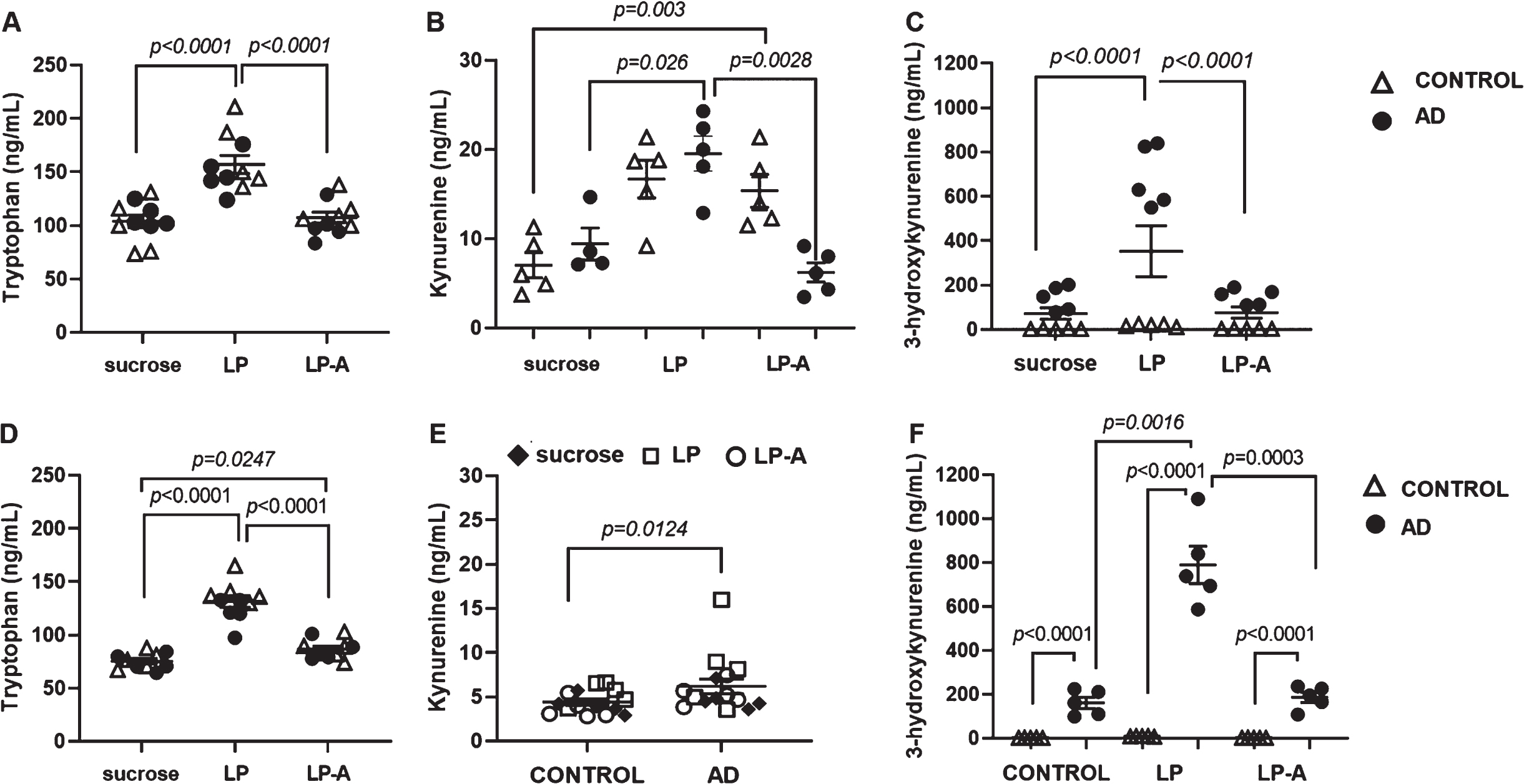
Activation of the kynurenine pathway of tryptophan metabolism is differentially affected by sex and supplementation. Tryptophan levels in heads of females (A) and males (D) were affected by supplementation. While only LP intervention increased tryptophan levels in females, LP and LP-A supplementation increased the levels of this metabolite in males. LP-A supplementation increased kynurenine levels in heads of control but not AD females (B). AD but not control males displayed significantly higher levels of kynurenine in heads (E). Concentrations of the neurotoxic 3-hydroxykynurenine in the heads of AD (C) female and (F) male flies were dramatically higher than their control counterparts. Supplementation with LP increased 3-HK levels significantly compared to sucrose- and LP-A fed females, independent of genotype (C). LP-supplementation increases 3-HK levels further in AD but not in control males (F). Error bars represent S.E.M. p-value obtained from Tukey-Kramer post hoc tests for multiple comparisons. Tryptophan levels were used as covariate in the analysis as indicated in Materials and Methods. Data points show unadjusted values.
