
Abstract
Background:
The interactions between environmental factors and genetic variants have been implicated in the pathogenesis of Alzheimer’s disease (AD). The altered gut microbiota (GM) and vitamin D deficiency are closely associated with the higher risk of AD.
Objective:
This study was performed to evaluate whether the crosstalk between GM and single nucleotide polymorphisms (SNPs) of vitamin D receptor (VDR) or vitamin D binding protein (VDBP) have a link with the risk of amnestic mild cognitive impairment (aMCI) in the Chinese elderly population.
Methods:
A total of 171 aMCI patients and 261 cognitive normal controls (NC) were enrolled in this study. Six tag SNPs of VDR and VDBP were genotyped by PCR-RFLP. The serum levels of vitamin D, Aβ1-42, and p-tau (181P) were determined by using of ELISA kits. The alterations in the GM were analyzed by full-length 16S ribosomal RNA (rRNA) gene sequencing.
Results:
The frequencies of AG genotype and A allele of VDR rs1544410 in aMCI group were significantly higher than that in NC group (genotype: p = 0.002, allele: p = 0.003). Patients with aMCI showed an abnormal GM composition compared with NC group. Interestingly, significant differences in GM composition were found between aMCI and NC group among individuals with AG genotype, as well as between individuals with AG and GG genotype of VDR rs1544410 among patients with aMCI.
Conclusion:
These results implicated that the crosstalk between gut microflora and vitamin D receptor variants are associated with the risk of aMCI in Chinese elderly population.
Keywords
INTRODUCTION
Alzheimer’s disease (AD) is a chronic neurodegenerative disease characterized by a progressive decline in memory and other cognitive impairment [1]. Mild cognitive impairment (MCI) is considered to be a transitional state between AD and normal aging, which is an important stage of prevention, prognosis, and treatment in the management of AD [2]. It is estimated that in 2019 the prevalence of dementia was nearly 57.4 million cases worldwide, and this figure is expected to triple by 2050. That means the number of dementia cases in China will increase from 16.5 million in 2020 to 48.98 million in 2050 [3–5]. In China, AD is also the leading cause of disability in people older than 65 years [6] and has become the fifth cause of death after stroke, ischemic heart disease, chronic obstructive pulmonary disease, and lung cancer [7]. Although the pathogenesis of AD remains unclear, increasing evidence has shown that interaction between genetic and environmental risk factors have been implicated in the pathogenesis of cognitive impairment in the aging population [8].
Accumulating evidence has shown that vitamin D exerts neuroprotective effects in the development of neurodegenerative disease [9–11], and vitamin D deficiency may contribute to brain decline and increase the risk of AD [12–14]. Epidemiological studies have suggested that vitamin D is closely associated with the risk of AD in the elderly population [15, 16]. Research has shown that the single nucleotide polymorphisms (SNPs) of genes related to vitamin D synthesis and metabolism affect the circulating level of vitamin D [17]. Equally notable is that vitamin D binding protein (VDBP) rs2282679 may influence AD pathology independently of vitamin D [15]. In addition, previous studies have confirmed that vitamin D receptor (VDR) SNPs (rs1544410, rs731236, rs7975232, and rs10735810) are closely related to cognitive function [18–21]. Nevertheless, the subjects of these studies were mainly Europeans and North Americans. There are few reports on the relationship between VDBP/VDR gene SNPs and cognitive impairment in Chinese elderly population.
The gut microbiota (GM) is composed of trillions microbial cells in the gastrointestinal tract and plays a critical homeostatic role for host health. Studies in animal models have implicated that impaired cognition is associated with the alterations of GM [22]. As a dietary nutrient, vitamin D deficiency, high-dose vitamin D supplementation, or ultraviolet B light (UVB) overexposure are strongly linked to intestinal microbial homeostasis [23, 24]. Some studies have shown that the microbial communities in human blood, saliva, and intestine are involved in the occurrence and development of AD [25–30]. Recent studies suggest that GM communities have been a potential key player in the pathology of AD and the crosstalk between gut and brain has provided some implications for immunopathogenesis and treatment strategies of AD [30–32]. The intestinal microbiota and its metabolites may participate in the onset and progression of AD via regulating microglia activation, neuroinflammation, oxidative stress, amyloid-beta aggregation, and blood–brain barrier permeability [33–35]. As it is known, sufficiently high quality ultra-long reads can be generated by the full-length 16S rRNA sequencing to offer higher taxonomic resolution than partial 16S rRNA gene short-read sequencing [36, 37]. Considering that previous studies were limited to 16S rRNA gene sequencing profiling taxonomic resolution, and the difference of dietary habits between the north and the South in China (previous studies were conducted in southern China), and the influence of region and diet on the composition of intestinal flora, the further investigations are still needed to confirm the different composition of GM between patients with aMCI and healthy controls in northern China. A genome-wide association analysis has found a variation in the VDR gene that influences the composition of host GM [38]. Recently, increasing attention has been paid to the interactions between genetic variants and environmental risk factors in the etiology of disease [39]. Evidence has suggested that the interaction between environmental factors and genetic variants contributes to the pathogenesis of AD [40, 41]. However, little is known about the roles of interplay between VDBP/VDR gene polymorphisms and GM in the pathogenesis of amnestic mild cognitive impairment (aMCI). So we hypothesized that the crosstalk between GM and the variants of VDR or VDBP genes have a link with the risk of aMCI in the Chinese elderly population.
In the present study, we conducted a cross-sectional study to investigate the links of interactions between the altered GM and SNPs of VDR/VDBP genes with the risk of aMCI in the Chinese elderly population. A total of 171 aMCI patients and 261 matched were enrolled in this study. The SNP loci associated with aMCI were identified by PCR-RFLP. Fecal microbiome were tested using full-length 16S rRNA gene amplicon sequencing. Finally, the metabolic pathways of altered gut microbiome were predicted by PICRUSt2. This study will provide a basis for further investigating the roles of the interactions between genetic and environmental factors in the pathogenesis of AD.
