
Abstract
Using an amyloid precursor protein (App) gene knock-in (KI) mouse of Alzheimer’s disease (AD), we investigated the expression of olfactory-related genes in olfactory impairment caused by AD. We observed the change in olfactory behavior in the App-KI mice. There was no significant difference, however, in the mRNA expression levels of olfactory-related genes between the olfactory epithelia of wild-type (WT) and App-KI mice. Amyloid-β deposition was confirmed throughout the olfactory pathway in App-KI mice, but not in WT mice. These show that the change in olfactory behavior in the App-KI mice might cause by the impairment of the olfactory pathway.
INTRODUCTION
Tasting food is an important part of our daily lives, and in particular, smell plays an important role in tasting food (e.g., olfactory-driven food choices and the detection of spoiled food before it enters the mouth). Therefore, a decrease in olfactory function reduces the enjoyment of eating. Unfortunately, various illnesses, such as Alzheimer’s disease (AD), often induce olfactory dysfunction. AD, the major form of dementia, is a syndrome caused by numerous progressive illnesses that affect memory and cognitive capacity, and many studies have reported olfactory impairment in patients with AD [1]. In one comparative study between healthy subjects and patients with AD, it was reported that some olfactory functions, such as odor detection threshold, discrimination, and identification, were significantly lower in patients with AD than in healthy controls [2]. Although several studies have been conducted to clarify these pathogenic mechanisms, details about their processes remain controversial.
In this study, to estimate the detailed mechanism of olfactory dysfunction in AD, we used knock-in (KI) mouse models of AD that endogenously overproduce amyloid-β (Aβ) without non-physiological overexpression of amyloid precursor protein (APP) and measured their olfactory sensitivity. In addition, to investigate whether changes in the expression levels of olfactory-related genes caused olfactory impairment or not, we evaluated those expressions in the olfactory epithelium (OE), which contributes to the detection of odorants.
METHODS
Animals
AppNL - G - F mice harboring three familial AD mutations (Swedish, Iberian, and Arctic) were used as an App-KI model of AD. The original line of App-KI mice, established as the C57BL/6J (B6) congenic line (a genetic background strain) by repeated backcrosses [3], was provided by the RIKEN BRC through the National Bio-Resource Project of the MEXT/AMED, Japan. B6 mice, purchased from CLEA Japan, Inc. (Tokyo, Japan), were used as wild-type (WT) mice. Male mice were used for each experiment. For genotyping, genomic DNA was extracted from the biopsied mouse tails. The following PCR primers were used for genotyping: for WT (product size: 700 bp): 5′-ATCTCGGAAGTGAAGATG-3′ and 5′-TGTAGATGAGAACTTAAC-3′; for KI (product size: 400 bp): 5′-ATCTCGGAAGTGAATCTA-3′ and 5′-CGTATAATGTATGCTATACGAAG-3′. The PCR reaction was performed using the following protocols: 94°C for 2 min, 30 cycles of 98°C for 10 s, 55°C for 30 s, and 68°C for 60 s [3].
The temperature and humidity of the room in which the mice were housed were maintained at 24°C and 55%, respectively, with a 12-h light/dark cycle (lights were switched on at 0700 h). The mice were housed at the CLEA Japan Animal Care Facility until they were 18 months of age, following which they were housed at the Kyoto Women’s University Animal Care Facility during the experimental period. All experiments were performed in accordance with protocols approved by the Kyoto Women’s University Animal Care Committee (approval number: 2020-4).
Olfactory behavior test
The olfactory behavior test, for which 18–21-month-old WT (n = 14) and App-KI mice (n = 12) were used, was based on Witt’s report [4]. To habituate to the experimental environment, the mice were individually placed in cages identical to the test cages (28W×44D×20.5H cm). After a 15-min habituation period, the mice were transferred to a new cage. This habituation was repeated three times for each animal. Shortly after the third habituation period, the mice were transferred to the test cage, and a filter paper (5×5 cm) was transfused with solvent (distilled water). After 1 min interval, a filter paper scented with a test odorant was introduced into the test cage. The exploration times for the filter paper were measured during the 3-min test period, and the total time spent exploring the filter paper was assessed, with the total exploratory time calculated as exploratory time (scent)-exploratory time (water). The animals’ behavior was recorded using a commercial digital video camera. The odorants, which were measured on different days, were 30% vanilla extract (McCormick & Co., Hunt Valley, MD, USA) and 10% 2-methylbutyric acid (2-ME; Sigma-Aldrich, St. Louis, MO, USA).
