
Abstract
Alzheimer’s disease (AD) has become a major health problem among the elderly population. Some evidence suggests that metabolic disturbance possibly plays a role in the pathophysiology of AD. Currently, the study of metabolomics has been used to explore changes in multiple metabolites in several diseases, including AD. Thus, the metabolomics research in AD might provide some information regarding metabolic dysregulations, and their possible associated pathophysiology. This review summarizes the information discovered regarding the metabolites in the brain and the blood from the metabolomics research of AD from both animal and clinical studies. Additionally, the correlation between the changes in metabolites and outcomes, such as pathological findings in the brain and cognitive impairment are discussed. We also deliberate on the findings of cohort studies, demonstrating the alterations in metabolites before changes of cognitive function. All of these findings can be used to inform the potential identity of specific metabolites as possible biomarkers for AD.
INTRODUCTION
As the world population approaches an aging society, dementia is becoming a major health problem. More than forty-seven million people have been affected by dementia worldwide and this number is expected to increase [1]. Among the number of dementia cases, Alzheimer’s disease (AD) is associated with two-thirds of dementia cases [2]. AD is characterized by the deposition of extracellular amyloid-β protein (Aβ) as neuritic plaques and the intracellular accumulation of phosphorylated tau as neurofibrillary tangles [3]. Neuronal loss and reactive microgliosis are additional pathologic changes in the AD brain [3]. The exact pathophysiology of AD is still elusive, but several risk factors, in addition to aging, also enhance the risk of AD, including genetic risk factors such as apolipoprotein E4 (APOE4), as well as type 2 diabetes, obesity, dyslipidemia, andmetabolic syndrome [2]. These AD risk factors also involve metabolic dysfunction, which causes alterations of various metabolites in the body. Therefore, studies in metabolic changes could reveal some underlying mechanisms related to metabolic dysregulation and could discover some novel biomarkers or early therapeutic targets for dementia AD in an aging population.
To study the metabolic changes in AD, many metabolomic studies in these conditions have been conducted. The studies of the metabolome or metabolomics have revealed the simultaneous disturbance of multiple metabolites which has led the investigators to identify and observe the ongoing occurrence of multiple processes during the progression of the disease. The short details of metabolomic studies have been reviewed elsewhere [4]. Briefly, the metabolites are extracted from the sample such as blood or organ tissues, and then undergo analyses via mass spectrometry or nuclear magnetic resonance spectroscopy (NMR). The separation and identification of the metabolites are based on the mass per charge ratio (m/z ratio). Due to the large amount of data accrued from metabolomic studies, many forms of statistical analyses have been utilized to understand those data. However, due to the diversity of metabolites, the findings have been dissimilar among the studies. The different models used in the animal studies and groups of diagnosed patients have also contributed to the inconsistency of the data. Thus, this present review aims to collect and summarize the findings from metabolomic studies in dementia and AD from both in vivo and clinical studies. The controversial findings will also be discussed.
The original articles published on the PubMed database before September 2020, selected for relevance, are included in this review. The MeSH terms ‘metabolomic’, ‘dementia’, and ‘aging’ were used to search for articles. Fifty-five original articles were screened and 22 of these articles were excluded as they were not related to metabolomics research or dementia and aging. The remaining 33 references which were selected on the basis of the relevance to the topic of this review were included. The article selection method is displayed in Fig. 1.

Article selection method.
ALTERATIONS IN METABOLITES FROM AD MODELS IN ANIMAL STUDIES
Phospholipids
Phospholipids are one of the major constituents of the cell membrane. The myelination of neurons in the central nervous system also plays an important role in signal transduction and this myelination also depends on normal lipid metabolism, including that of phospholipids [5]. Therefore, the alterations in phospholipids and the association with the development of AD have been a focus in several studies (Table 1) [6–11]. The increases in the brain sphingomyelins (SM), one of the phospholipids, have been shown in genetically modified AD mice, including AβPP/PS1 mice, and PBL4 mice at the age of 8 months [6, 7]. The PBL4 mice are created by the hbace1 knock-in process which makes these mice able to produce human Aβ protein precursor (AβPP) while AβPP/PS1 mice are double transgenic mice which produce both humanized AβPP and PSEN1. The Aβ plaques develop in the brain of AβPP/PS1 mice at six months of age whereas they do not in PBL4 mice, but rather produce Aβ oligomers instead. The models of mice with elevated AD risk factors, including a model of chronologically aged female mice and a model of human APOE4 knock-in mice, showed the increases in brain ceramide and SMs, respectively [8, 9]. A human APOE4 knock-in mice model is an animal model containing a humanized APOE4 allele as an AD risk factor whilst preserving locomotor activities and working memory. The other studies raised inconsistent findings regarding several glycerophospholipids, mainly phosphatidylcholine (PC). In one study, the PCs were found to be increased at 8 months of age in AβPP/PS1 mice [6]. In another study, young PBL4 mice had reduced the brain PC levels, while some subtypes of PC were elevated in old PBL4 mice [7]. These different metabolomic profiles may be associated with the different types of Aβ generated in each model. PBL4 mice exhibited the accumulation of Aβ*56 and Aβ hexamers in the brain and with sparse development of Aβ plaque [12], whereas AβPP/PS1 mice produced Aβ1–42 which led to Aβ plaque formation [13]. The levels of LysoPC were also found to be elevated in the brains of aged human APOE4 knock-in female mice [9]. These data suggested that the perturbations of phospholipid metabolisms in the brain might be associated with AD.