MATERIALS AND METHODS
Subjects
The subjects of this study are patients with aMCI and cognitively normal individuals. All of them are over 60 years old. A total of 171 aMCI patients and 261 cognitive normal controls (NC) were recruited from community health service centers in Shijiazhuang city, Hebei Province, China, which is located at E114°28′ and N38°02′, from July 2018 to July 2019.
The participants were diagnosed as aMCI according to the Petersen’s criteria (P-MCI) as follows: 1) chief complaints of memory loss (especially confirmed by family members or other informants); 2) possessing objective memory impairment; 3) global cognitive functioning assessed by Mini-mental State Examination (MMSE) scores between 19 and 27 (cutoff points for illiterate≤19, primary school≤24, and secondary school and above≤27) and Montreal Cognitive Assessment (MoCA) scores between 13 and 24 (cutoff points for illiterate≤13, primary school≤19, and secondary school and above≤24); 4) normal ability of daily living; 5) no dementia. In our study, all participants were diagnosed and evaluated by at least two experienced neurologists.
Patients suffering from severe psychiatric disorders, nervous system diseases, poor hearing, and vision, or consuming psychotropic medicines were excluded. In addition, the subjects with hypertension, diabetes, and cancer or who were being treated wtih drugs were also excluded for microbiome analysis. This study was approved by the ethics committee of the First Hospital of Hebei Medical University and the written informed consent was obtained from each subject.
SNP selection
The SNPs of VDR or VDBP were selected from the public SNP database, NCBI dbSNP database (https://www.ncbi.nlm.nih.gov/snp/), with conditions limited to the Chinese Han Beijing population (CHB), as well as the associated published studies. Finally, a total of 6 SNPs were selected for analysis, including two VDBP SNPs (rs4588 and rs7041) and four VDR SNPs (rs7975232, rs1544410, rs731236, and rs2228570) (Table1).
Description of SNPs in VDBP and VDR genes
DNA extraction
Blood samples were collected from subjects before breakfast in the morning. The total genomic DNA was extracted from the whole blood using the Wizard® Genomic DNA Purification Kit (Promega, Madison, WI, USA) according to the manufacturer’s instructions. NanoDrop® ND-1000 (Thermo Fisher Scientific, Waltham, MA, USA) was applied to determine the density and purity of genomic DNA.
SNP genotyping
VDR polymorphisms (rs7975232, rs1544410, rs731236, and rs2228570) and VDBP polymorphisms (rs4588 and rs7041) were assessed by PCR-RFLP. Primers examining the polymorphism were prepared as previously described [42, 43]. PCR products were digested with restriction enzymes, as demonstrated in Table 2.
Primer sequences of the rs7041, rs4588, rs7975232, rs731236, rs2228570, and rs1544410 SNPs
RFLP analysis (Thermo Scientific™) was performed to identify genotypes of 6 polymorphisms: rs7041 by FastDigest BsuRI restriction enzyme, rs4588 by FastDigest StyI restriction enzyme, rs7975232 by FastDigest ApaI restriction enzyme, rs731236 by FastDigest TaqI restriction enzyme, rs2228570 by FastDigest FokI restriction enzyme, and rs1544410 by FastDigest BsmI restriction enzyme (Fig. 1A).
Three genotypes of each polymorphism were identified by sequencing the PCR products with Sanger chain termination method. The polymorphism of PCR-RFLP analysis was double determined (Fig. 1B). Sequencing was performed by General Biosystems (General Biosystems Co. Ltd. Anhui, China).
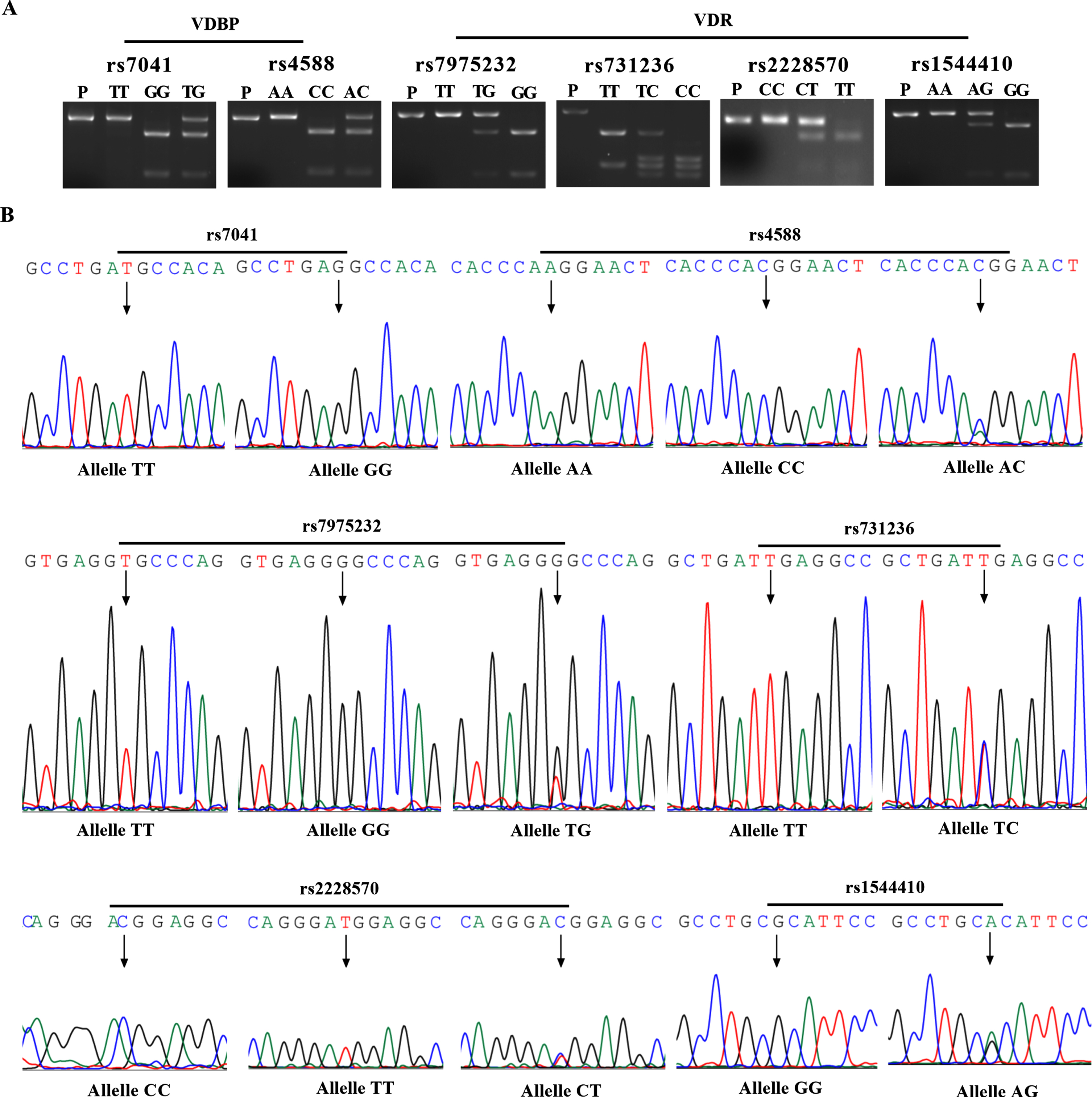
The SNPs of VDBP and VDR were detected by restriction fragment length polymorphisms (RFLP) and DNA sequencing. A) RFLP of rs7041 and rs4588 in VDBP gene, and rs7975232, rs731236, rs2228570, and rs1544410 in VDR gene by FastDigest BsuRI, StyI, ApaI, TaqI, FokI, and BsmI restriction enzyme, respectively. B) Sanger sequencing chromatograms corresponding to confirmed SNPs in VDBP and VDR genes. P, PCR Product.