Immunohistochemistry
Immunohistochemistry was performed as described previously reported [5]. The brain and OE from WT and App-KI mice were prepared by treatment with 4% paraformaldehyde in phosphate-buffered saline. The brain and OE were sliced at 50 and 14-μm thickness, respectively. Fluorescent immunostaining using mouse anti-amyloid-β (Aβ) 82E1 (1 : 200; IBL, Gunma, Japan) and rabbit anti-glial fibrillary acidic protein (GFAP) (1 : 500; SHIMA Laboratories, Tokyo, Japan) antibodies for the brain, and goat anti-OMP (1 : 500, Fijifilm Wako Pure Chemicals, Osaka, Japan) for the OE were performed. Additional details of the procedure are presented in the Supplementary Methods.
Real-time PCR
Real-time PCR was performed as previously described [6–9]. Briefly, the OE region was separated from the WT and App-KI mice. Total RNA from the OE was extracted using RNeasy Mini Columns (Qiagen, Venlo, Netherlands). Total RNAs of both strain mice were extracted from 6 samples randomly selected from the collected samples. Genomic DNA digestion was performed with an RNase-free DNase I Set (Qiagen). First-strand cDNA was generated from total RNA by reverse transcription (Superscript IV Reverse Transcription Kit; Life Technologies, Gaithersburg, MD, USA). The mRNA transcript levels were determined by real-time RT-PCR (QuantStudio 1; Thermo Fisher Scientifics). PCR amplification was performed using SYBR Green PCR system (Thermo Fisher Scientifics). PCR primers for SYBR Green PCR system are described in Supplementary Table 1. The delta-delta method was used for relative quantification [10]. Since the epithelial tissue surrounding the olfactory cells was present in the OE samples, the mRNA expression of Omp, a mature olfactory receptor neuron (ORN) marker, was used as an internal control to analyze the molecules associated with olfactory detection.
Statistical analysis