Two-way ANOVA showed a significant effect of treatment (F2,22 = 6.67; p = 0.0054) and a significant genotype-by-treatment interaction (F2,22 = 7.41; p = 0.0035) on KYN levels among females. As shown in Fig. 5B, LP-A supplementation induced a significant 2-fold increase in KYN levels in heads of control females compared to their unsupplemented counterparts. A similar phenomenon occurs in response to LP- but not LP-A supplementation in AD females with a ∼2-fold increase in KYN levels compared to their sucrose- and LP-A fed counterparts. On the other hand, levels of KYN were significantly affected by genotype (F1,23 = 7.37; p = 0.0124) among males, with AD males displaying significantly higher KYN levels than control males independent of supplementation (Fig. 5E).
There was no effect of either genotype or supplementation on head levels of KYNA in female and male flies. However, there was a statistically significant effect of genotype (F1,23 = 1156.73; p < 0.0001) and treatment (F2,23 = 36.11; p < 0.0001) on the head levels of 3-HK among females. Regardless of supplementation, levels of this tryptophan-derived metabolite were ∼37-fold higher in AD females compared to controls. Interestingly, supplementation with LP increased 3-HK levels significantly, ∼4.5 fold, compared to sucrose- and LP-A fed females, independent of genotype (Fig. 5C).
Two-way ANOVA showed a significant effect of genotype (F1,23 = 2356.24; p < 0.0001) and treatment (F2,23 = 7.90; p = 0.0024) and a significant genotype by treatment interaction (F2,23 = 4.24; p = 0.0271) on 3-HK levels in males. Like females, AD males displayed strikingly higher levels of 3-HK, ∼78-fold, compared to controls independent of supplementation. However, unlike females, the effect of supplementation on 3-HK was not homogenous across genotypes. As shown in Fig. 5E, levels of 3-HK increased significantly in AD males, but not in control males, in response to LP-supplementation, ∼4.2-fold, compared to unsupplemented (p = 0.0016) and LP-A (p = 0.0003) supplemented AD cohorts. Overall, our findings suggest that AD flies, particularly males, produced considerably high levels of the neurotoxic 3-HK. While effects from LP-A supplementation were limited to increases in TRYP levels in our fly model of AD, LP-supplementation affected TRYP, KYN and 3-HK levels. However, this differential activation of the KP pathway of tryptophan metabolism in response to supplementation occurred in a genotype- and sex-specific manner.
DISCUSSION
This study highlights the potential of a fruit fly D. melanogaster model of AD as an easily manipulable and high throughput screening tool to assess the effect of a probiotic intervention on complex behavioral traits. Our findings here show that four doses of LP-A delivered orally via a GMP over two weeks had a positive impact on memory in a Drosophila model of AD. This effect on cognition, however, was sex specific.
While, in general probiotic interventions in flies have relied on the incorporation of the probiotic preparation in the Drosophila media [35, 48], herein, we used a modified version of the CAFÉ assay [42]. This novel administration method allowed us to provide a fresh source of probiotic solution every ∼8 h to ensure bacteria viability. Flies were also incubated at 30°C during LP-A feeding, which is within the optimum temperature range for growth for most lactic acid bacteria [49], like Lactobacilli. Additionally, this capillary-based administration allowed for food intake recording and the identification of differences among cohorts with high sensitivity, as previously suggested [50].