Measurement of serum vitamin D, Aβ1-42, and p-tau (181P)
Blood samples were collected from subjects before breakfast in the morning. Serum levels of vitamin D, Aβ1-42, and p-tau (181P) were measured by using the human VD ELISA kit (intra-assay precision: CV% <10%; inter-assay precision: CV% <15%) (Shanghai Li Chen Biotechnology Co. Ltd. Shanghai, China), human Aβ1-42 ELISA kit (intra-assay precision: CV% <15%; inter-assay precision: CV% <15%) and p-tau (181P) ELISA kit (intra-assay precision: CV% <15%; inter-assay precision: CV% <15%) (JiangSu Mei Biao Biological Technology Co. Ltd. Yancheng, China) following the manufacturer’s instructions. Serum concentrations of vitamin D, Aβ1-42, and p-tau (181P) were determined from the lowest dilution of serum samples within a standard curve and reported as ng/mL, pg/mL, and pg/mL respectively. Each ELISA plate contained a positive reference standard to generate a standard curve. The absorbance was detected at the wavelength of 450 nm by a microplate reader (Promega, Madison, WI, USA). Each values were determined from the absorbance values of the standards, using linear regression (vitamin D: R2 = 0.98; Aβ1-42: R2 = 0.98; p-tau (181P): R2 = 0.99).
Stool sample collection and DNA extraction
All stool samples were freshly collected from each participant using sterile collection containers and immediately preserved in anhydrous ethanol [44]. The stool samples were sub packed in 2 ml Eppendorf tubes and immediately placed in –80°C until analysis. Total genomic DNA was extracted from stool samples using a genomic DNA extraction kit (Promega, Madison WI, USA). The quality, quantity, and purity of extracted genomic DNA were determined by 1.0% agarose gel electrophoresis and NanoDrop® ND-1000.
Amplicon sequencing
The total bacterial genomic DNA was used as a template for PCR amplification of V1–V9 region of the bacterial 16S ribosomal RNA gene in a multiplex approach with the primers 27F (AGAGTTTGATCCTGGCTCAG) and 1492R (GGTTACCTTGTTACGACTT). PCR amplification was carried out using TransStart® FastPfu DNA Polymerase (TransGen Biotech, Beijing, China) under the following cycling conditions: 98°C for 2 min, followed by 35 cycles of 95°C for 30 s, 60°C for 45 s, and 72°C for 90 s, followed by a final step at 72°C for 10 min. Amplified products were detected by 2% agarose gel electrophoresis. Equal amounts of amplified products were pooled according to the concentration, which detected by 2% agarose gel electrophoresis. The target bands were purified using a QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The sequencing adaptors were ligated onto the purified amplified products through DNA binding enzyme, and AMpure PB magnetic beads were used to purify and select the DNA fragments to construct the SMRT bell library. The constructed library was quantified by Qubit, and the insert size was detected by Agilent 2100, and then sequenced by PacBio Sequel (PacBio, Menlo Park, CA, USA). These works were performed by Novogene (Beijing, China).
Statistical analysis
The data obtained from the clinical examination and experimental results were tabulated and statistically analyzed using SPSS21.0 (SPSS, Inc., Chicago, IL, USA) and GraphPad Prism 8.0 (GraphPad Software, Inc., San Diego, CA, USA). Characteristics of the subjects were presented as mean ± SD for continuous variables and as proportions for categoric variables. Student’s T-test or Mann-Whitney test and χ2 test were used to analyze the quantitative data and categorical, respectively. The Hardy Weinberg equilibrium (HWE) was tested for each SNP using Pearson’s χ2 test. Genotype and allele frequencies of VDBP or VDR polymorphisms were compared using χ2 test or Fisher’s exact test. p-value less than 0.05 were regarded as statistically significant.