The results are expressed as mean±standard error of the mean. All statistical analyses were performed using GraphPad Prism 6 software (GraphPad Software, San Diego, CA, USA). The significant differences were evaluated using Welch’s t-test. For all analyses, statistical significance was set at p < 0.05.
RESULTS
To evaluate olfactory changes associated with AD, we first measured changes in olfactory behavior to attractive (vanilla extract) and aversive (2-ME) odors in 18–21-month-old mice. App-KI mice showed preference and avoidance behaviors toward vanilla extract and 2-ME, respectively, similar to that was seen in WT mice. Although it was confirmed that even App-KI mice exactly distinguish the odor qualities between attractive and aversive odors, we also observed the change in olfactory behavior to vanilla extract in App-KI mice (Fig. 1). The total exploratory time to vanilla extract in App-KI mice was significantly shorter than that in WT mice (p < 0.05), whereas there was no significant difference in the total exploratory time to 2-ME between WT and App-KI mice (Supplementary Table 2).
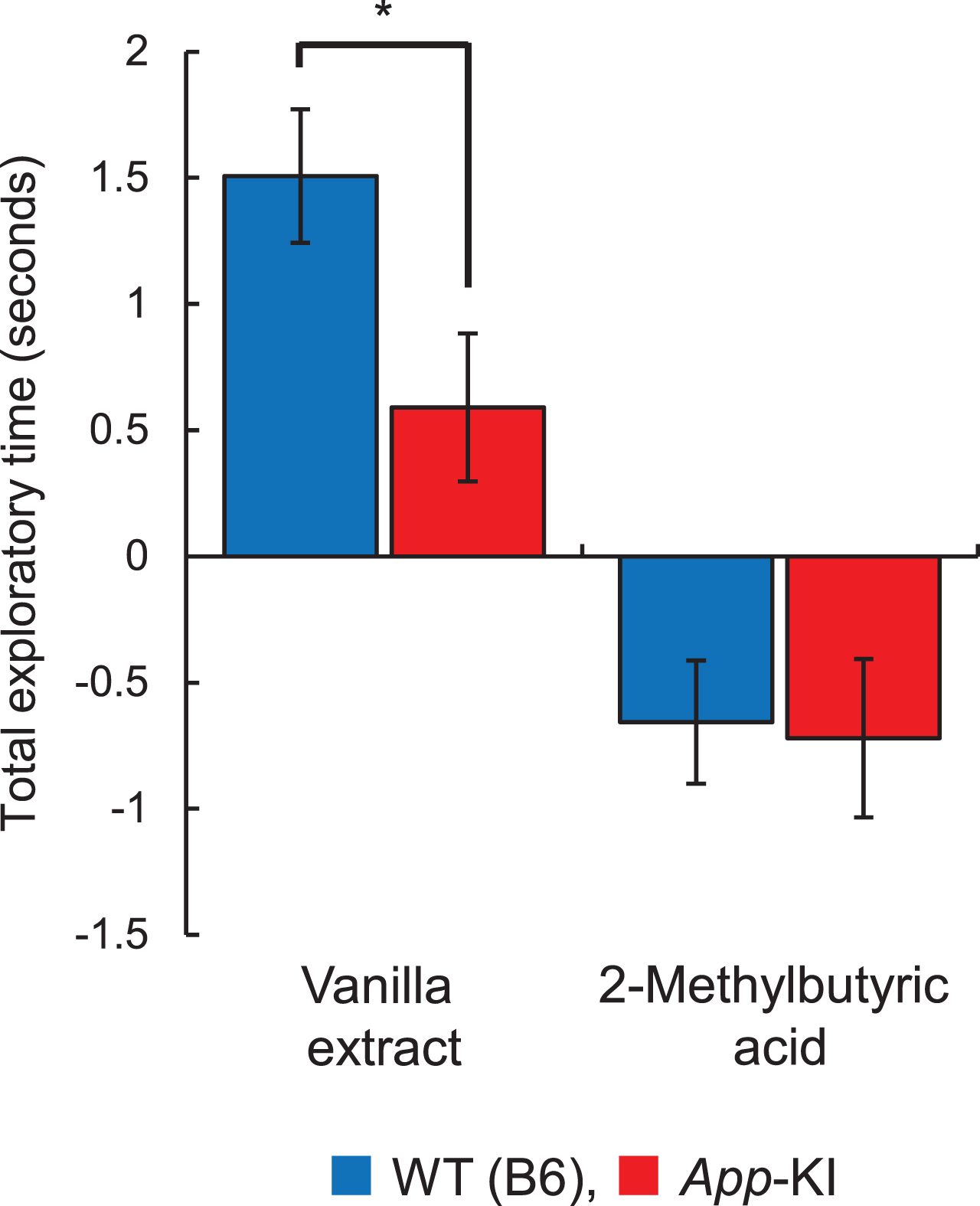
Change of olfactory preference of App knock-in (KI) mice. The total exploratory time for vanilla extract and 2-methylbutyric acid is shown. Blue and red columns indicate wild-type (WT) (n = 14) and App-KI (n = 12) mice, respectively. *p < 0.05 (Welch’s t-test).
Next, to investigate the cause of the change in olfactory behavior, we observed the expression of olfactory-related molecules in the OE. The expression of OMP, a molecular marker of an ORN, was confirmed in the OE of WT and App-KI mice (Fig. 2A). There was no obvious difference in the OMP expression patterns between the two genotypes. Although the mRNA expression levels of the olfactory-related molecules Olfr15, Golf, Cnga2, Cnga4, Cngb1, Tmem16B, Taar2, Taar3, and Taar7f were compared between WT and App-KI mice, no significant differences were observed (Fig. 2B and Supplementary Table 2).