Changes in brain and blood metabolites in animal models with dementia
Aβ, amyloid β; APOE, apolipoprotein E; BCAAs, branched-chain amino acids; cAMP, cyclic adenosine monophosphate; CoA, coenzyme A; DHA, docosahexaenoic acid; EAA, essential amino acids; FIA-MS/MS, flow-injection - tandem mass spectrometry; GC-MS, gas chromatography-mass spectrometry; GC-MS/MS, gas chromatography-tandem mass spectrometry; hAPOE, human APOE; LC-MS, liquid chromatography-mass spectrometry; LC-MS/MS, liquid chromatography-tandem mass spectrometry; LysoPCs, Lysophosphatidylcholines; NA, not available; NAAG, N-acetyl-aspartyl-glutamate; NEAA, non-essential amino acids; PCs, phosphatidylcholine; PUFAs, polyunsaturated fatty acids; SAMP8, senescence-accelerated prone 8; SMs, sphingomyelin; TCA, tricarboxylic acid; UPLC-MS, ultraperformance liquid chromatography-mass spectrometer; UPLC-MS/MS, ultraperformance liquid chromatography-tandem mass spectrometer; WT, wild type.
Changes in phospholipid levels were also observed in plasma; however, these were not associated with the changes in the brain. In AβPP/PS1 mice, the level of PCs in plasma was decreased at the age of 8 months [6]. Additionally, the reductions of LysoPC in plasma were observed in the mice with hippocampal Aβ1-42 injections [10]. The acute Aβ1-42 injections into the hippocampus result in impaired learning and memory processes, and neurodegeneration which resembles AD. This model could demonstrate the direct effects of Aβ1-42 on the brain and cognitive function of the animal. Conversely, the PCs and LysoPCs were relatively higher in the male mice when compared to the female mice despite being of the same APOE genotype [9]. The lower PCs and LysoPCs in female mice might be associated with AD because the incidence of AD was slightly higher in females than in males [14]. Additionally, the mice with APOE2 replacement, which showed beneficial effects on cognitive function, had increases in the levels of phospholipid in the plasma [11]. These data suggested that the elevations of phospholipid in the plasma of male and APOE2 replacement mice might be inversely associated with AD. In the case of sphingolipid in the plasma, dihydrosphingosine was decreased in mice after hippocampal Aβ1-42 injections [10]. The associations of sphingolipid changes in plasma and AD need to be investigated further.
In summary, the phospholipid changes in the brain and plasma in animal models were associated with AD. The PCs tended to be elevated in the brain from AD models, while the circulating PCs may confer beneficial effects. Increased sphingolipid levels in the brain were also associated with AD. The dysregulation of phospholipids may be associated with Aβ formation and aggregation, impairment of synaptic activity, and neuronal cell death, resulting in the development of AD [15].
Fatty acids
The brain is a metabolically active organ, that utilizes glucose as a major source of energy [16]. Although fatty acid can be an alternative energy source for brain metabolism, there are still some disadvantages [17]. Firstly, to achieve the same level of energy production, more oxygen is required for fatty acid oxidation than glucose metabolism which may lead to tissue hypoxia of the brain [17]. Secondly, the metabolism of fatty acid through the β-oxidation process and the electron transport chain of mitochondria produced a large number of oxidative stress [17]. Finally, the rate of ATP generation from the fatty acids might be too slow and could not meet the usual metabolic demand of the brain since it depends on long and complex metabolism pathways [17]. Therefore, the disturbances in fatty acid metabolism, as indicated by altered levels of fatty acid and acylcarnitine, may be associated with AD (Table 1) [6, 18–20]. The brain of aged human APOE4 knock-in mice showed the decreased fatty acid levels indicating utilization of fatty acids as a fuel source by neurons during the neuronal hyperactivity stage, which was demonstrated by the upregulation of tricarboxylic acid (TCA) cycle metabolites including citrate, isocitrate, and malate [18]. However, in the brain of aged female mice without any AD pathology or APOE risk factors, the fatty acids were increased as a consequence of sphingomyelinase activity [8]. The alterations in metabolites in the TCA cycle were observed in SAMP8 mice [20]. SAMP8 mice are naturally occurring mice which exhibit phenotypes of accelerated aging and neuropathology such as microgliosis, neurodegeneration, and hippocampal Aβ without plaque formation. Higher levels of polyunsaturated fatty acids (PUFAs) were also found in the blood of SAMP8 mice [20]. In addition to fatty acid, the changes observed in the levels of acylcarnitine might reflect the abnormal lipid metabolism. In the female AβPP/PS1 mice, the acylcarnitine levels were altered in the brain indicating the dysregulation of lipid metabolism in the brain in cases of AD [6]. Furthermore, aged-SAMP8 mice and human APOE4 knock-in female mice showed increased acylcarnitine levels in their blood [9, 20]. Acylcarnitines were also found to be higher in female or aging mice in these studies [9, 11]. However, the carnitine level was increased in both male and female human APOE4 knock-in mice [9]. These findings suggested that the disturbances in the lipid metabolism in relation to increased acylcarnitine levels were associated with risk factors for AD. However, in the female AβPP/PS1 mice, the acylcarnitine C4:1 and C6:1 levels were reduced in the blood during the period of 8–18 months of age when compared to their wild-type counterpart at each age [6]. These different findings might be due to differing pathologies of AD in each study.
