
Abstract
Background:
Amyloid-β peptide (Aβ) deposition in Alzheimer’s disease (AD) is due to an imbalance in its production/clearance rate. Aβ is transported across the blood-brain barrier by LRP1 and P-gp as efflux transporters and RAGE as influx transporter. Vitamin D deficit and polymorphisms of the vitamin D receptor (VDR) gene are associated with high prevalence of mild cognitive impairment (MCI) and AD. Further, vitamin D promotes the expression of LRP1 and P-gp in AD-animal model brains.
Objective:
To associate VDR polymorphisms Apa I (rs7975232), Taq I (rs731236), and Fok I (rs2228570) with the risk of developing MCI in a Chilean population, and to evaluate the relationship of these polymorphisms to the expression of VDR and Aβ-transporters in peripheral blood mononuclear cells (PBMCs).
Methods:
VDR polymorphisms Apa I, Taq I, and Fok I were determined in 128 healthy controls (HC) and 66 MCI patients. mRNA levels of VDR and Aβ-transporters were evaluated in subgroups by qPCR.
Results:
Alleles A of Apa I and C of Taq I were associated with a lower risk of MCI. HC with the Apa I AA genotype had higher mRNA levels of P-gp and LRP1, while the expression of VDR and RAGE were higher in MCI patients and HC. For Fok I, the TC genotype was associated with lower expression levels of Aβ-transporters in both groups.
Conclusion:
We propose that the response to vitamin D treatment will depend on VDR polymorphisms, being more efficient in carriers of protective alleles of Apa I polymorphism.
Keywords
INTRODUCTION
Mild cognitive impairment (MCI) is the transitional period between normal aging and dementia, without compromising the usual activities of the patient’s daily life [1, 2]. MCI is the best predictor of future dementia, including Alzheimer’s disease (AD) [2]. It has been estimated that MCI has an accelerated rate of progression to AD; in a follow-up year, the conversion rate was 10–30% (MCI to AD), compared to cognitively normal elders who tend to develop dementia at a rate of 1–2% per year (normal to AD) [2]. In 2011, the concept of MCI due to AD was defined to indicate the initial stages of disease [3]. In this period, abnormal deposits of proteins in the brain, such as amyloid-β (Aβ) and p-Tau, can also be detected in plasma and cerebrospinal fluid [4, 5].
The mainstay of the Aβ hypothesis of AD is that a gradual and chronic imbalance in the production, versus the clearance, of Aβ leads to a slow rise in its steady state levels in brain tissue [6, 7]. Recent results estimate that small deficits of clearance efficiency of Aβ from the brain of only to 2–5% could explain the difference of Aβ accumulation observed in AD brains over 20 years compared to controls [8]. The Aβ clearance pathway includes 1) enzymatic degradation [9, 10], 2) bulk flow through the cerebrospinal fluid (CSF) [11], 3) endocytosis by astrocytes and microglia cells [12], and 4) transport across the blood-brain barrier (BBB) [13].
The BBB that separates blood from interstitial fluid in the brain plays a central role in controlling brain homeostasis and regulating brain microenvironment. The soluble Aβ peptide can be exported from brain to the systemic circulation through transporters in the BBB that mediate the clearance of Aβ and vice versa. Efflux pathways occur through the low-density lipoprotein receptor–related protein-1 (LRP1) and glycoprotein P (P-gp). LRP-1 is located in the abluminal side of the endothelial cells of the BBB and binds to Aβ relocating it from the brain interstitium into the endothelial cell [14, 15]. LRP1 is also involved in Aβ endocytosis and transcytosis through the endothelial cells and its subsequent systemic elimination via the liver, spleen, and kidney [16]. In the luminal membrane of the BBB endothelial cells, P-gp works by transporting substrates to the blood. P-gp mediates Aβ clearance in an ApoE-dependent manner, but the precise mechanism is unknown [14, 17]. LRP-1 and P-gp seem to form an effective and complete transport system that coordinates Aβ transport and elimination from the brain. In contrast, the receptor for advanced glycation end products (RAGE) is located in the luminal side of BBB endothelial cells and is responsible of the influx pathway of Aβ. RAGE recovers free Aβ from the periphery and introduces it into the brain, contributing to brain Aβ aggregation [18–21].
Vitamin D plasma concentration has been evaluated in relation to several neurodegenerative diseases, such as MCI, AD, multiple sclerosis, and Parkinson’s disease (PD) [22–26]. Many studies have linked low levels of vitamin D with a high prevalence of AD, cognitive impairment, and low hippocampal volume in humans [27–30]. Vitamin D is considered one of the modifiable factors that can contribute to better management of AD [31, 32]. The genomic effect of vitamin D is mediated through its interaction with a high-affinity nuclear vitamin D receptor (VDR), a member of the nuclear steroid hormone receptor superfamily that regulates the expression of different genes in a ligand-dependent manner [33, 34]. The binding of vitamin D to the VDR allows a conformational change in VDR that forms either a homodimer or heterodimer with retinoic alpha receptor (RXRα). These complexes translocate to the nucleus and mediate the transcription of several genes that are involved in multiple functions like cell cycle activity, immune functions, DNA replication and stress response [35–37]. Transcriptome-wide analysis shows that 200 to 600 genes are primary targets of the vitamin D-VDR complex. This data is supported by the genome-wide view on VDR-binding sites by chromatin immunoprecipitation combined with high throughput sequencing (ChIP-seq) in mononuclear human cells [38, 39] and microarray analyzes [26, 41].