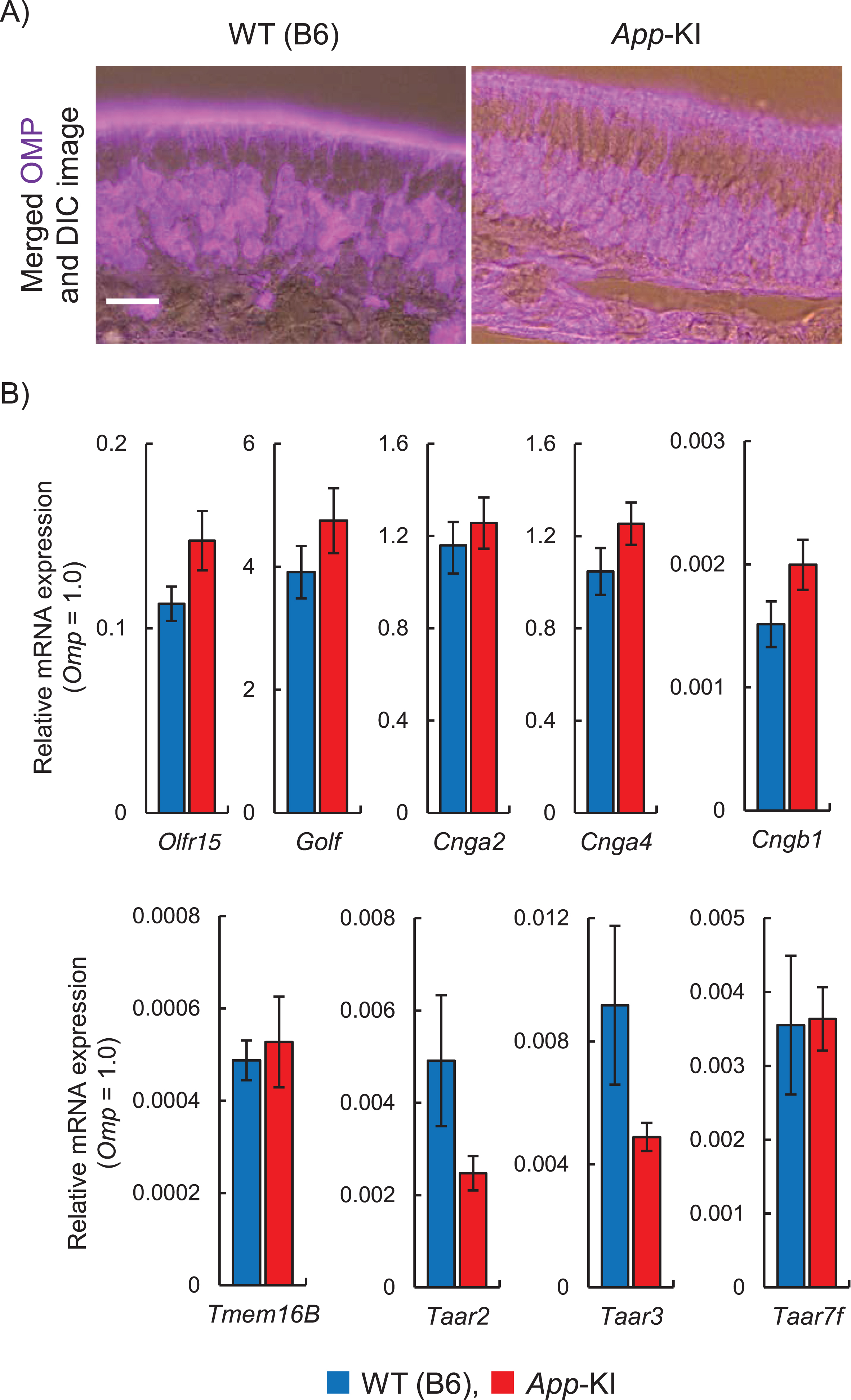
Expression of olfactory-related genes in the olfactory epithelium (OE) of App knock-in (KI) mice. A) Representative images of olfactory marker protein (OMP; magenta) expression in the OE. The images with merged fluorescence and differential interference contrast (DIC) images are shown. Scale bar is 20μm. B) Relative mRNA expression levels of Olfr15, Golf, Cnga2, Cnga4, Cngb1, Tmem16B, Taar2, Taar3, and Taar7f in the OE. Omp mRNA was used as an internal control (n = 6; Welch’s t-test).
Furthermore, we investigated Aβ deposition in the olfactory pathway, including the olfactory bulb, anterior olfactory nucleus, olfactory tubercle, piriform cortex, amygdala, and entorhinal cortex. In addition to Aβ deposition in the hippocampus (Fig. 3A), Aβ deposition was confirmed throughout the olfactory pathway in App-KI mice (Fig. 3B).