We detected a profound difference (i.e., > 30%) in intake between flies consuming the initial 1:1 dilution and those fed the sucrose alone. Although we can attribute these differences to palatability, an additional reason could be difficulty swallowing the LP-A preparation, a much thicker solution than sucrose alone. As shown previously, food dilution can modify intake volumes [50]. Flies may have a hard time ingesting thicker solution, thus lowering the amount they consume. This dramatic reduction in food consumption with LP-A at 1:1 dilution reduces the magnitude of the effect of the intervention which could impact experimental outcomes. A 1:2 dilution allowed for a more subtle intake difference between flies fed LP-A versus those fed sucrose alone (i.e., ∼12%), while the intervention dose was maximized.
Interestingly, food intake analysis with LP-A at 1:2 dilution revealed relevant sex-specific differences in feeding behavior. Regardless of genotype, supplemented females ate significantly less than their unsupplemented counterparts. AD males, however, proved to be unaffected by LP-A supplementation, eating considerably more than control males and AD females. Clinical studies suggest that the severity of dementia in AD increases in proportion to the increment in appetite [51]. It is clear from our learning and memory assays that AD males were further compromised than females, which could account for the striking increase in food intake reported among AD males. Like flies, male patients with AD generally display worse outcomes, are reported to die earlier [52, 53], and display an earlier onset of preclinical symptoms (i.e. mild cognitive impairment) compared to women [54]. Our findings also agree with earlier studies reporting a decreased response to satiety in male rodent models of AD [55]. Authors herein attributed these disturbances to a defective gut-brain signaling, which validates the therapeutic relevance of our approach.
Previous work highlights the role of the gut-brain axis as modifier of AD progression in Drosophila [56] and humans [57]. Recent literature attributed neuroprotective effects to Lactobacillus strains in fly models of AD [58, 59]. Similar findings have been reported in rodent models of AD [60] and in AD patients [61], where Lactobacillus-based preparations or multi-strain probiotic preparations including Lactobacillus, respectively, have shown to ameliorate memory deficits. However, the use of GMPs for neurodegenerative diseases has not been comprehensively studied even though this approach offers a means to deliver therapeutic peptides with greater efficacy, precision, and high degree of tissue specificity [22]. Our group has recently shown [5] that an orally delivered Lactobacillus-based GMP releasing Ang-(1-7) was able to target the gut-brain axis, modify the gut microbiome, and elicit neuroinflammatory protective effects in aged rats. Here, we confirmed that this intervention induced positive behavioral changes in our fly model of AD, but in a sex-specific manner.
Ours is the first study showing the benefits of Ang-(1-7) supplementation delivered via a GMP in a Drosophila model of AD. Recent studies in rodent models and humans suggest an inverse correlation between AD severity and the circulating levels of Ang-(1-7) [18–20]. These findings justify the need for supplementation. Parenteral administration of Ang-(1-7), however, poses challenges due to its short half-life in plasma [21] and has failed to impact the gut-brain axis and associated neuroinflammatory parameters in rodent models [5]. The GMP-based method we used here allows for oral administration and sustained peptide production with the added benefits of Lactobacillus colonization of the gut and changes of gut-brain parameters, as described previously [5, 59]. While increased circulating levels of Ang-(1-7) have been demonstrated following oral gavage with LP-A in rodents [39], limitations of detection sensitivity and tissue sample size as well as the short half-life of the peptide may account for our inability to measure Ang-(1-7) levels in tissues and whole flies.