It has been demonstrated that 12-months supplementation of vitamin D improves cognitive function and decreases Aβ-related biomarkers in AD patients [42]. In a previous study, we also demonstrated that a 6-months supplementation of vitamin D to MCI patients was associated with an improved cognitive status on a follow up evaluation at 18 months. In addition, vitamin D treatment was accompanied by an increase in Aβ plasma levels [43], suggesting that vitamin D may improve Aβ efflux from the brain. Moreover, in vivo and in vitro studies have shown that vitamin D promotes the expression of LRP1 and P-gp, in correlation with a higher transport of Aβ peptide [44–47]. Interestingly, the multidrug-resistance protein 1 (MDR1) gene, which encodes the P-gp protein, is regulated by VDR [46]. The induction of MDR1 by vitamin D is mediated by VDR/RXR binding to several vitamin D response elements (VDREs) located upstream of the MDR1 gen [48], which partly contributes to explain the vitamin D response. Vitamin D treatment also enhances the expression of RAGE and promotes LRP1 expression in mice hippocampal endothelial cells [44]. Taken together, these findings indicate that vitamin D can modulate Aβ clearance, although the relationship of vitamin D and expression of Aβ transporters in human tissue has not yet been studied.
It is now accepted that single nucleotide polymorphisms (SNPs) in different genes may increase AD risk [49, 50]. SNPs located in the VDR gene have been associated with increased risk of MCI and AD acting by modulating the response to vitamin D [51–56]. Fok I (rs2228570) is a start codon polymorphism, where the change of T to C (ATG to ACG) causes initiation of translation at the second start codon, producing a shorter protein that lacks three N-terminal amino acids [57]. Apa I (rs7975232) is an A to C variation in intron 8, and Taq I (rs731236) is a C to T synonymous change in exon 9. Apa I and Taq I variations do not alter the amount, structure, or function of the VDR protein, but are strongly linked to a poly-A repeat that may affect mRNA stability [58, 59]. The poly-A repeats are located on 3’-untranslated regions (3’-UTR) and therefore the effects of these variants in VDR expression, structure, and function have been difficult to define. It is known that 3’-UTR is involved in expression, especially through the regulation of mRNA stability, including the steroid receptor [57]. Several studies have demonstrated an association between VDR polymorphisms and the risk of MCI and AD in European [51, 60] and other populations [52, 55]; however, there are no studies showing the association of these polymorphisms to MCI in Chilean population.
In the present work, we evaluated the association of the VDR polymorphisms Apa I, Taq I, and Fok I to the risk of developing MCI in Chilean population. Furthermore, we evaluated the relationship of these SNPs with the expression of VDR and the Aβ transporters P-gp, LRP1, and RAGE in peripheral blood mononuclear cells (PBMCs) obtained from MCI patients and healthy controls (HC) to understand the function of vitamin D and VDR in the clearance of Aβ and its possible role in AD pathogenesis.
METHODS
Subjects
One hundred and ninety-four individuals of both genders and over 60 years old were enrolled in this study: 128 HC and 66 MCI. All subjects signed a written informed consent and the study abides to the ethical guidelines of the 1975 Declaration of Helsinki. The study protocol was approved by the Ethics Committee of the Hospital Clínico de la Universidad de Chile (HCUCH). Some samples were obtained in the Biobank of tissues and fluids of the University of Chile.
MCI diagnosis was established following both the guidelines of the Alzheimer’s Association and the National Institute on Aging [61] and Petersen criteria [1]. The Clinical Dementia Rating (CDR) and Clinical Dementia Rating Sum of Boxes (CDR-SOB) were applied to determine dementia severity. All MCI patients had a global CDR score of 0.5 with CDR-SOB scores ranging from 0.5 to 2.5. The scores of HC were 0 for global CDR and CDR-SOB. The demographic data of the study participants are shown in Table 1.
Sociodemographic and clinical features of the study participants
HC, healthy controls; MCI, mild cognitive impairment; MoCA, Montreal Cognitive Assessment; MIS, Memory Index Score; MMSE, Mini-Mental State Examination.
All participants were evaluated with the Mini-Mental State Examination (MMSE) [62] and some of them with the Montreal Cognitive Assessment (MoCA) [63]. The MMSE has a maximal score of 30 with lower scores associated with more important cognitive deterioration. Traditionally, a 23/24 cut-off has been used to select patients with suspected cognitive impairment or dementia [64]; however, a 21/22 cut-off score was validated in Chilean population [65]. The MoCA test, validated in Spanish in our country [66], and the Montreal Cognitive Assessment Memory Index Score (MoCA-MIS) were applied to 65 HC and 38 MCI patients. The maximum score for MoCA is 30, with lower scores associated with more important cognitive deterioration. The score of the MoCA-MIS ranging from 0 to 15 was calculated by adding the number of words remembered in free delayed recall multiplied by 3, category-cued recall multiplied by 2, and multiple-choice-cued recall multiplied by 1 [67]. HC were submitted to the same neurological and neuropsychological evaluations.
The exclusion criteria for both groups were any medical diagnosis or judgment problems that prevented their participation. The presence of dementia, non-degenerative MCI, severe depression, or MCI of uncertain origin were also considered exclusion criteria for the MCI group.