In conclusion, changes in fatty acid metabolism were found the brain and the blood of AD models in animal studies. The elevation in fatty acid levels and acylcarnitine levels potentially reflect the increase in the metabolic demand associated with AD or the risk factors of AD.
Amino acids
Amino acids play an important role in the function of the brain as the precursors of several neurotransmitters [21]. Some metabolomic research on AD has focused on the changes in these metabolites (Table 1) [6, 20]. Overall changes in amino acids were observed in the brain of human APOE4 knock-in mice. Male APOE4 mice had higher levels of glucogenic amino acid in the brain than the female mice which had a higher level of LysoPCs. This information suggested that the female mice, which are associated with a higher risk of dementia and AD, tended to exhibit a lipid metabolic profile [9]. Furthermore, the decreases in amino acids and biogenic amines in the blood were associated with the age in one animal metabolomic study [11]. These findings suggested that, overall, the increased levels of several amino acids were inversely associated with dementia and AD. The metabolomic study in AβPP/PS1 mice showed transient changes in several amino acids in the different ages both in the blood and brain. These indicated the different precipitating factors of AD including impairment of amino acid management by the liver, disturbance of neurotransmitter function, or increased oxidative stress [6]. Additionally, changes in amino acid levels in the plasma were observed in several studies, suggesting changes in both metabolisms and the oxidative state in the AD model [10, 20].
Polyamine is a biogenic amine derived from the amino acid arginine and is involved in diverse physiological functions [22]. The polyamines were increased in the model of AβPP/PS1 mice at the ageof 6 and 8 months, while the changes in the bloodwere only observed at around 10 months in the same model [6]. An increase in polyamine might promote the higher activity of the N-methyl-D-aspartate (NMDA) receptor, leading to neuronal excitotoxicity. However, in the PBL4 mice, the level of putrescine, one of the polyamines, was decreased [7]. The differences in the changes between these two studies might be explained by the differences in Aβ formation in different AD models.
Conclusively, the alterations in amino acids and biogenic amines such as polyamine could reflect several pathophysiologies of AD. The dynamic changes of the disease might lead to the fluctuation of the levels of these amino acids and polyamine metabolites which made the findings inconsistent among the studies.
METABOLIC CHANGES IN ANIMAL MODELS WITH AD FOLLOWING NEUROPROTECTIVE INTERVENTIONS
Several neuroprotective compounds were used in these models of dementia and AD to improve cognitive function and brain pathology. Thoseinterventions altered the level of the metabolites both in the brain and the blood (Table 2) [10, 19,20]. J147 is the neuroprotective compound derived from curcumin and provides beneficial effects on cognitive function [23]. J147 treatment altered themetabolites in SAMP8 mice as shown by decreases in acylcarnitine and PUFAs, and increases in aminoacids in the blood including the gamma-glutamyl amino acids, branched-chain amino acid, and dipeptides, which may be associated with the improvement of the metabolism and oxidative status [20]. Additionally, the upregulations of glutamate and alpha-ketoglutarate and a decrease in cAMP in the brain were also observed after the J147 treatment indicating metabolic improvement [20]. Fisetin, a bioactive flavonoid, also provides neuroprotective effects via multiple mechanisms [24]. Treatment with fisetin improved cognitive function and changed the metabolites in the brain and blood of old SAMP8 mice to be similar to metabolites those in the young SAMP8 mice. The differences in changed metabolites between Fisetin and J147 might be due to the different mechanisms of each compound [19, 20]. GRb1 and GRg1, the major active ingredients of ginseng were administrated in 12-week-old mice with hippocampal Aβ1-42 injection. These treatments showed neuroprotective effects and improved the metabolites in the plasma [10]. The high dosage of GRb1 and GRg1 decreased the level of phenylalanine but increased the levels of tryptophan and LysoPCs in plasma [10]. However high dosage GRg1, but not GRb1 increased the dihydrosphingosine level in plasma [10]. This inconsistency might be due to the different chemical structures of these two compounds.
Change in brain and blood metabolites in animal models with AD following neuroprotective interventions
Aβ, amyloid β; BCAAs, branched-chain amino acids; cAMP, cyclic adenosine monophosphate; GC-MS/MS, gas chromatography-tandem mass spectrometry; IP, intraperitoneal; LC-MS/MS, liquid chromatography-tandem mass spectrometry; LysoPCs, Lysophosphatidylcholines; PO, peroral; PUFAs, polyunsaturated fatty acids; UPLC-MS/MS, ultraperformance liquid chromatography-tandem mass spectrometer; SAMP8, senescence-accelerated Mice P8.
In summary, the interventions that improve cognitive function and brain pathology also change the level of several blood and brain metabolites of AD models. The inconsistent changes in metabolites between studies might be due to the different models of dementia and the different mechanisms of the interventions. A study using a single model with an intervention with a specific target should be further investigated.