The GM findings were analyzed using Quantitative Insights into Microbial Ecology (Qiime) software (Version 1.9.1) [45]. Alpha diversity and beta diversity index analyses were conducted in R software (Version 2.15.3). Wilcoxon’s rank sum tested for alpha diversity, and Adonis tested for microbial community clustering (PCoA) using binary_jaccard distance matrices. The linear discriminant analysis (LDA) effect size (LEfSe) was carried out using LEfSe software (Version 1.0). These analyses were performed by NovoMagic v3.0 (Novogene Technology, Beijing, China) website. Functional pathways of the altered faecal microbiota were predicted with Phylogenetic investigation of communities by reconstruction of observed states 2 (PICRUSt2) (https://github.com/picrust/picrust2) (GitHub Inc. San Francisco, CA, USA) [46].
RESULTS
Demographic characteristics
The demographic characteristics of the subjects in NC and aMCI groups were summarized in Table 3. The data included age, gender, education, the MMSE scores, and the MoCA scores. There were no significant differences between the aMCI group and NC group in gender and education years (p > 0.05). The subjects of the aMCI group have significantly lower MMSE scores (p < 0.001) and MoCA scores (p < 0.001) than those of which in NC group.
Demographic characteristics of the study sample
Wilcoxon signed-rank and χ2 tests were used. MMSE, Mini-Mental State Examination; MoCA, Montreal Cognitive Assessment; AFT, Animal Fluency Test; BNT, Boston Naming Teat; DSF, Digit Span Forward; DSB, Digit Span Backward; CDT, Clock Drawing Test; TMT-A, Trail Making Test-A; TMT-B, Trail Making Test-B.
Bsml polymorphism in VDR gene is closely related to the risk of aMCI
Except for the rs4588 in the VDBP gene (p < 0.05), the polymorphisms of rs7041 in the VDBP gene and rs7975232, rs1544410, rs731236, and rs2228570 in the VDR gene were all in Hardy-Weinberg equilibrium (p > 0.05, data not shown) in the aMCI and NC groups. As shown in Table 4, there were no statistically significant differences (p > 0.05) in genotype or allele frequencies between aMCI and NC groups for 5 individual SNPs (rs7041 and rs4588 in VDBP gene, rs7975232, rs731236, and rs2228570 in the VDR gene). But the frequencies of AA genotype and A allele of VDR rs1544410 in aMCI group were higher than that in the NC group (genotype: p = 0.002, allele: p = 0.003).
Genotypic and allelic frequencies of the 6 SNPs in VDBP and VDR genes
Comparisons were performed using the Pearson χ2 test or Fisher’s exact test. Indicates the statistical significance after Bonferroni correction. OR, odds ratio; CI, confidence interval.
Lower Aβ1-42 levels and higher p-tau (181P) levels in serum of aMCI patients
We randomly selected 43 aMCI patients and 31 cognitive normal controls from the subjects and detected the serum levels of Aβ1-42 and p-tau (181P) with ELISA kit. As shown in Fig. 2, the subjects in aMCI group had significantly lower Aβ1-42 levels (p < 0.001) (Fig. 2A) and higher p-tau (181P) levels (p < 0.001) (Fig. 2D) compared with NC group. Spearman’s correlation analysis was used to assess the potential correlations between the serum Aβ1-42 or p-tau (181P) levels and MMSE/MoCA scores respectively. The results showed a positive correlation between the serum Aβ1-42 levels and MoCA scores (r = 0.26, p = 0.02) (Fig. 2C), but there was no correlation with MMSE scores (r = 0.12, p = 0.3) (Fig. 2B); there was a negative correlation between serum p-tau(181P) and MMSE (r = –0.40, p = 0.0009) (Fig. 2E)/MoCA (r = –0.44, p = 0.0002) (Fig. 2F) scores.

The serum levels of Aβ1-42, p-tau (181P) protein, and vitamin D in NC or aMCI groups and its correlations with cognitive function. A) The serum levels of Aβ1-42 in NC and aMCI groups were detected by ELISA. B, C) The correlations between serum levels of Aβ1-42 with MMSE (B) or MoCA (C) scores by Spearman’s correlation coefficient. D) The serum levels of p-tau (181P) protein in NC and aMCI groups were detected by ELISA. E, F) The correlations between serum p-tau (181P) levels with MMSE (E) or MoCA (F) scores by Spearman’s correlation coefficient. G) The levels of serum vitamin D in aMCI patients (n = 47) were significantly lower than that in NC group (n = 47). H, I) The correlations between serum vitamin D levels with MMSE (H) or MoCA (I) scores by Spearman’s correlation coefficient. J) ROC curve of serum vitamin D levels was established to distinguish aMCI from NC group.
Decrease of serum vitamin D levels increased the susceptibility of aMCI in elderly population
To evaluate the association between the serum vitamin D levels and the risk of aMCI in elderly population, we randomly selected 47 aMCI patients and 47 cognitive normal controls from the subjects, and the serum vitamin D levels were detected with ELISA kit. As shown in Fig. 2, the subjects in aMCI group had significantly lower serum vitamin D levels (p < 0.05) than that in NC group (Fig. 2G). The results of Spearman’s correlation analysis indicated a positive correlation between the serum levels of vitamin D with MMSE scores (r = 0.23, p = 0.02) (Fig. 2H), as well as with MoCA scores (r = 0.27, p = 0.008) (Fig. 2I). Next, we evaluated the predictive value of serum vitamin D levels for the diagnosis of aMCI patients. ROC curves analysis indicated that the serum vitamin D levels had an AUC of 0.64 (95% CI: 0.53–0.75) (Fig. 2J). When the Youden’s index reaches its maximum value 1.26, the cutoff point for serum vitamin D levels was 37 ng/mL, with 49% sensitivity and 78% specificity. The results suggested that the serum vitamin D levels potentially had diagnostic value for aMCI patients to some extent.