VDR genotyping
Genomic DNA was extracted from peripheral blood collected in EDTA-containing Vacutainer™ tubes using DNA Genomic PureLink® Kit (Invitrogen) according to the manufacturer’s instructions. The polymorphisms genotyping were analyzed by real-time PCR using TaqMan® SNP Genotyping Assay (ThermoFisher). The probes for VDR polymorphisms Apa I (rs7975232), Taq I (rs731236) and Fok I (rs2228570) were designed and synthesized by Applied Biosystems (Assay ID: C__28977635_10, C__2404008_10, C__12060045_20) for Apa I, Taq I, and Fok I, respectively).
Peripheral blood mononuclear cells (PBMCs) isolation, RNA extraction, and cDNA synthesis
PBMCs were isolated from peripheral blood using Ficoll-Hypaque density centrifugation as previously described [43]. PBMCs were preserved in 1 ml TRIZol Reagent (Ambion) and stored at –80°C until mRNA extraction. Total RNA was extracted with TRIzol Reagent, according to the provider’s instructions. cDNA was synthesized from 1μg of RNA using High Capacity cDNA Reverse Transcription Kit (Thermo Fisher).
mRNA quantification
The mRNA levels of VDR, P-gp, LRP-1, and RAGE were quantified by real-time PCR using Master Mix SsoAdvanced™ Universal SYBR® Green Supermix (Bio-Rad) according to the manufacturer instructions. All results were normalized to the geometric mean of 18S rRNA and SDHA housekeeping genes. The mRNA levels were calculated using ΔCT method. Primer sequences for all genes are included in Table 2.
Primers sequences
Statistical analysis
All sociodemographic data were analyzed by the t-student test or Mann-Whitney test according to the distribution of data previously calculated with the Kolmorogov-Smirnov test (n > 50). Comparisons in mRNA levels for each gene were calculated by nonparametric Mann-Whitney test in agreement to the non-parametric distribution of data calculated by Shapiro-Wilk test (n < 50). The differences between genotypes were calculated with the Kruskal Wallis test and the post-hoc Dunn’s test, and for dominance models we used the Mann-Whitney test. All data are presented as median±interquartile range.
The Hardy-Weinberg equilibrium (HWE) and distribution of the genotypic and allelic frequencies were calculated by the chi-square (χ2) test. To evaluate the association of each SNP with MCI, we used the retrospective analysis odds ratio (OR). Statistical analysis was carried out in GraphPad Prism 8.0.2 software.
RESULTS
Genotypic and allelic frequencies
The distribution of alleles and genotypes frequencies of Apa I, Taq I, and Fok I VDR polymorphisms were evaluated in MCI patients and HC. Table 3 shows the distribution of the Apa I polymorphism (A > C); the analysis of the genotypic frequencies was different between MCI patients and HC (p = 0.020). We found that the AA frequency was lower in MCI patients (17% versus 36%, MCI and HC respectively), meanwhile the AC (48% versus 38% MCI and HC, respectively) and CC (35% versus 27% MCI and HC, respectively) frequencies were higher in MCI patients.
Distribution of genotypic and allelic frequencies of VDR polymorphism Apa I
HC, healthy controls; MCI, mild cognitive impairment; OR, odds ratio; HWE, Hardy-Weinberg equilibrium.
The analysis of the allele frequencies was also different between MCI patients and HC (p = 0.01). Our results show that the C allele was more frequent in MCI patients (59% versus 45%, MCI and HC respectively), whereas the A allele was less frequent in MCI patients (41% versus 55%, MCI and HC, respectively). Furthermore, the OR showed that carrying the C allele could be a risk factor to develop MCI (OR = 1.74; IC95% 1.14 to 2.68). By contrast and concordant with a higher frequency in HC, the A allele might reduce the risk of MCI (OR = 0.57, IC95% 0.37 to 0.87). However, these results must be taken with caution because the HC groups were not in Hardy-Weinberg Equilibrium (HWE).
The distribution of genotypic and allelic frequencies of the Taq I polymorphism (T > C) are presented in Table 4. The analysis of the genotypic frequencies of the Taq I was different between MCI patients and HC (p = 0.043). We found that the TT (48% versus 41%, MCI and HC, respectively) and TC (47% versus 41%, MCI and HC, respectively) genotype frequencies were higher in MCI patients; meanwhile the CC frequency was lower in MCI patients than HC (5% versus 17%, MCI and HC, respectively). Only three individuals were carriers of the CC genotype in MCI patients. Regarding the allelic frequency, we found that the T allele was higher in MCI patients (72% versus 62%, MCI and HC, respectively), suggesting that carrying the T allele might be a risk to developed MCI. Meanwhile, the C allele was less frequent in MCI patients (28% versus 38%, MCI and HC, respectively). The C allele could be a protective factor against MCI, and this could increase with two copies of C allele (OR = 0.63, IC95% 0.40 to 0.99).
Distribution of genotypic and allelic frequencies of VDR polymorphism Taq I
HC, healthy controls; MCI, mild cognitive impairment; OR, odds ratio; HWE, Hardy-Weinberg equilibrium.
We found no differences in the distribution of either the genotypes or alleles in the Fok I polymorphism (T > C) (Table 5), in accordance with the evidence that this SNP is more related to PD [68]. Both Taq I and Fok I were in HWE in HC and MCI.
Distribution of genotypic and allelic frequencies of VDR polymorphism Fok I
HC, healthy controls; MCI, mild cognitive impairment; OR, odds ratio; HWE, Hardy-Weinberg equilibrium.
