
Abstract
Background:
Until now, both cross-sectional and longitudinal studies have identified controversial findings about the association between daytime napping and Alzheimer’s disease (AD) or cognitive decline. Therefore, it remains unclear about the causal association between daytime napping and AD or cognitive decline.
Objective:
We aim to investigate the causal association between daytime napping and AD.
Methods:
Here, we conduct a bidirectional Mendelian randomization (MR) analysis to investigate the causal association between daytime napping and AD using large-scale GWAS datasets from daytime napping including 452,633 individuals of European ancestry and AD including 35,274 AD and 59,163 controls of European ancestry. A total of five MR methods are selected including inverse-variance weighted (IVW), weighted median, MR-Egger, MR-PRESSO, and contamination mixture method.
Results:
MR analysis highlights significant causal association of AD with daytime napping using IVW (beta = -0.006, 95% CI [–0.009, –0.002], p = 2.00E-03), but no significant causal association of daytime napping with AD using IVW (OR = 0.76, 95% CI 0.53-1.10, p = 1.40E-01).
Conclusion:
Our bidirectional MR analysis demonstrates the causal effect of AD on daytime napping. However, there is no causal effect of daytime napping on AD. Our current findings are consistent with recent evidence from other MR studies that highlight little evidence supporting a causal effect of sleep traits on AD and support the causal effect of AD on sleep traits.
INTRODUCTION
Daytime napping is generally reported in Alzheimer’s disease (AD) [1, 2]. It is reported that the prevalence of napping in elderly ranges from 20% to 60% [3]. 40% of the older adults who napped daily or snoozed for more than an hour on nap days were likely to develop AD [2]. However, the association between daytime napping and cognition remain conflicting, as reported in previous cross-sectional studies [2]. Some studies supported the benefits of napping on cognitive performance [4–7]. Cai and colleagues evaluated the association of afternoon napping with cognitive function in 2,214 elderly participants from the aging Chinese population [4]. They found that the napping group had better cognitive function [4]. Li and colleagues evaluated the cross-sectional association of self-reported post-lunch napping with cognitive function in 2,974 individuals and found significant association [5]. Using multiple regression analysis, they found that moderate napping (30–90 min) was associated with better cognition than non-napping (0 min), short napping (<30 min), and extended napping (>90 min), respectively [5].
In addition to these above benefits, some cross-sectional studies highlighted the adverse effects of napping on cognitive performance [8]. Owusu and colleagues conducted a cross-sectional study using 2,549 participants and identified napping to be a potential marker of cognitive decline, or a contributor to cognitive decline [8]. A Korean population-based cohort study in 2,501 participants showed that napping (30–60 min) was associated with worse cognitive function, and napping > 60 min was associated with better cognitive function [9]. A British population-based cohort study identified the daytime napping to be an increased risk of all-cause mortality [10].
Until recently, Li and colleagues consider that these inconsistent effects from the cross-sectional studies may result from the one single nap assessment for each participant [2]. Li and colleagues have evaluated the longitudinal association between daytime napping and AD using the data from 1,401 participants of the Rush Memory and Aging Project during up to 14-year follow-up [2]. They found that older adults had longer and more frequent daytime napping with aging, especially AD with more than doubling the annual increases in daytime napping duration or frequency [2]. Conversely, longer and more frequent daytime napping was associated with the increased risk of AD [2]. Meanwhile, Li and colleagues found similar findings about the association between daytime napping and cognitive decline [2]. Collectively, Li and colleagues have identified a potential bidirectional association between daytime napping and AD or cognitive decline, and may improve our understanding of the shared common pathophysiological mechanisms [2].
However, other longitudinal studies have highlighted the protective effects of daytime napping on cognitive decline, which do not support the findings from Li and colleagues [2]. Keage and colleagues conducted a longitudinal, population-based study in 2012 cognitively normal individuals aged≥65 years from the MRC Cognitive Function and Ageing Study (CFAS) [6]. They identified daytime napping to be associated with the decreased risk of cognitive decline in 2 and 10 years [6]. Kitamura and colleagues investigated the longitudinal association between daytime napping and cognitive decline using the data from 389 community-dwelling individuals aged≥65 years during up to 5-year follow-up and found that daytime napping reduced the risk of cognitive decline [11]. Therefore, it is urgent to establish the causal association between daytime napping and AD or cognitive decline, as described by Li and colleagues in their discussion [2].