Association of VDR gene variants with the gut microbiota
To investigate the association of VDR gene variants with the gut microbiota in aMCI patients, the microbial community features were analyzed by full-length 16S ribosomal RNA (rRNA) gene sequencing. Although no significant differences in microbial alpha and beta diversity were found between individuals with AG and GG genotype in VDR among total samples (Fig. 3A, D, G). However, the results of alpha diversity analysis showed that there were significant differences between subjects with AG and GG genotype of VDR in NC (Simpson index: p = 0.03; goods_coverage index: p = 0.02) and aMCI group (goods_coverage index: p = 0.002). The results of PCoA plot analysis based on binary_jaccard distances suggested that dispersion between subjects with AG and GG genotype VDR in aMCI group (Adonis; p = 0.02) (Fig. 3I) was significantly different, but not in NC group (Adonis; p = 0.5) (Fig. 3H). LEfSe analysis results indicated that the fecal microbiota diversity differed between aMCI and NC groups. The aMCI group had greater abundance of the Agathobacter, Catabacter, butyrate_producing_baterium_A2_175, TM7_phylum_sp_oral_clone_HD027, and Actinobateria, and the NC group had greater abundance of the Aeromonadales (data not shown). The abundance of more than 19 bacterial taxa in fecal microbiota composition were significantly different between subjects with AG and GG genotype VDR in aMCI group (Fig. 3J). We further evaluated the correlation of the relative abundances of bacterial taxa at genus level with cognitive test scores in aMCI patients (Fig. 3K). The genera of Subdoligranulum, Holdemania, peptostreptococcus and Gemella were positively correlated with cognitive scores, and the genera of Bacteroides, Megasphaera, Akkermansia, Acinetobacter, Enterococcus, Turicibacter, Kocuria, Candidatus_Soleaferrea, Ruminiclostridium, Comamonas, Mitsuokella, and Adlercreutzia were negatively correlated with cognitive scores in aMCI patients.

The microbial community features of individuals with AG or GG genotype in rs1544410 of VDR polymorphism in NC and aMCI population. Species richness was calculated using Simpson (A-C) and goods_coverage (D-F) index, and the Beta diversity (G-I) was calculated for individuals with AG or GG genotype in the context of total, NC, and aMCI groups using binary_jaccard dissimilarity matrix. A p-value <0.05 is considered significant. Total indicated the combination of NC and aMCI group. J) Histogram of the LDA scores by LEfSe analysis for different abundant taxa in aMCI patients with AG (red) or GG genotype (green) in rs1544410 of VDR gene. The LDA scores (log10)>2 and p < 0.05 are listed. K) The correlations between fecal microbiomes with MMSE scores, MoCA scores, or serum levels of vitamin D at genus level in patients with aMCI.
Interactions of VDR gene variants and the gut microbiota on the risk of aMCI
Giving that the VDR gene variants were associated with the gut microbiota and the altered bacterial taxa at genus level were correlated with cognitive test scores in aMCI patients, we further evaluated the interactions of VDR gene variants and the gut microbiota on the risk of aMCI in elderly population. No significant differences in microbial alpha and beta diversity were found between NC and aMCI groups among the subjects with GG genotype or the combination of GG and AG genotype in VDR gene (Fig. 4A, D, G, J). But for those subjects with