CHANGES IN METABOLITES ASSOCIATED WITH AD: EVIDENCE FROM CLINICAL STUDIES
Some of the clinical studies used a metabolomic approach to compare the profiles of brain metabolites between normal subjects and AD patients (as shown in Table 3) [25–30]. The authors demonstrated that the changes in sphingolipids, glycerophospholipids, and fatty acids levels in the brain were associated with AD clinical status. Those changes were associated with the severity of AD, as indicated by changes in sequence from AD > asymptomatic AD > normal [25, 26]. A reduction in brain glucose metabolism, as indicated by decreases in the ratios of brain metabolites in glycolysis, was also observed in progressive AD [27]. Additionally, the disturbances of several amino acids and biogenic amines in the AD brain indicated that the transmethylated process was dysregulated in cases of AD [28]. The changes in the levels of sphingolipids, phospholipids, fatty acids, glucose, amino acids, and polyamines in these studies were associated with brain AD pathologies, measurable using the Consortium to Establish a Registry for Alzheimer’s Disease (CERAD) and Braak scores, which are used to represent the neuritic plaque burden and level of neurofibrillary tangle in the brain, respectively. Increases in SDMA and threonine levels were also associated with brain-related AD phenotypes [31]. In addition to the brain, an untargeted-metabolomic study of cerebrospinal fluid (CSF) also detected changes in the levels of metabolites in AD patients, when compared with non-AD controls [29]. Another study into CSF in dementia patients demonstrated a reduction in metabolites related to polyamine and tryptophan-kynurenine [30]. All of these findings from metabolomics research in the human brain revealed the dysregulation of the metabolites which may be related to cell membrane integrity, alteration in the source of energy, and disturbance in neurotransmitter metabolism. Furthermore, the impairment of the blood-brain barrier which occurs during the progression of AD might lead to the differing composition of the metabolites in the brain and CSF. The study of ratios of the metabolites between these two areas might provide useful information regarding blood-brain barrier function.
Changes in brain, CSF, and blood metabolites in patients with cognitive decline and associated brain pathologies
AD, Alzheimer’s disease; Aβ, amyloid β; CA, cholic acid; CDCA, chenodeoxycholic acid; CERAD, Consortium to Establish a Registry for Alzheimer’s Disease; CSF, cerebrospinal fluid; DCA, deoxycholic acid; DHA, docosahexaenoic acid; EPA, eicosapentaenoic acid; ESI-MS/MS, electrospray ionization - mass spectrometry; FIA-MS/MS, flow-injection - tandem mass spectrometry; GABA, gamma-Aminobutyric acid; GC-MS, gas chromatography - mass spectrometry; GC-MS/MS, gas chromatography - tandem mass spectrometry; GDCA, glycodeoxycholic acid; GLCA, glycolithocholic acid; GSH, thiol-reduced glutathione; HDL, high-density lipoprotein; HPLC-MS/MS, high-performance liquid chromatography - tandem mass spectrometry; IQR, interquartile range; LC-MS, liquid chromatography - mass spectrometry; LC-MS/MS, liquid chromatography - tandem mass spectrometry; MCI, mild cognitive impairment; NA, not available; MRI, Magnetic resonance imaging; NAA, N-acetylaspartate; NAB, neocortical Aβ burden; NAG, N-acetylglutamate; PCs, phosphatidylcholine; PE, phosphoethanolamine; PET, Positron emission tomography; SAM, S-adenosylmethionine; SDMA, symmetric dimethylarginine; SMs, sphingomyelin; SPARE-AD, Spatial Pattern of Abnormality for Recognition of Early Alzheimer’s disease; TC, total cholesterol; TDCA, taurodeoxycholic acid; UPLC-MS, ultraperformance liquid chromatography - mass spectrometer; UPLC-MS/MS, ultraperformance liquid chromatography - tandem mass spectrometer.
Most of the clinical metabolomics studies specific to AD were conducted using plasma or serum samples (Table 3) [32–42]. However, despite the convenience of accessing the specimens, plasma or serum may not directly reflect the changes in the brain. However, information from plasma and serum contains information regarding abnormality throughout the body, which might be indirectly associated with the development of dementia. The changes in lipid metabolites from plasma and serum are the main findings from those clinical studies (Table 3) [25, 44]. The levels of acylcarnitines and free carnitine in plasma were both reduced when the disease progressed from normal to mild cognitive impairment (MCI) or from MCI to AD [32]. In one study, acylcarnitines including C8, 10, and 12 were associated with a higher volume of gray matter in the brain [32]. Reductions in levels of acylcarnitines were also observed in another two studies, which compared the presence of metabolites in AD and MCI [33, 34]. These findings indicated that decreases in serum and plasma acylcarnitine were associated with clinical symptoms of AD. In the case of other lipids, one untargeted-metabolomic study found that the levels of ether phospholipid, PCs, SMs, and sterols were reduced in correlation with the increasing severity of AD [35]. In another study, SMs were lower and PC was higher in cases of early AD in comparison to MCI [33]. Conversely, several phospholipids were increased in AD or MCI, when compared to those of non-AD and non-MCI controls [33, 37]. The findings within each study, even though conducted specifically in association with AD, cannot be directly compared due to the different methodologies, variety of metabolites, and clinical symptoms. In the case of fatty acid and cholesterol levels, one study found that the docosahexaenoic acid (DHA) and cholesterol level in HDL were reduced in dementia and AD patients, while another study showed that the levels of fatty acids, including docosapentaenoic acid and 3,4-dihydroxybutanoic acid, were increased in dementia patients [38, 39]. Wider age ranges and different methods were used in these two studies therefore the results are not directly comparable. For example, the NMR method was used in the first study to compare aging participants, but a gas chromatography-mass spectrometry (GC-MS) metabolomic study was used in the latter study. These differences in methodology might contribute to the different findings. Several studies also investigated the changes in blood metabolomic profiles and the associations with AD pathologies. Increases in blood SM were associated with several brain pathologies verified by MRI and the levels of tau protein and Aβ in the CFS [25, 43]. However, some SMs were inversely associated with amyloid positivity on a PET scan and CSF Aβ, indicating specific associations of these SMs to Aβ in both the brain and CSF [43]. An increase in PC in the blood was also associated with amyloid positivity [44], ventricular volume change [43], and CSF Aβ level [43]. However, some phospholipids were inversely associated with brain pathologies including PC aa C40:6 and PE 39:7[25, 44]. PC ae 42:4 and PC ae 44:4 were negatively associated with Aβ positivity on positron emission tomography (PET) scans of the brain, but were positively associated with ventricular volume changes and an increase in CSF Aβ level [43]. In addition to the phospholipids, acylcarnitine C3 was negatively associated with CSF Aβ [25]. In conclusion, the disturbances of lipid metabolites in dementia and AD patients were observed, as shown by the lowering of acylcarnitine levels and the fluctuation in PCs, SMs, and fatty acids. These findings suggested that a disruption in lipid metabolism occurred in association with AD and these changes were also related to AD pathologies. A study involving a homogenous population and using similar methods should be conducted to verify the results and identify the specific pathways responsible for these metabolic changes.