Decreased mRNA levels of VDR and its target genes in HC and MCI patients carrying the Apa I risk allele
To evaluate the effect of VDR polymorphisms on the expression of VDR and its target genes, the Aβ transporters P-gp, LRP1, and RAGE, we determined their mRNA levels in PBMCs in a subgroup of 29 HC and 29 MCI patients with vitamin D insufficiency [69] (Supplementary Table 1). Our results show that the mRNA expression of VDR and its targets genes are similar in HC and MCI patients (Supplementary Figure 1). Therefore, we further analyzed the relationship between genotypes and mRNA levels. First, we analyzed the relationship between Apa I genotypes and mRNA levels. We found a decrease of VDR mRNA levels in MCI patients carrying the AC genotype, but not in HC (Fig. 1A, B). Since the C allele could be a risk allele (Table 3), we hypothesized that this allele may be acting in a dominant model in relation to the allele A [70]. Thus, we grouped individuals into AA and C carriers (AC/CC) from each group. When we applied this dominance model, we found that the mRNA expression of VDR was higher in AA genotype than AC/CC in both groups (Fig. 1C, D). Therefore, these results imply that the C allele is associated with a lower mRNA expression of VDR.

mRNA levels of VDR in association to Apa I genotype in PBMCs from healthy controls (HC) and mild cognitive impairment (MCI) patients. Expression levels were separated by their genotype for HC (A) and MCI patients (B). A dominant model of aggrupation was applied, where the C allele is a risk allele for MCI. The genotypes were grouped as AA and AC/CC in HC (C) and MCI patients (D). Each point represents a patient. Statistics analysis: for (A, B) Kruskal Wallis test and post hoc Dunn’s test, for (C, D) Mann-Whitney’s test. *p < 0.05, **p < 0.005.
We applied the same dominant model to evaluate the mRNA levels of the VDR target genes, P-gp, LRP1 and RAGE. We found that the AA genotype had higher mRNA levels of P-gp (Fig. 2A) and LRP1 (Fig. 2C) in HC; meanwhile this was not observed in MCI patients where the P-gp and LRP1 levels were not different between AA and AC/CC genotypes (Fig. 2B, D). For RAGE, we found that the AA genotype had higher mRNA levels than AC/CC in both HC (Fig. 2E) and MCI patients (Fig. 2F). Taken together, these results suggest that the presence of the C allele decreases the expression of VDR and Aβ transporters in PBMCs from HC, whereas in MCI patients, differences were observed for RAGE only.

mRNA levels of Aβ transporters in association to Apa I genotype in PBMCs from healthy controls (HC) and mild cognitive impairment (MCI) patients. Expression levels are expressed by dominant model, where the genotypes were grouped as AA and AC/CC. Relative expression of mRNA P-gp in HC (A) and MCI patients (B). Relative expression of mRNA LRP-1 in HC (C) and MCI patients (D). Relative expression of mRNA RAGE in HC (E) and MCI patients (F). Each point represents a patient. Statistics analysis: Mann-Whitney’s test. *p < 0.05.
Although we found differences of genotypic and allelic frequencies of Taq I (Table 4), we found no differences in mRNA expressions of VDR (Supplementary Figure 2) or Aβ transporters (Supplementary Figure 3).
Decreased mRNA levels of VDR and its target genes in HC and MCI patients carrying the Fok I TC genotype
Although the distribution of genotypes and alleles of Fok I polymorphism were not different between MCI patients and HC (Table 5), we also evaluated the mRNA expression of VDR and Aβ transporters in relation to Fok I genotype. Since we could not define a risk allele for Fok I in our population, the dominant model was not applicable.
Figure 3 shows that the mRNA expression of VDR was lower in the heterozygous genotype (TC) compared to the homozygous TT in HC, whereas no differences between genotypes were found in MCI patients (Fig. 3B). Regarding the expression of the Aβ transporters, P-gp expression was decreased in TC genotype in MCI patients compared to CC genotype (Fig. 4B), unlike HC where no differences between genotypes was observed (Fig. 4A). On the contrary, LRP1 was lower expression in TC genotype from the HC (Fig. 4C), without differences in the MCI patients (Fig. 4D). For RAGE, we observed a decrease in TC genotype in relation to TT genotype, in both groups (Fig. 4E, F).