AG genotype in VDR gene, the alpha diversity showed significant differences between NC and aMCI group (observed_species index: p = 0.01; Chao1 index: p = 0.01; ace index: p = 0.01) (Fig. 4B, E, H), as well as the beta diversity in PCoA plot based on binary_jaccard (R2 = 0.07, p = 0.01). The results of LEfSe analysis suggested that the fecal microbiota diversity differed between aMCI and NC groups. The aMCI group had greater abundance of the Agathobacter, Catabacter, butyrate_producing_baterium_A2_175, TM7_phylum_sp_oral_clone_HD027 and Actinobateria, and the NC group had greater abundance of the Aeromonadales (data not shown). Interestingly, more than 23 bacterial taxa in fecal microbiota composition were confirmed to be the distinguished microbes in aMCI and NC groups among the subjects with AG genotype in VDR (Fig. 4M). As indicated in Fig. 4N, the relative abundances of some genus were positively, and the others were negatively linked with the cognitive test scores and vitamin D levels.
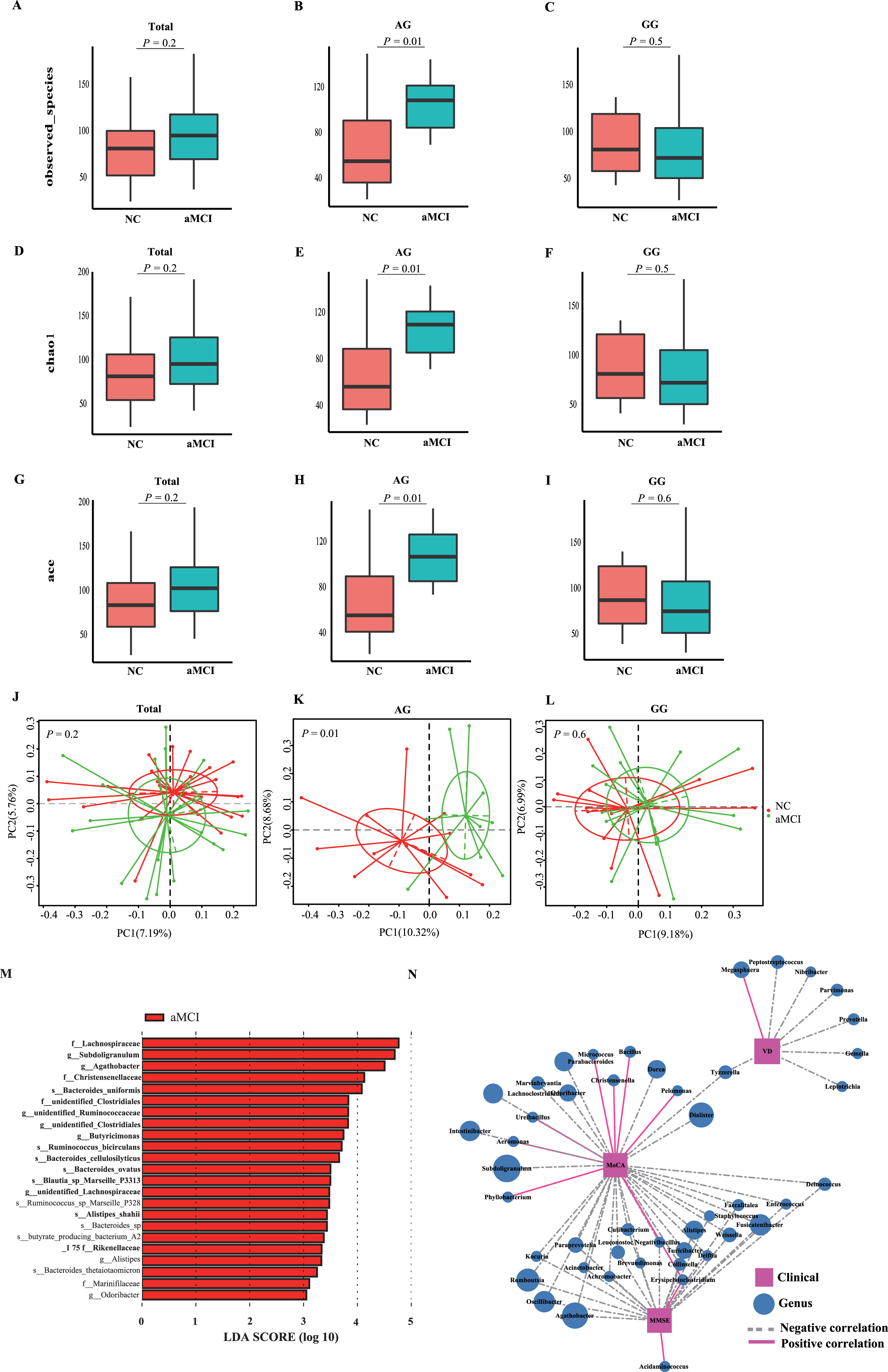
The microbial community features of NC or aMCI patients in the population with AG or GG genotype in rs1544410 of VDR polymorphism. Species richness was calculated using observed_species (A-C), Chao1 (D-F), and ace (G-I) index, and the Beta diversity (J-L) was calculated for NC or aMCI patients in the context of total, AG, and GG genotype population using binary_jaccard dissimilarity matrix. A p-value <0.05 is considered significant. Total indicated the combination of individuals with AG and GG genotype. M) Histogram of the LDA scores for different abundant taxa in aMCI patients (red) and NC (green) (not shown in the figure) with AG genotypein rs1544410 of VDR gene. The LDA scores (log10) >2 and p < 0.05 are listed. N) The correlations between fecal microbiomes with MMSE scores, MoCA scores or serum levels of vitamin D at genus level in subjects with AG genotype.
Bacterial metabolic pathways predicted by PICRUSt2
In order to understand the functional links between the changed intestinal microbiota profiles with aMCI, the metabolic pathways related to the altered fecal composition in aMCI patients with AG or GG genotype in rs1544410 of VDR polymorphism were predicted using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database [46]. Here the top 24 metabolic pathways were presented, and none of the significant differences among the four groups were identified in the KEGG level 2 pathways (Fig. 5A). At KEGG level 3, according to the microbiota profile changes between NC and aMCI group among subjects with AG genotype in rs1544410 of VDR polymorphism, the analysis results revealed that the metabolic pathways of glycine, serine, and threonine metabolism (Fig. 5B), phenazine biosynthesis (Fig. 5D), and protein processing in endoplasmic reticulum (Fig. 5F) were enriched in NC groups; and the selenocompound metabolism (Fig. 5C) was enriched in aMCI groups. Comparing the metabolic pathways between subjects with AG and GG genotype in rs1544410 of VDR polymorphism among aMCI patients, we found that phenazine biosynthesis (Fig. 5D) and polycyclic aromatic hydrocarbon degradation (Fig. 5E) were enriched in subjects with GG genotype, and Toll and Imd signaling pathway was enriched in subjects with AG genotype (Fig. 5G). This result indicated that the crosstalk between the VDR gene polymorphism and intestinal microbiota were linked with the risk of aMCI in elderly population.