In addition to the lipids, alterations in other metabolites were also detected in several metabolomic clinical studies (Table 3) [33, 37–42]. Increased secondary bile acid levels and a decrease in primary bile acid level in the circulation were found in AD patients when compared to MCI or control participants [40]. These findings suggested a correlation between gut dysbiosis and cognitive decline, which has been demonstrated in several metagenomic studies [41]. Changes in amino acids and biogenic amines have also been observed in the blood of AD and MCI patients. One study which compared early AD patients with MCI patients, found that the levels of acetylornithine and methionine sulfoxide were increased, while the putrescine was decreased [33]. However, another untargeted metabolomic study found that the level of putrescine was higher in MCI patients who later underwent conversion to AD when compared with normal controls or MCI patients without AD conversion [42]. The differences in the findings of these two studies could be due to the use of different metabolomic methods and the different methods of cognitive assessment used with AD patients. The higher level of cognitive function observed in the latter study may be associated with the higher putrescine level since that study demonstrated that the downstream metabolisms of putrescine differed between MCI and AD patients [42]. Another study also found that the level of arginine, the precursor of polyamines and other amino acids in MCI patients was elevated [34]. These findings indicated disturbances in amino acid and polyamine metabolism which may result in the dysregulation of neurotransmitter metabolism in AD patients. Disturbances in other metabolites including glutamine [38], choline [37], Soraphen A [37], and uric acid [39] were also observed in dementia and AD patients. Some metabolites also showed an association with AD pathologies in other studies [25, 44]. Anandamide was positively associated with Aβ positivity in the brain [44], while valine and serotonin were negatively associated with ventricular volume change and Aβ level in CSF, respectively [25, 43].
THE ASSOCIATION BETWEEN METABOLIC ALTERATIONS AND COGNITIVE FUNCTION IN CLINICAL STUDIES
The changes in metabolites in the blood and the brain were associated with cognitive function as shown in several studies (Table 4) [25, 45]. In the brain, the untargeted-metabolomic study found that the increases in fatty acids in the brain were associated with better cognitive function in AD patients [26]. Regarding blood metabolites, the changes in lipids, bile acids, amino acids, and other metabolites were observed. Some SMs and PCs were found to be associated with worsening cognitive function [25, 31] or increased severity of dementia [43]. However, several studies showed that increased acylcarnitines were associated with better global cognition [31, 32]. The free cholesterol level in HDL and DHA were found to be associated with the global cognitive ability [38]. In the case of other metabolites, increased secondary bile acids were associated with dementia severity [40]. Furthermore, alterations in amino acids and biogenic amines were observed. Arginine [25] and asparagine [31] were associated with poorer cognitive function, while spermidine [25], valine, and alpha-amino adipic acid [43] showed associations with better cognitive function. In the large-scale metabolomic study, glycoprotein acetyl, glutamate, and ornithine were associated with the impaired cognitive ability [38]. Another large untargeted metabolomic study found that maltose and other pertinent metabolites were also associated with worsening global cognitive function [45].
The association between brain/blood metabolites and cognitive function in patients with cognitive decline
AD, Alzheimer’s disease; ADAS-Cog13, Alzheimer’s Disease Assessment Scale–Cognitive Subscale 13 items; CA, cholic acid; CDCA, chenodeoxycholic acid; DCA, deoxycholic acid; DHA, docosahexaenoic acid; EPA, Eicosapentaenoic acid; FIA-MS/MS, flow-injection tandem mass spectrometry; GC-MS, gas chromatography-mass spectrometry; GDCA, glycodeoxycholic acid; GLCA, glycolithocholic acid; HDL, high-density lipoprotein; LC-MS, liquid chromatography-mass spectrometry; LC-MS/MS, liquid chromatography-tandem mass spectrometry; LysoPCs, Lysophosphatidylcholines; MCI, mild cognitive impairment; MMSE, Mini-Mental State Examination; NMR, Nuclear magnetic resonance; PCs, phosphatidylcholine; SMs, sphingomyelin; TDCA, taurodeoxycholic acid; UPLC-MS, ultraperformance liquid chromatography-mass spectrometer; UPLC-MS/MS, ultraperformance liquid chromatography-tandem mass spectrometer.