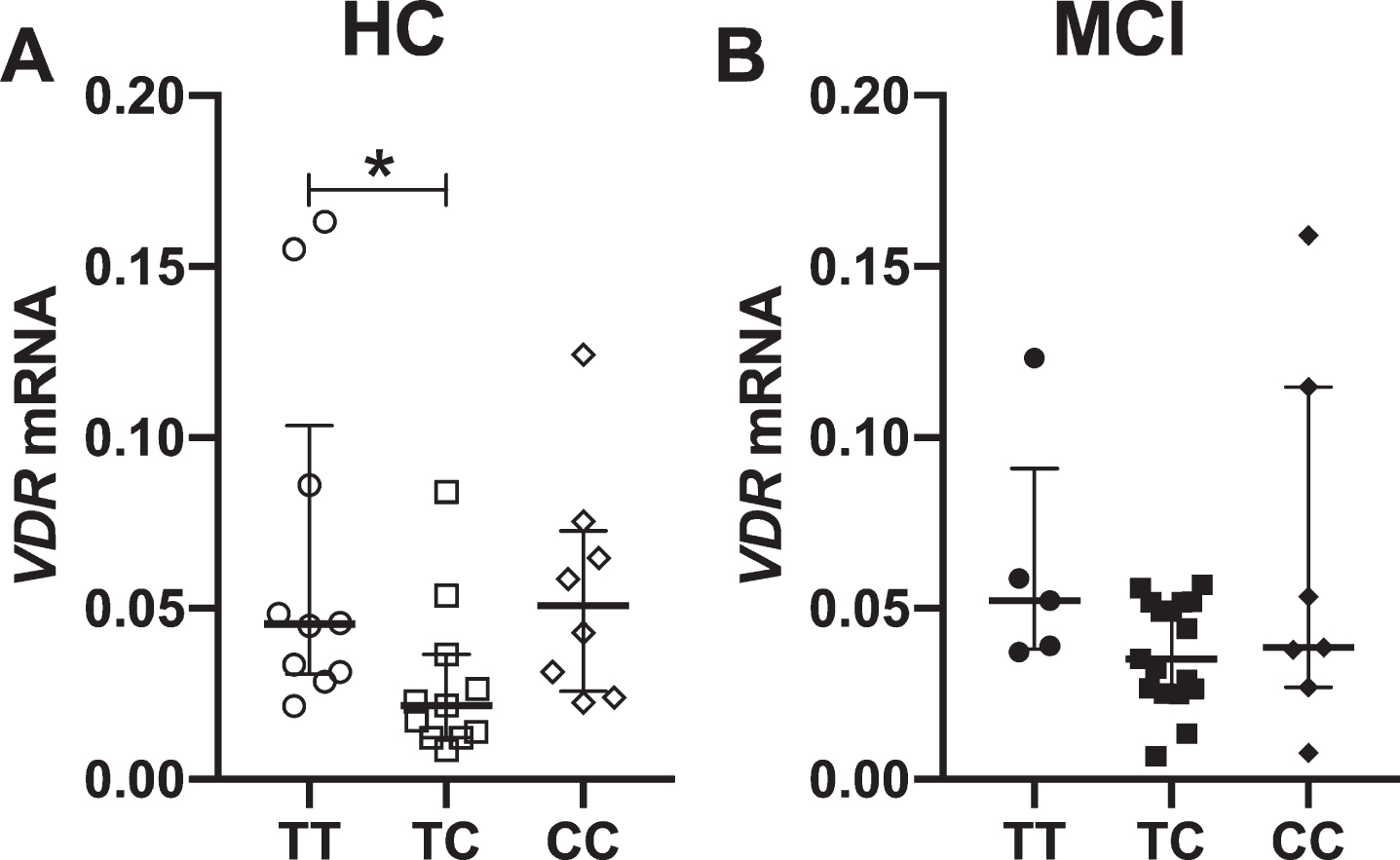
mRNA levels of VDR in association to Fok I genotype in PBMCs from healthy controls (HC) and mild cognitive impairment (MCI) patients. Expression levels were separated by their genotype for HC (A) and MCI patients (B). Statistics analysis: Kruskal-Wallis test and post hoc Dunn’s test. *p < 0.05.
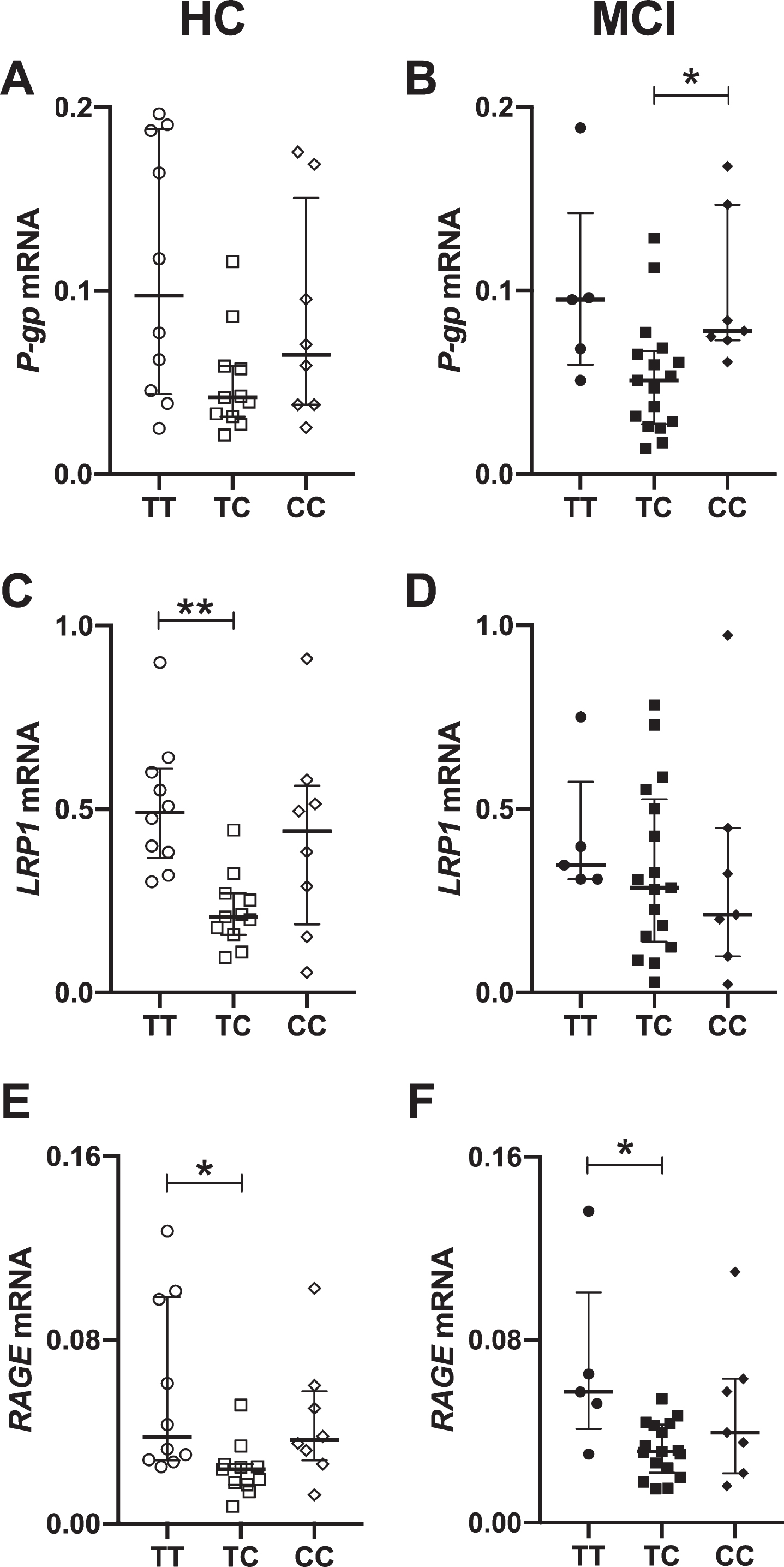
mRNA levels of Aβ transporters in association to Fok I genotype in PBMCs from healthy controls (HC) and mild cognitive impairment (MCI) patients. Relative expression of mRNA P-gp in HC (A) and MCI (B) patients. Relative expression of mRNA LRP-1 in HC (C) and MCI patients (D). Relative expression of mRNA RAGE in HC (E) and MCI patients (F). Each point represents a patient. Statistics analysis: Kruskal-Wallis test and post hoc Dunn’s test. *p < 0.05, **p > 0.01.
DISCUSSION
In this report we show for the first time the association of VDR SNPs with MCI in a Chilean population. Our findings suggest that the A allele of Apa I is associated with a lower risk of developing MCI (Table 3, Fig. 5A). Control individuals with the AA genotype had higher levels of mRNA expression of VDR (Fig. 1), P-gp and LRP1 (Fig. 2). The mRNA expression of VDR (Fig. 1) and RAGE (Fig. 2) were higher in both HC and MCI patients with AA genotype. We also found that the C allele of Taq I decreases the risk of developing MCI (Table 4, Fig. 5A); however, we found no differences in mRNA expressions of VDR or Aβ transporters (Supplementary Figures 2 and 3). Finally, although we did not find a risk or protective allele for MCI in the Fok I SNP (Table 5), the expression of VDR mRNA was lower in TC genotype in HC (Fig. 3), and we also found lower expression levels of Aβ transporters in these heterozygous individuals (Fig. 4).
Our findings suggest that the A allele of Apa I and C allele of Taq I are associated with a lower risk of MCI. Reports associating these SNPs in other populations found contrasting results. While the A allele of Apa I was more frequent in the control group than AD patients in a Low Silesian population [60], the AA genotype of Apa I was a risk factor for MCI in the Uygur people [55]. Another study suggested that AC genotype could be a risk factor for the development of AD in Turkish population [54]. The T allele of Taq I was higher in AD patients in Turkish and Iranian population [60]. Moreover, a haplotype constituted by the T allele of the Taq I and C allele of Apa I represents a risk factor for AD in Turkish population [71]. Differences between populations could be attributed to the genetic structure among populations with differences in linkage disequilibrium with other SNPs, different genetic background with epistatic genes and other factors that might predispose the development of the disease. Moreover, latitude, genetic traits, age, gender, diet, living conditions, levels of outdoor activity, and cultural behaviors are essential factors in determining vitamin D levels and VDR regulation [72, 73].