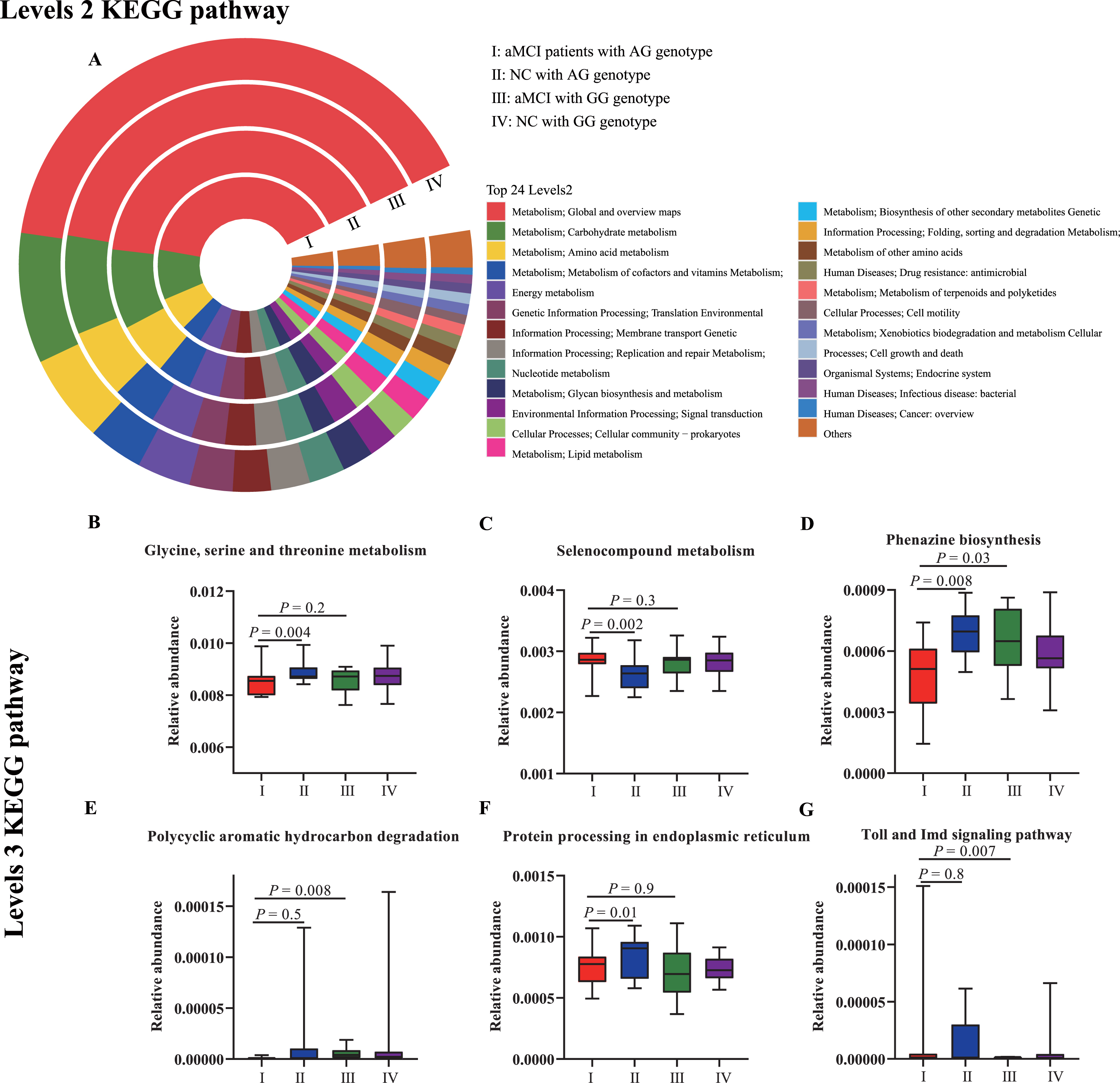
Predicted metabolic pathways related to the altered fecal composition in aMCI patients with AG or GG genotype in rs1544410 of VDR polymorphism. The principal pathways of the gut microbiota were inferred from full 16S rRNA gene amplicon sequences in feces collected from aMCI patients with AG genotype (I), NC with AG genotype (II), aMCI with GG genotype (III), and NC with GG genotype (IV) by PICRUSt2 algorithm. A) A comparison of the enriched markers on level 2 of the KEGG functional category. And there no pathways at level 2 were significantly among the four groups. B-G) A comparison of the enriched markers on level 3 of the KEGG functional category.
DISCUSSION
The classical functions of vitamin D are to regulate calcium-phosphorus homeostasis and control bone metabolism. However, it has been reported that patients with AD or aMCI have a significantly lower level of plasma vitamin D, compared with the subjects with normal cognitive function [47]. A systematic evaluation conducted by Alzain et al. also reported that vitamin D deficiency is very common in patients with cognitive impairment, suggesting vitamin D deficiency is related to the increased risk of dementia [48]. It is reported that maintaining an adequate vitamin D status may reduce the risk of AD in a Mendelian randomization study [49], and a meta-analysis study concluded that the average concentration of serum vitamin D in AD patients was lower than that in the control group [50]. Vitamin D supplementation improves cognitive function in the elderly with MCI [51]. Przybelski et al. used logistic regression analysis in the retrospective investigation of the elderly over 60 years old and found that the plasma 25(OH)D levels are positively correlated with MMSE scores [52]. In this study, our results suggested that the serum levels of Aβ1-42 were lower and the p-tau (181P) levels were higher in patients with aMCI, and the serum vitamin D levels of aMCI patients were lower than that of cognitive normal controls, which suggested that the level of vitamin D is lower in patients with cognitive impairment. In addition, ROC curve is a comprehensive index reflecting the sensitivity and specificity of continuous variables. The results of ROC curve analyses indicated the serum vitamin D levels with a 49% sensitivity and 78% specificity for screening aMCI patients, seem to be not a good biomarker, but may play an important auxiliary role in the diagnosis of aMCI.
Extensive evidence suggests that vitamin D deficiency has been associated with cognitive impairment and AD [21, 53]. Abnormal expression of VDBP and VDR genes, the key genes for vitamin D transport and exerting functions, may affect the level of vitamin D and its biological function in vivo, suggesting that genetic variations may be involved in the risk of aMCI. It was reported in elderly Uygur people that the A allele of VDR gene ApaI and the T allele of VDR gene BsmI may be related to the pathogenesis of aMCI [54]. A Netherlands study had shown that carriers of BsmI and TaqI variant alleles of VDR have the poor general cognitive ability [55]. In our study, we found that the VDR rs1544410 (BsmI) A allele was associated with the increased risk of aMCI as compared with the G allele. However, not as the studies in Western populations [20, 55], our study did not observe the significant association between VDR gene polymorphisms (ApaI, TaqI, and FokI) and the risk of aMCI in Chinese elder people. Previous studies have reported that rs7041, rs4588, and rs2282679 polymorphisms of the VDBP gene are the determinants of vitamin D reduction in body [56]. Mokry et al. proposed SNP rs2282679 as a strong independent predictor of AD [15]. In the Chinese Han population, the rs4588 and rs2282679 polymorphism are significantly related to 25(OH)D level [57]. However, in this study, we did not find that rs7041 and rs4588 in the VDBP gene are associated with the risk of aMCI. That may be the results of the comprehensive effects, such as a different race, geographical location, cultural background, and genetic background.