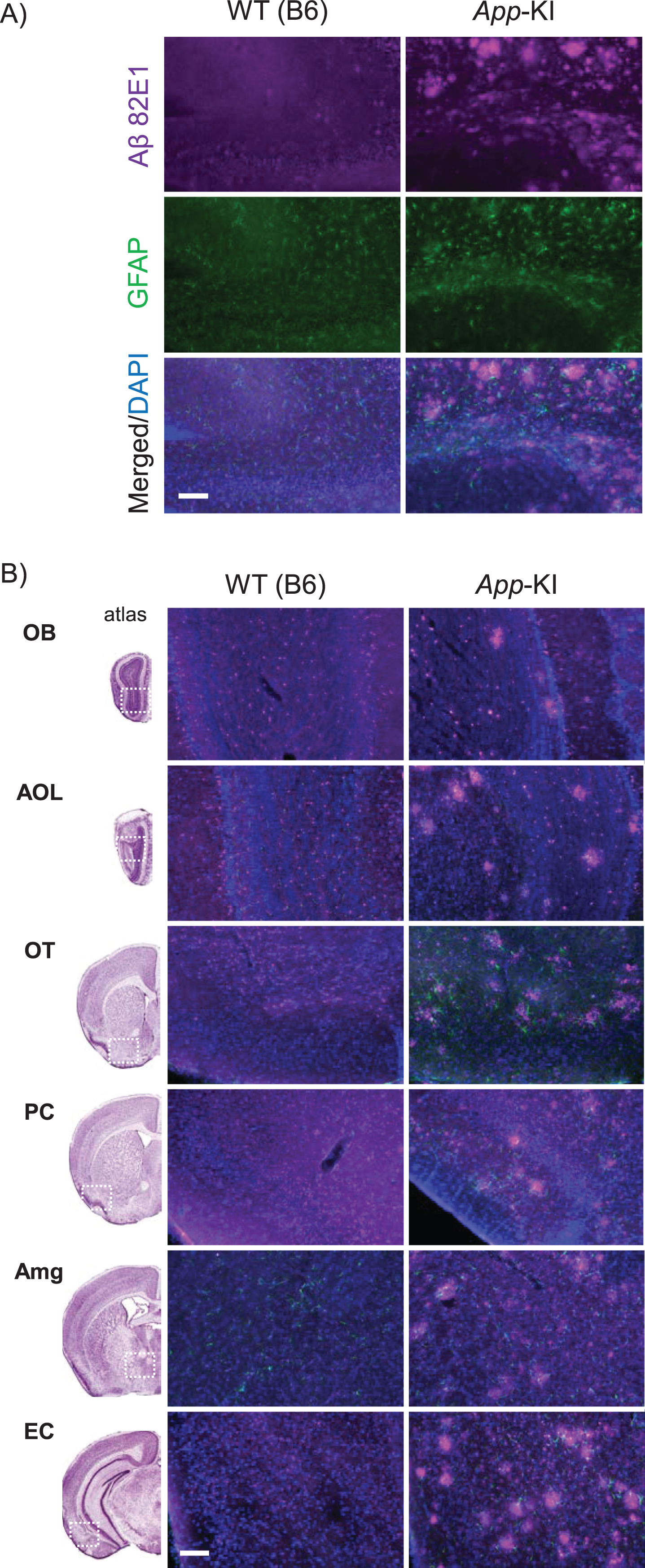
Amyloid-β (Aβ) deposition of wild-type (WT) and App knock-in (KI) mice. A) Representative images of the hippocampal region from coronal brain sections immunostained with anti-Aβ 82E1 (magenta) and anti-glial fibrillary acidic protein (GFAP) (green) are shown (blue in merged images indicates DAPI staining). B) Representative images of the olfactory pathway, including the olfactory bulb (OB), anterior olfactory nucleus (AOL), olfactory tubercle (OT), piriform cortex (PC), amygdala (Amg), and entorhinal cortex (EC), merged with Aβ 82E1, GFAP, and DAPI images are shown. The left column shows reference images, which include each brain area (refer to The Mouse Brain [22]). Scale bar is 100μm.
It has been generally accepted that the sense of smell deteriorates in patients with neurodegenerative disorders, including AD, and a decreased ability to smell is associated with the early stages of the disease. It is still unclear, however, which neural substrates are responsible for pathophysiological changes in the olfactory system caused by AD.
We started the behavioral experiment in mice after 18 months of age. Since it has been reported that Aβ deposition and cognitive impairment occur after 4 and 6 months of age in App-KI mice, respectively [3], 18-month-old mice were considered to be sufficiently aged to ensure these occurrences. Actually, Aβ deposition was confirmed throughout the olfactory pathway of App-KI mice, that were obtained from the mice at the end of the behavioral experiments but was not observed in WT mice.
Odorants interact with ORNs in the OE, lining the interior of the nose. The axons arising from the ORNs directly project to neurons in the olfactory bulb, which in turn projects to the pyriform cortex. The olfactory tract also projects to several other targets in the forebrain, such as the olfactory tubercle, amygdala, and entorhinal cortex. Projections from the pyriform cortex and other regions of the forebrain provide olfactory information to several additional regions of the cerebral cortex [11]. OE consists of various cell types, including microvillar and basal cells, as well as ORNs, which can be identified by their higher expression of OMP [12].