As reported in other populations we found no association of the Fok I SNP with MCI in the Chilean population. This SNP has been associated mostly with PD in carriers of the C allele [74–77]. Nevertheless, other researchers propose that the T allele is associated with a higher risk of PD [51, 78].
It has been proposed that an imbalance in the production versus the clearance of Aβ leads to a rise in its steady state levels in brain tissue [6, 7]. We propose that a decrease in Aβ transporters responsible for Aβ clearance through the BBB might explain the association between VDR SNPs and AD risk. To relate the VDR SNPs with the expression of VDR and some of the VDR-regulated genes relevant for Aβ clearance, we evaluated gene expression of Aβ transporters P-gp, LRP1, and RAGE in PBMCs from MCI patients and HC.
Since we found that the C allele for Apa I may be a risk allele to MCI, we established a dominance model for this allele [70]. We found that carriers of the AA genotype had higher levels of mRNA expression of VDR than carriers of one or two copies of the risk C allele (AC/CC), in HC and MCI patients (Figs. 1 and 5B). HC carriers of AA genotypes also had higher levels of Aβ transporters P-gp and LRP1 (Figs. 2 and 5B). Apa I is located near the untranslated region 3’-UTR and therefore could affect the expression, structure, function and stability of VDR mRNA. Likewise, it has been shown that this SNP is in linkage disequilibrium with a poly-A repeat in the 3’-UTR, that also affects mRNA stability [59, 79–82]. Due to the above and according to our results, we propose that the presence of the risk C allele affects the stability of VDR and its target genes mRNA. Our results suggest that Aβ clearance could be affected by genetic variation of VDR gen, and that the presence of the protective alleles will be associated with a better performance of Aβ clearance, explaining the association between VDR SNPs and MCI or AD (Fig. 5C). However, our results show that VDR-regulated genes are differentially modulated in MCI patients and HC and that the protective effect of the A allele seems to get lost in the context of the disease, according to the loss of cell homeostasis given in AD. However, it is necessary to evaluate the protein levels of Aβ transporters in endothelial cells of the BBB.