The benefits from Ang-(1-7), however, were sex-specific with AD males, but not females, displaying improved memory after 2 weeks of LP-A supplementation. To confirm whether this improved memory was due to Ang-(1-7), we added a cohort of males fed wild-type LP and found that LP-supplementation improved memory after training for both control and AD males as previously reported in rodent models of AD [60] and AD patients [61]. However, the proportion of flies preferring dark was much larger among those supplemented with LP-A. These findings suggest a synergistic effect between the bacteria and the angiotensin peptide. Contrarily, memory in AD females was preserved and training alone was sufficient to modify behavior and increase dark preference among them, a behavior that worsened after LP-A but not LP-supplementation. This sex-specific detrimental effect of Ang-(1-7) is not surprising since differences in the levels of angiotensin peptides, including Ang-(1-7), have been reported between males and females [62, 63]. Parenteral administration of Ang-(1-7) has also shown sex-specific effects in rodents [64].
We anticipated that LP-A supplementation targeted the brain via modulation of the KP of tryptophan metabolism as previously shown in LP-A supplemented rodents [5]. Our findings revealed that only LP-A supplementation increased head TRYP levels in supplemented males but not in females. Like male flies, Ang(1-7) has shown to increase tryptophan absorption in the gut of male rodents [26]. TRYP levels have shown to be negatively correlated to cognitive deficits in dementia [65] and non-dementia patients [66] which could account for the improved memory status in supplemented AD males. Of note, TRYP levels were overall higher in unsupplemented females than males which could account for the preserved memory in AD females. Interestingly, LP but not LP-A supplementation significantly increased TRYP levels in females. LP has a similar effect in males which may account for the cognitive benefits in males from LP-supplementation. This is not surprising since gut bacteria can affect tryptophan metabolism [67] and like flies, Lactobacillus-based probiotic administration has previously shown to augment circulating TRYP levels in humans [68, 69]. Additionally, LP-A supplementation increased KYN levels in control but not in AD females. Head KYN levels in males, however, were unresponsive to LP-A intervention.
While we found no differences in the head levels of the neuroprotective KYNA metabolite, ours is the first study reporting a dramatic increase in head 3-HK levels in a Drosophila model of AD. This is an exciting finding considering the current need for disease biomarkers in AD. Production of 3-HK has been associated to oxidative stress and neuronal death [45, 46]. Like flies, 3-HK has been reported to be strikingly increased in the serum of AD patients [30]. Of note, while levels of 3-HK were high in both AD males and females, the levels were a lot higher in males supporting the idea of a worse prognosis in this sex cohort. Our findings also showed that LP, but not LP-A, supplementation strikingly increased the levels of the neurotoxic 3-HK in AD males. This may explain why LP-A supplemented AD males performed better in the memory assay than LP-supplemented cohorts and raise the intriguing possibility that the accumulation of the neurotoxic 3-HK may deter the cognitive benefits derived from LP supplementation in the long term.
In summary, two-week oral administration of LP-A ameliorated the memory deficits of a Drosophila model of AD. The effect, however, was only in males while memory worsened in females in response to LP-A supplementation. The inclusion of LP-supplemented cohorts confirmed that this worsening could not be attributed to the bacteria but to Ang (1-7). Sex-specific differences in the levels of angiotensin peptides, including Ang-(1-7), and the activation of the KP of tryptophan metabolism in response to LP-A may account for these differences in therapeutic response between males and females. Dosage optimization may be required to address this sex-specific response. The fly represents an attractive model to carry on dose-optimization trials. A fly-based high throughput approach also offers an excellent opportunity to refine host/microbe interactions and identify bacterial constructs that may integrate more efficaciously into the gut of females versus males conferring sex-specific advantages. Overall, these findings can be translatable to higher-order organisms such as rodents to further our understanding of the therapeutic potential of Ang(1-7)-releasing GMP on AD-associated outcomes.
Footnotes
ACKNOWLEDGMENTS
We are grateful to Kenneth H. Moberg and Alfonso Martin-Peňa for helpful discussion and assistance on technical aspects of the aversive phototaxic suppression assays.
This work was funded by a supplement grant from the National Institute of Health/National Institute of Aging (3RO1AG054538-03S1) to TWB and CSC. This project was also supported by a pilot grant from the University of Alabama Nathan Shock Center (P30AG50886) to PJL and QL, and by the McWhorter School of Pharmacy Summer Research Grant to CAS and HS.