METABOLIC ALTERATIONS AS PREDICTORS FOR AD AND DEMENTIA IN METABOLOMICS RESEARCH
Some of the metabolomic studies in humans were conducted with prospective design to explore thepredictive biomarkers for AD and dementia (Table 5) [25, 46–48]. One study showed that a panel of twelve metabolites could separate MCI or AD from control healthy participants within two years of the follow-up period [46]. The elevated levels of some SMs also increased the risk of developing AD when compared with non-AD participants within four years of follow-up time [25]. The predictability of phospholipids and acylcarnitines depended on specific species of these molecules since they showed different directions among the specific types [25, 47]. The untargeted metabolomics study with a follow-up time of around eight years found that the alterations of metabolites from the consumption of diets and some endogenous metabolites such as hormones or amino acids were associated with the risk of developing AD from a non-AD state [47]. The increase in a cholesterol ester of HDL and the decrease in branched-chain amino acids were associated with a higher risk of AD [48]. The alterations in these metabolites before the development of AD might be related to the pathophysiologic changes of several organ systems or diet consumption, which could lead to the progression of AD.
Blood metabolites at the baseline as a predictor for cognitive impairment in patients
AD, Alzheimer’s disease; BCAAs, branched-chain amino acids; CA, cholic acid; CMPFP, carboxy-4-methyl-5-pentyl-2-furanpropanoic acid; FIA-MS/MS, flow-injection - tandem mass spectrometry; GC-MS, gas chromatography - mass spectrometry; GC-MS/MS, gas chromatography - tandem mass spectrometry; GDCA, glycodeoxycholic acid; HDL, high-density lipoprotein; HPLC-MS/MS, high-performance liquid chromatography - tandem mass spectrometry; LC-MS, liquid chromatography - mass spectrometry; LC-MS/MS, liquid chromatography - tandem mass spectrometry; LysoPCs, Lysophosphatidylcholines; MCI, mild cognitive impairment; NA, not available; NMR, Nuclear magnetic resonance; PCs, phosphatidylcholine; SMs, sphingomyelin; TDCA, taurodeoxycholic acid; UPLC-MS/MS, ultraperformance liquid chromatography - tandem mass spectrometer.
Patients with MCI have a higher risk of developing AD later in their life. Therefore, several studies focused on the alterations in metabolites in MCI as AD predictors [25, 40]. The sphingolipid SM C18:1 and the phospholipid PC aa 38:4 were associated with a higher risk of AD conversion from MCI [25]. Furthermore, the untargeted-metabolomic study showed that the elevation of 2,4-dihydroxybutanoic acid, PC (16:0/16:0), an unidentified carboxylic acid, lactic acid, and pyruvate, and the reduction of ribose-5-phosphate were observed in MCI patients who later developed AD [40]. The changes in these metabolites are associated with hypoxia, oxidative stress, and lipid membrane remodeling. Additionally, there is evidence that gut dysbiosis is involved in the progression to AD via the alteration of metabolites as indicated by a decrease in primary bile acid and increases in secondary bile acids [40]. In general, metabolomic studies reveal some pathophysiologies of AD that occur before the development of the symptoms.
Regardless of the causes of cognitive impairment, some studies demonstrated that the alterations in blood metabolites were also associated with the incidence of dementia [39, 49]. The increases in docosapentaenoic acid, 3,4-dihydroxybutanoic acid, and uric acid were found in the patients who developed dementia after approximately five years of follow-up [39]. In another study, the alterations in blood metabolites including increased plasma anthranilic acid and glutamic acid, and decreased taurine and hypoxanthine were associated with the risk of dementia development in the next fifteen years [49]. In the large prospective cohort study, one VLDL- and one HDL- specific lipoprotein lipid subclass were associated with increased risk of dementia, while three branched-chain amino acids, creatinine, and two VLDL-specific lipoprotein lipid subclasses were associated with a lower risk of dementia [48]. The alterations in these metabolites might involve the pathophysiologies of cognitive impairment and possibly increase the risk of developing AD in the future. Therefore, the restoration of the levels of these metabolites might be one of the targets to reduce the chance of developing AD.
CONCLUSIONS AND FUTURE DIRECTIONS
The metabolic alterations in both animal and clinical studies are shown in Fig. 2 and the metabolic changes in AD models after received neuroprotective intervention are summarized in Fig. 3. Of these findings, the changes in lipid metabolites were most significant. The changes in lipid levels in the brain, especially of SM and PC, were related to the development of AD most probably because these lipids are the essential constituents of the cell membrane where Aβ processing occurs. The changes in the levels of glucose, TCA metabolites, acylcarnitine, and some fatty acid metabolites could reflect dysregulated energy utilization in the brain. The perturbation of amino acid levels and derivatives might be associated with impaired neurotransmitter function. These findings were closely related to AD status and brain pathologies of AD. Although the alterations in blood metabolites could not directly reflect the pathologic changes in the brain, they may be associated with systemic disease or health status which could precipitate the development of dementia and AD. The disturbances in blood amino acids and their derivatives, bile acids, and lipid metabolites in AD and dementia patients indicated that systemic oxidative stress, gut dysbiosis, and impaired lipid metabolism might involve alteration of blood metabolites which could directly or indirectly affect brain function and AD pathologies. The authors suggest that a prospective design of a further study into metabolomics and AD will give vital information regarding the metabolic changes which occur before disease manifestation. Findings from prospective studies will be useful in understanding the pathophysiology behind these metabolic changes and their association with both dementia and AD. Furthermore, details regarding the perturbations of metabolites found in prospective research could be used as identifiable novel biomarkers in making an early diagnosis of dementia and AD. That information could help the patients to receive earlier treatment as well as potentially increase the possibility of slowing down the progression of the cognitive impairment.