Until now, genome-wide association study (GWAS) of daytime napping [12] and AD [13] are now publicly available and provide strong support to evaluate the causal association using Mendelian randomization (MR) [14, 15]. Here, we conduct a bidirectional MR analysis to evaluate the causal link between daytime napping and AD using large-scale GWAS datasets.
METHODS
Study design
We perform a bidirectional MR study using large-scale GWAS summary datasets from daytime napping [12] and AD [13]. MR consists of three assumptions: 1) genetic variants as the potential instrumental variables associate with the exposure (such as daytime napping) [16]; 2) genetic variants are not associated with the confounders of an outcome (such as AD) [16]; 3) genetic variants cause the outcome (AD) only through the exposure (daytime napping) [16]. The second and third assumptions are defined to be the independence from pleiotropy. Recent MR studies have provided more detailed information about the study design [14, 16–20]. Figure 1 provides the data sources and the MR design.

The flow chart about the MR study design. GWAS, genome-wide association studies; IVW, Inverse-variance weighted.
Daytime napping GWAS dataset and genetic variants
The daytime napping GWAS is from the UK Biobank including 452,633 individuals of European ancestry [12]. 255,746 (59% females), 172,897 (50% females), and 23,990 (33.9% females) participants reported never/rarely, sometimes, and always napping, respectively [12]. This GWAS identified 123 independent genetic variants to be associated with daytime napping (p < 5.00E-08), which we selected as the potential instruments, as provided in Supplementary Table 1 [12].
AD GWAS dataset and genetic variants
The AD GWAS dataset is from the International Genomics of Alzheimer’s Project (IGAP) including 94,437 individuals (35,274 clinically diagnosed AD and 59,163 controls) of European ancestry [13]. AD was diagnosed in participants with dementia meeting clinical criteria for probable or possible disease established by the National Institute of Neurological and Communication Disorders and Stroke and the Alzheimer Disease and Related Disorders Association (NINCDS-ADRDA) criteria [13]. IGAP AD GWAS in stage 1 (21,982 AD and 41,944 controls) is publicly available [13]. IGAP identified 26 and 8 independent AD genetic variants with the genome-wide significance p < 5.00E-08 and suggestive association p < 5.00E-07, respectively. Here, we selected these 34 AD genetic variants as the potential instruments. Table 1 and Supplementary Table 2 provide the demographic profile about the AD GWAS, and the detailed information about 34 AD genetic variants, respectively.
The demographic profile about the AD GWAS dataset
AD, Alzheimer’s disease; AAO, age at onset; AAE, age at examination; ADGC, Alzheimer Disease Genetics Consortium; CHARGE, Cohorts for Heart and Aging Research in Genomic Epidemiology Consortium; EADI, European Alzheimer’s Disease Initiative; GERAD, Genetic and Environmental Risk in Alzheimer’s Disease consortium [13].
Establishing the Wald estimator
Using the 123 independent daytime napping genetic variants, we extracted their corresponding summary results in AD GWAS dataset. Similarly, using the 34 AD genetic variants, we extracted their corresponding summary results in daytime napping GWAS dataset. For the same effect allele from each daytime napping genetic variant G
j
, we assume that their corresponding summary results in daytime napping GWAS dataset to be the beta coefficients and their standard errors (
MR analysis
Multiple MR methods were used to combine the single Wald estimator from each genetic variant to get the overall estimate. These methods include inverse-variance weighted (IVW) [21], MR-Egger [22], weighted median [21], MR-PRESSO (Mendelian Randomization Pleiotropy RESidual Sum and Outlier) [23], and contamination mixture method [24]. IVW assumes no horizontal pleiotropy and all genetic variants are valid instrumental variables [21, 24]. IVW is a main and popular MR approach, which had been used in recent MR studies [25–30]. MR-Egger could test the directional pleiotropy using MR-Egger intercept test, and correct for the presence of pleiotropy [22]. If the intercept is 0, MR-Egger is the same as IVW [22]. The simple median assumes that at least 50% of genetic variants are valid instrumental variables [21]. Weighted median assumes that at least 50% of the weight is from the valid instrumental variables [21, 31]. MR-PRESSO tests the pleiotropy and identify the horizontal pleiotropic outliers [23, 32]. Contamination mixture method is newly developed method for MR analysis with hundreds of genetic variants [24]. It performs MR robustly and efficiently even there are invalid instrumental variables [24]. All statistical tests were calculated using ‘MendelianRandomization’ [32] and ‘MR-PRESSO’ [23] with the significance threshold p < 0.05.