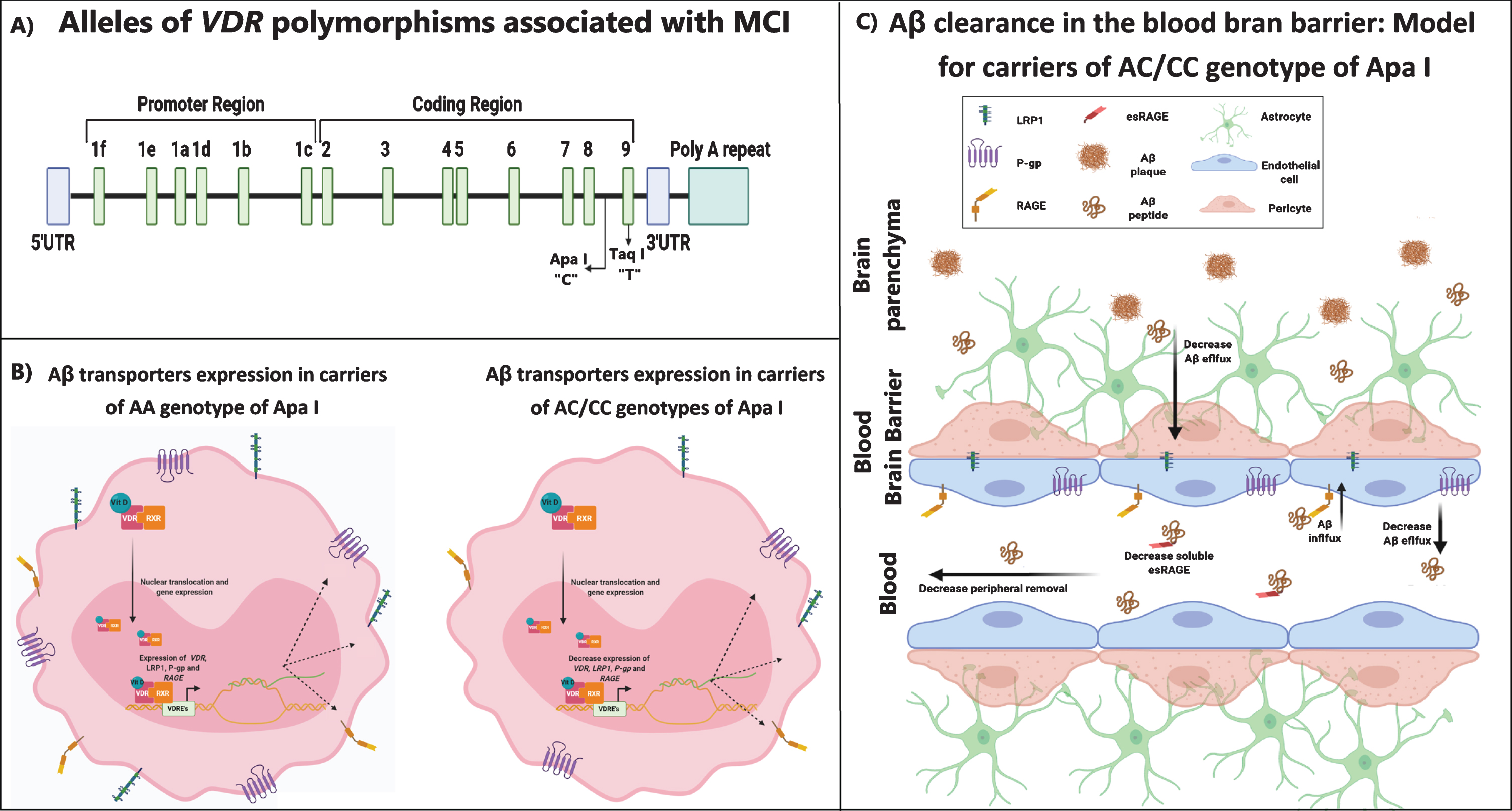
Proposed model of VDR polymorphism in PBMCs and BBB. A) Apa I and Taq I polymorphism location in the VDR gen. The C and T alleles for Apa I and Taq I, respectively, are associated with MCI in Chilean population. B) PBMCs expression model of VDR and Aβ transporters in carriers of AA genotype of Apa I. In AA carriers the VDR expression is increased, promoting the formation of the vitamin D-VDR-RXR complex. In this context P-gp, LRP-1 and RAGE expressions are promoted due to the presence of the vitamin D regulatory element (VDREs) in P-gp gene and probably in LRP-1 and RAGE genes. In AC/CC carriers the VDR expression is decreased, leading to lower formation of vitamin D-VDR-RXR complex. These lower VDR levels are associated with a decrease in P-gp, LRP-1 and RAGE expressions. C) The BBB mediates the clearance of soluble Aβ peptide from the brain to the systemic circulation and vice versa through efflux and influx Aβ transporters. The efflux pathways occur through the P-gp and LRP1 located in the abluminal side of the endothelial cells. The influx pathway is mediated by RAGE located in the luminal side of endothelial cells. RAGE recovers free Aβ from the periphery blood and introduces it into the brain, contributing to brain Aβ aggregation. Meanwhile, the endogenous soluble RAGE isoform (esRAGE) binds to free Aβ in the peripheral circulation, and prevents its interaction with the RAGE transporter, decreasing the Aβ aggregation. Based on our results in PBMCs, we propose a BBB model for AC/CC carriers. In this model the VDR expression was decreased, affecting the expression of P-gp, LRP-1 and probably also esRAGE. The decrease in Aβ transporters expression might have an impact on Aβ clearance, decreasing the Aβ efflux and increasing influx.