Multiple lines of evidence from animal and human studies have suggested that the intestinal, blood, and salivary microbiota involved in the pathogenesis of AD [25–29]. VDR plays an essential role in the composition and metabolites of gut microbiota, which were identified in the latest research on intestinal epithelium-specific VDR knock out (VDRΔ IEC) mice and myeloid cell-specific (VDRΔ lyz) mice [58]. Reduction of intestinal VDR has been reported in aging and various inflammatory diseases [59]. VDR is highly expressed in the epithelial cells of intestinal tract [60]. Downregulation or deficiency of VDR in the intestinal epithelial cells changes the composition of the GM through Paneth cells, which play a key role in establishing and maintaining the intestinal microbiota [60]. VDR genetic variants influence VDR activity, and the most common SNPs that influence VDR expression within the immune system include rs1544410, rs7975232, rs731236, and rs10735810 [61]. Additionally, genome-wide association analysis has shown that VDR polymorphisms also influence the composition of GM [38]. A study in Saudi Arabia reported VDR TaqI, ApaI, and BsmI influence the vitamin D/VDR axis play a role in obesity, which may be due to inflammation caused by changes in microbial translocation and intestinal permeability [62]. Meta analyses also found that an association between the VDR rs1544410 and increased odds of MCI [63]. On the other hand, a study suggests BsmI associated with a decreased risk of colorectal cancer [64]. We found that the VDR rs1544410 A allele is associated with the risk of aMCI in elderly. However, it is not clear that whether the VDR variant influences the composition of gut microbiota, and finally impair the cognitive function in elderly population. Therefore, in the present study, the association of VDR gene variant with the composition of gut microbiota in aMCI patients or NC population was investigated and the results indicated the microbial alpha and beta diversity were significantly different between individuals with AG and GG genotype in VDR among NC or aMCI subjects. The LEfSe analysis results indicated that the abundance of more than 19 bacterial taxa in fecal microbiota composition were significantly different between subjects with AG and GG genotype VDR in aMCI group. These altered abundances of bacterial taxa at genus level were positively or negatively correlated with the cognitive test scores in aMCI patients. Our results demonstrated that the VDR gene variants have an impact on the composition of gut microbiota and potentially link with the risk of aMCI in elderly population.
Alzheimer’s disease is a complex neurodegenerative disorder caused by a combination of genetic effects and environmental factors [65–67]. We have found that the VDR gene variants were associated with the gut microbiota and the altered bacterial taxa at genus level were correlated with cognitive test scores in aMCI patients, then we further evaluated the links of crosstalk between the genetic variants of VDR gene and the environmental factor altered GM with the risk of aMCI in the Chinese elderly population. For those subjects with AG genotype in VDR gene, the alpha and beta diversity showed significant differences between NC and aMCI group, and more than 23 bacterial taxa in fecal microbiota composition were confirmed to be the distinguished microbes in aMCI and NC group. The relative abundances of some genus were positively or negatively linked with the cognitive test scores and vitamin D levels. The metabolic pathways predicted by PICRUSt2 revealed that some bacterial metabolic products were involved in the pathological process of aMCI. Among the KEGG pathways level 3, the microbiome of aMCI patients with AG genotype demonstrated the most profound changes including reduction of glycine, serine, and threonine metabolism, phenazine biosynthesis, and protein processing in endoplasmic reticulum, and increased selenocompound metabolism, compared with NC with AG genotype. Studies have shown that the glycine, serine, and threonine metabolism pathways disrupted in the prodromal Parkinson’s disease and depression [68, 69]. Moreover, as an inhibitory neurotransmitter, glycine is crucial for controlling synaptic plasticity in the central nervous system [69], and cognitive impairment is associated with synaptic plasticity deficits [70]. It is worth noting that glycine, serine, and threonine metabolism pathways were upregulated in malignant disease [71]. It provides a clue for us to explain the negative correlation between tumor and dementia from the perspective of intestinal flora. This result indicated that the crosstalk between the VDR gene polymorphism and intestinal microbiota were linked with the risk of aMCI in elderly population.
There are some advantages and disadvantages in this study. The first advantage is that the changes in fecal microbiota were analyzed using full-length 16sRNA sequencing, which had a better resolution for distinguishing particular taxa, allowing accurate representation of bacterial composition in the sample [39]. Secondly, the serum levels of Aβ1-42 and p-tau (181P) were randomly detected to help screen the aMCI subjects. Thirdly, this study has established a link between the interaction of VDR variant and GM with the cognition impairment, which may contribute to better understand the roles of genetic variants and environmental factors in the pathogenesis of aMCI or AD. Nonetheless, this study also has several limitations. The first is that the levels of Aβ1-42 and p-tau (181P) in cerebrospinal fluid (CSF) were not determined in this study. Secondly, the numbers of subjects undergoing fecal microbiological analysis were relatively small. Thirdly, detection of a few tag SNPs in VDR/VDBP gene as potential risk factors may be relatively weak for aMCI. The larger cohort and longitudinal long-term studies are needed in future study.
In summary, we conducted a cross-sectional study and found the serum vitamin D levels were lower in aMCI patients, which potentially had diagnostic value for aMCI patients to some extent. The VDR gene polymorphism rs1544410 were associated with the gut microbiota and the altered bacterial taxa at genus level were correlated with cognitive test scores in aMCI patients. Particularly, the crosstalk between the VDR gene variants and intestinal microbiota were functionally linked with the risk of aMCI in elderly population. This study will provide evidence to better understand the impact of crosstalk between the genetic variants and environmental factors on the pathogenesis of aMCI or AD.
Footnotes
ACKNOWLEDGMENTS
This work was supported from the Hebei Provincial Natural Science Foundation (H2018206358, H2020206224 and H2020206105), The Science and Technology project of the People’s Livelihood in Hebei Province (20377707D), Special Funding for Local Science and Technology Development Guided by the Central Government (206Z7701G), Hebei Medical applicable technology tracking project (GZ20200061), the Key Program of traditional Chinese medicine in Hebei Province (Z2022015), and CAMS Innovation Fund for Medical Sciences (2019-I2M-5-055).