RESULTS
MR analysis show no significant causal association of daytime napping with AD using IVW (OR = 0.76, 95% CI 0.53–1.10, p = 1.40E-01), weighted median (OR = 0.68, 95% CI 0.41–1.14, p = 1.40E-01), MR-Egger (OR = 1.01, 95% CI 0.27–3.78, p = 9.84E-01), and MR-PRESSO (OR = 0.76, 95% CI 0.53–1.10, p = 1.40E-01), as provided in Table 2. Interestingly, MR analysis highlights significant causal association of AD with daytime napping using IVW (beta = –0.006, 95% CI [–0.009, –0.002], p = 2.00E-03), weighted median (beta = –0.006, 95% CI [–0.009, –0.006], p = 1.97E-09), MR-Egger (beta = –0.006, 95% CI [–0.01, –0.002], p = 6.00E-03), and MR-PRESSO (beta = –0.006, 95% CI [–0.009, –0.002], p = 3.36E-03). Importantly, all these estimates are consistent in terms of direction and magnitude, as provided in Table 2. Meanwhile, MR-Egger intercept test and MR-PRESSO global test found no evidence of pleiotropy or horizontal pleiotropic outliers (Table 2). The individual causal estimates about the causal association of AD with daytime napping using IVW are provided in Fig. 2.
MR results about the causal association between daytime napping and AD
OR, odds ratio; CI, confidence interval; IVW, Inverse-variance weighted meta-analysis; MR-PRESSO, Mendelian Randomization Pleiotropy RESidual Sum and Outlier.
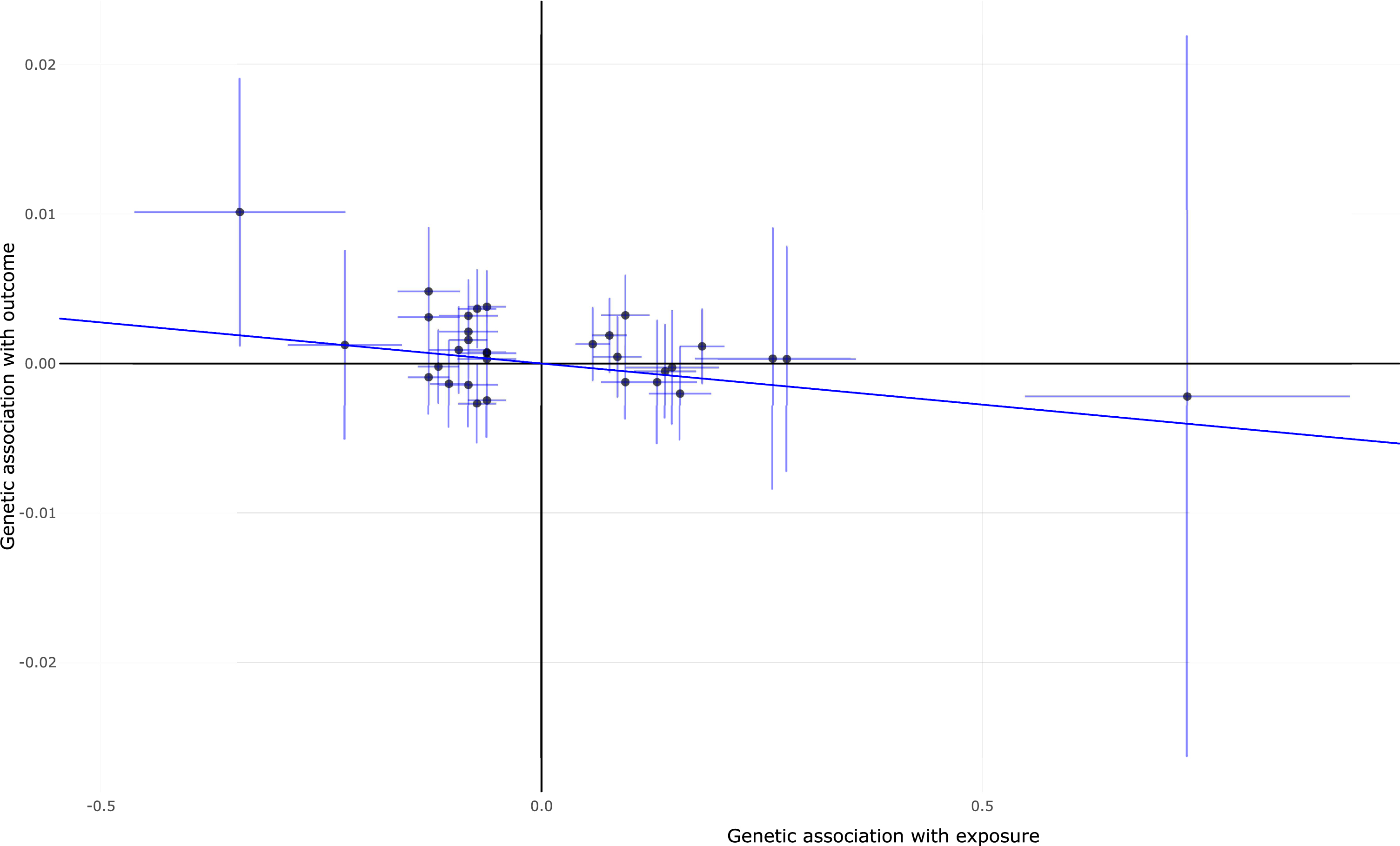
Individual estimates about the causal effect of AD on daytime napping using IVW method. The x-axis shows the effect and standard error for single genetic variant on AD and the y-axis shows the effect and standard error for single genetic variant on daytime napping. IVW, inverse-variance weighted.
DISCUSSION