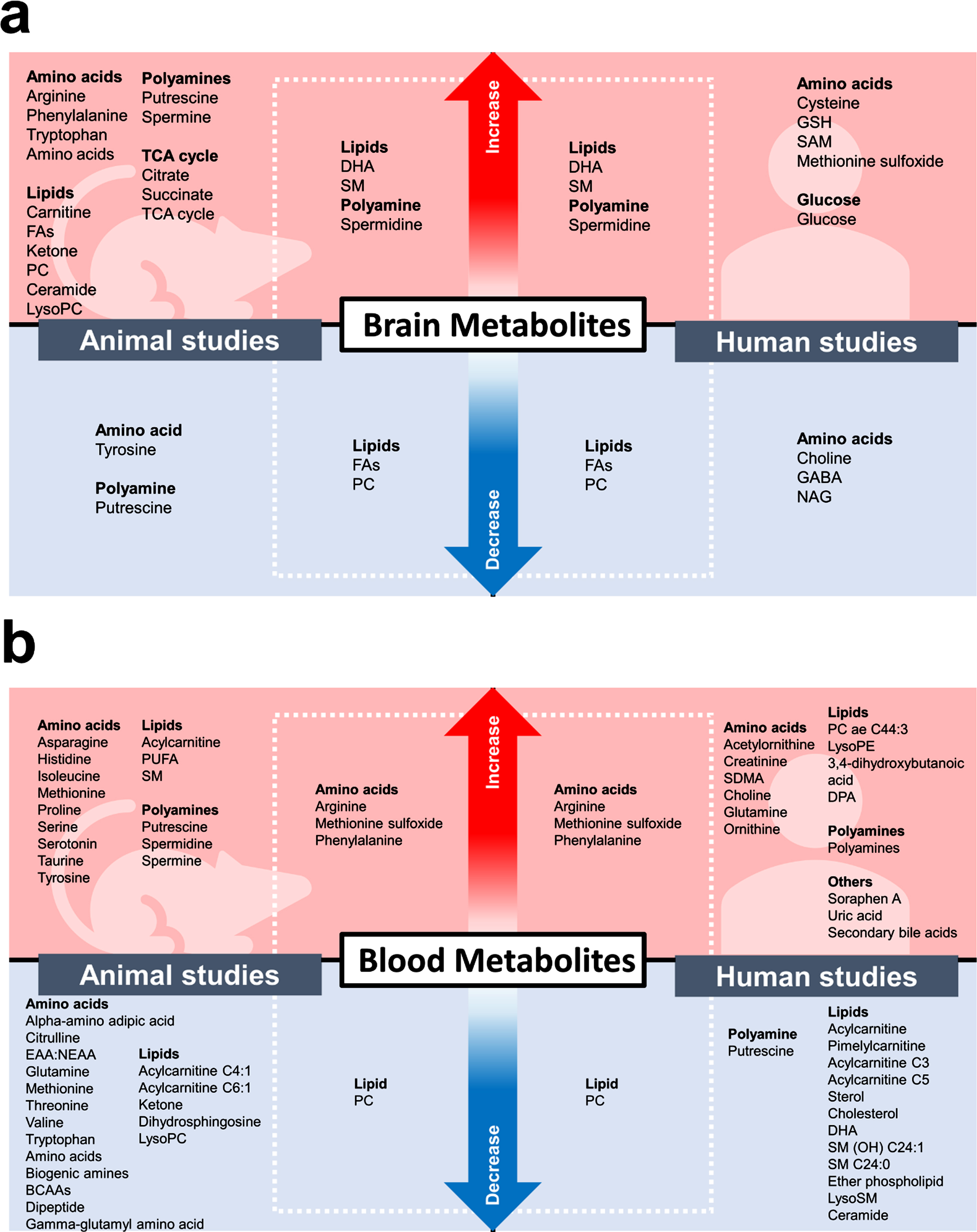
The alterations in metabolites found in the brain (a) and the blood (b) in animal and human metabolomic-based studies. BCAAs, branched-chain amino acids; DHA, docosahexaenoic acid; DPA, docosapentaenoic acid; EAA, essential amino acids; FA, Fatty acid; GABA, gamma-Aminobutyric acid; GSH, glutathione; NAG, N-acetylglutamate; NEAA, non-essential amino acids; PC, Phosphatidylcholine; PE, Phosphatidylethanolamine; PUFA, Polyunsaturated fatty acid; SAM, S-adenosylmethionine; SDMA, symmetric dimethylarginine; SM, Sphingomyelin; TCA, tricarboxylic acid.