On the other hand, carriers of the A allele had higher levels of Aβ transporter RAGE, and this result was unexpected due to its responsibility on the influx pathway of Aβ [18–20]. RAGE has three isoforms; one isoform is located in the endothelial cell of BBB and participates in the influx of Aβ, the other two are non-membrane bound isoforms, produced by alternative splicing of RAGE gene called endogenous soluble RAGE isoform (esRAGE) which is a RAGE antagonist [19, 83–85]. esRAGE binds to free Aβ in the peripheral circulation, and prevents its interaction with the RAGE transporter and the subsequent internalization to the brain [83–85]. In AD and MCI patients, the levels of esRAGE are lower than in healthy controls [86, 87]. In our research, we cannot discriminate between the three RAGE isoforms; therefore it cannot be determined whether the increase in mRNA observed in individuals with protective alleles corresponds to RAGE or esRAGE.
Despite the association of Taq I SNP with MCI in our population (Table 4), we did not find associations with VDR or VDR-regulated genes expression (Supplementary Figures 2 and 3), in spite of some studies suggesting that the T allele is related to higher levels of VDR mRNA and protein [88, 89]. Taq I is in linkage disequilibrium with others SNPs like Apa I and Bsm I [53, 91]; hence it might be interesting to evaluate the relationship of Taq I with other VDR SNPs.
Although we did not find association between Fok I SNP and MCI in our population, the expression of VDR and Aβ transporters was lower in TC genotype in HC. It is known that Fok I causes a change in protein length [92]. The C allele interacts more efficiently with the transcription factor TFIIB and shows relatively increased VDR function compared to the T allele [92]. In this context, several studies have evaluated the levels of expression and transcriptional activity of the VDR in Fok I genotypes; and the results are controversial [80, 93–95].
To the best of our knowledge, there are no studies exploring the relationship of VDR SNPs and MCI or AD in the Latin American population, although several VDR SNPs have been studied in other pathologies. There is a correlation of Apa I to the prevalence of low bone mineral density in Turner syndrome patients [96], and Taq I with primary osteoarthritis [97], both in Mexican population. Brazilian studies have associated VDR SNPs to maternal and neonatal vitamin D levels and pregnancy outcome [98], meanwhile Bsm I and Taq I are associated with polycystic ovary syndrome [99]. In the Chilean population, Bsm I SNP was associated to a higher incidence of type I diabetes mellitus in children [100], and Fok I together with Bsm I were correlated to type II diabetes mellitus in adult patients [101, 102].
The principal limitation in our study was that the HC group was not in HWE for Apa I. We believe this is due to an unknown biological cause specific for Apa I. Our study population is homogeneous with the same ethnic characteristic, sampling and the same genotyping conditions and tools, so we rule out that it is due to poor stratification or genotyping error. We consider the HWE will be reached if the number of individuals is increased.
In conclusion, Latin America and the Caribbean show a considerable prevalence of vitamin D insufficiency in different age groups [103], being the elderly the most affected [104, 105]. Vitamin D deficiency has been linked to reduced cognitive response and memory loss, linking this deficiency to neurocognitive disorders like AD and MCI. In animal AD models, vitamin D supplementation has been shown to lower Aβ accumulation in the brain, decrease plaque burden and improve basal synaptic transmission, cognitive function, adult hippocampal neurogenesis, neurotransmitters synthesis and cerebral clearance of Aβ1–40 from the brain across the BBB [47, 106–108]. It has been demonstrated that 12 months supplementation of vitamin D improve cognitive function and decrease Aβ-related biomarkers in AD patients [42]. In a previous study, we demonstrated that a 6-months period of vitamin D supplementation to MCI patients was associated with an increase in Aβ1–40 plasma levels (reflecting an increased efflux from the brain). In addition, it was accompanied by an improved cognitive status on a follow up evaluation at 18 months [43]. According to this, vitamin D supplementation has proven to be an efficient coadjutant therapy in AD patients [109], however we propose that the response to vitamin D treatment will depend on VDR SNPs, being more efficient in carriers of protective allele for Apa I. Since vitamin D supplementation increases Aβ plasma levels in MCI patient [43], it is of great interest to evaluate the effect in the Aβ transporters in relation to the VDR genotype.
Footnotes
ACKNOWLEDGMENTS
We would like to acknowledge all the patients and their families for their participation in this research. We appreciate the work of the staff of the Biobank of tissues and fluids of the University of Chile. This work was supported by the grants: “Fondo Nacional de Desarrollo Científico y Tecnológico, Fondecyt” from Chile. Grant numbers: 11171061 (CS) and 1190958 (MB).