Odorant signal transduction occurs in the olfactory cilia of the ORNs, as follows: odorants bind to odorant receptors, triggering the activation of G protein, in turn activating adenylate cyclase 3 to produce cyclic adenosine monophosphate (cAMP) [13]. cAMP binds to cyclic nucleotide-gated (CNG) channels, promoting their opening and allowing Na+ and Ca2 + ions to flow into the intracellular medium, depolarizing the cell. This transient increase in intracellular Ca2 + concentration opens Ca2 +-activated Cl (CAC) channels, which amplify the CNG channel signal [14]. The ensuing depolarization generates action potentials, which are then sent to the OB.
We used vanilla extract as the odor sample, which is an attractive odorant, and 2-ME, which is an aversive odorant. Although more than 1000 olfactory receptor genes have been found in the mouse genome [15], the agonist-receptor relationships have not been clarified in most studies; therefore, the olfactory receptors that respond to vanilla and 2-ME are unknown. The olfactory behavior of App-KI mice indicated that the total exploratory time to attractive odorants was longer than that for water, and the total exploratory time to aversive odorants was shorter than that for water. In other words, sensitivity to odorants in App-KI mice was not completely lost. In contrast, the exploratory behavior time of App-KI mice for the vanilla extract was significantly lower than that in WT mice, confirming that changes in olfactory behavior occur in App-KI mice. No obvious difference, however, was observed in olfactory behavior regarding 2-ME between the two genotypes. It is considered that there are odorants that are likely to change and those that are difficult to change caused by AD. In olfactory identification tests on patients with AD, Umeda-Kameyama et al. [16] reported that the accuracy of identification of odorants such as perfume and rose is significantly reduced, whereas that of other odorants such as sweaty sock and roasted garlic is not significantly changed. Furthermore, in our olfactory behavior test, the olfactory behavior is expressed as the difference in the exploratory time between the solvent and the odorant. Because the exploratory time for the solvent, namely water, is short, it may be difficult to observe the difference in the aversive odorants, which have a shorter exploratory time than the solvent.
Since olfactory impairment was observed in App-KI mice, the expression of olfactory-related molecules was investigated in the OE of mice. Odor detection ability relies largely on the primary olfactory pathway, with ORNs in the OE receiving odor signals, which are then relayed to the OB. Although it has been reported that OMP expression was not observed in AβPP/PS1 transgenic mice [17], an obvious OMP signal was observed in the mouse model used in this study, while no obvious morphological differences were observed in the OE. Furthermore, no significant differences were observed in the mRNA expression of representative olfactory-related genes, including the olfactory receptor Olfr15, olfactory-specific G protein Golf, CNG channel subunits Cnga2, Cnga4, Cngb1, CAC channel Tmem16B, and trace amine-associated receptors Taar2, Taar3, and Taar7f. Therefore, it appears that the difference in olfactory behavior was not caused by a decrease in the expression levels of these genes.
Some previous studies using animal models have suggested that apoptosis due to Aβ deposition in the OE may contribute to the deterioration of the sense of smell. In a transgenic mouse model that selectively overexpresses mutated humanized APP (hAPP) in either mature or immature ORNs, intracellular hAPP accumulation seemed to induce neuronal loss, indicating that the mechanisms involved in neuronal apoptosis might be cell autonomous [18, 19]. In WT mice, the nasal injection of an adenovirus expressing either Bri-Ab40 or Bri-Ab42 fusion proteins, which are cleaved sequentially by furin to secrete the human Aβ40 or Aβ42 peptide, respectively, did not increase apoptotic cells in the OE, but disrupted correct axonal targeting to the OB [20]. In contrast, Aβ deposition has been observed in the olfactory pathway of App-KI mice. Therefore, it is possible that the odor signal detected by the OE was not correctly transmitted, resulting in olfactory impairment. In Tg2576 transgenic mice, it has been suggested that olfactory impairment correlates with Aβ deposition in the olfactory pathway [21].
In this study, we investigated the relationship between olfactory impairment and the expression levels of olfactory-related genes in the OE. We propose that the expression levels of olfactory-related genes are irresponsible for the olfactory impairment caused by AD. We believe that the data gathered in this study will provide useful information with which to clarify the mechanisms of olfactory impairment caused by AD.
Footnotes
ACKNOWLEDGMENTS
This work was supported in part by a Grant-in-Aid for Scientific Research (B) 19H02905 to MN from the Japan Society for the Promotion of Science, and by the Adaptable and Seamless Technology Transfer Program through Target-driven R&D (A-STEP) from the Japan Science and Technology Agency (JPMJTR194F to MN and TM).