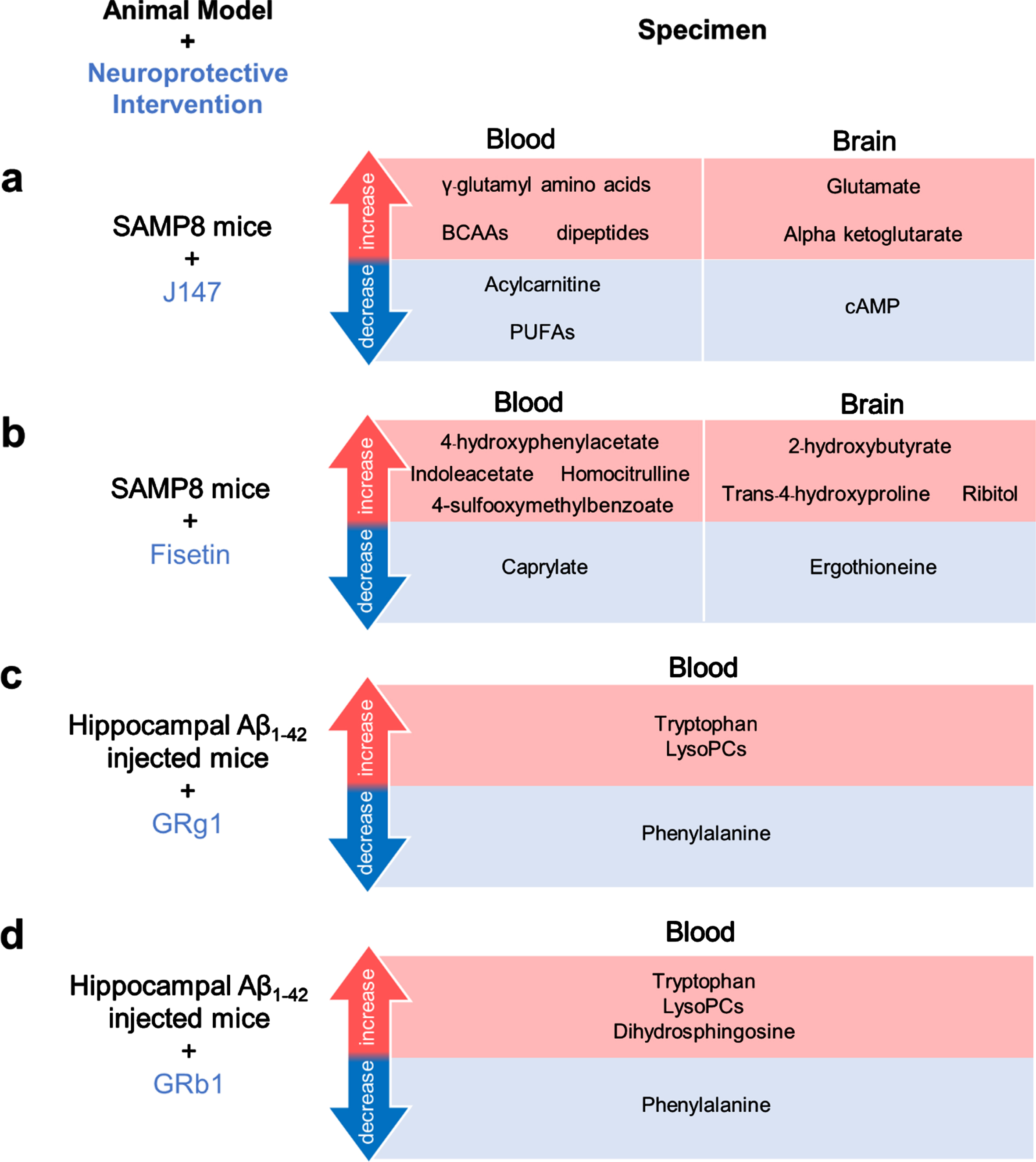
The alterations in metabolites found in the blood and/or the brain from in vivo models of Alzheimer’s disease following neuroprotective interventions. (a) Male SAMP8 mice after J147 treatment, (b) Male SAMP8 mice after Fisetin treatment, (c) Male Khun ming mice injected with Aβ1-42 into the hippocampus after GRg1 treatment, and (d) Male Khun ming mice injected with Aβ1-42 into the hippocampus after GRb1 treatment. Aβ, amyloid-β; BCAAs, branched-chain amino acids; cAMP, cyclic adenosine monophosphate; LysoPCs, Lysophosphatidylcholines; PUFA, Polyunsaturated fatty acid; SAMP8, senescence-accelerated prone 8.
There are some points to be included in a selection of the focus of future research. Most of the focus needs to be on the changes in the lipid metabolites which are related to the metabolism of the cell membrane, which is important for the function of the central nervous system. In addition, the pathophysiology of cognitive impairment is partially linked with the alterations of neurotransmitters in the brain. However, the knowledge surrounding these is still limited and inconsistent, and this information needs further investigation. Additionally, only a few clinical studies have investigated the association of metabolic alterations in both the brain and blood. The studies presented in this review mostly investigated the brain or the blood separately. Since the generalcharacteristics of participants and the criteria for diagnosis or categorization in each study differ extensively, these limit the interpretation of metabolic disturbances in the brain and the blood among the studies. Some of the animal studies included both brain and blood metabolomics, but the results were still controversial. In addition, when interpreting the data, it is important to accept that the metabolites between animal and human studies could be different.
Footnotes
ACKNOWLEDGMENTS
This work was supported by Senior Research Scholar Grant from the National Research Council of Thailand (SCC.), the NSTDA Research Chair Grant from the National Science and Technology Development Agency Thailand (NC), and the Chiang Mai University Center of Excellence Award (NC).