Until now, both the cross-sectional studies and longitudinal studies have identified controversial findings about the association between daytime napping and AD or cognitive decline [2, 11]. Here, we conduct a bidirectional MR analysis to investigate the causal association between daytime napping and AD using two large-scale GWAS datasets from daytime napping (n = 452,633) and AD (n = 94,437). We highlighted the causal association of AD with reduced daytime napping, but no causal association of daytime napping with AD. Our findings may provide additional useful information about the bidirectional longitudinal relationship between daytime napping and AD [2]. To our knowledge, this is the first bidirectional MR analysis to examine the causal association between daytime napping and AD.
Until now, multiple MR studies have investigated the causal effect of sleep traits on AD. Our findings are comparable with these MR findings [33–36]. Anderson and colleagues conducted a MR study to evaluate the causal association of multiple sleep traits with AD [33]. However, they found little evidence supporting a causal effect of sleep traits on AD [33]. Take daytime napping for example, Anderson and colleagues found no causal effect of daytime napping on AD using IVW (OR = 0.67, 95% CI 0.45–1.02, p = 0.06), weighted median (OR = 0.74, 95% CI 0.42–1.29, p = 0.28), MR-Egger (OR = 0.95, 95% CI 0.22–4.20, p = 0.95) [33]. Cullell and colleagues evaluated the causal effect of 12 sleep traits including daytime sleepiness on AD, and only identified the sleep efficiency to be significantly associated with reduced risk of AD using IVW (p = 0.036) [34]. Andrews and colleagues found no causal effect of two sleep traits including insomnia symptoms and sleep duration on AD [35]. Huang and colleagues did not find evidence to support the causal effect of six sleep traits on AD [36].
Meanwhile, Huang and colleagues also investigated the causal effect of AD on six sleep traits [36]. They found that AD was significantly associated with shorter sleep duration, reduced self-reported long sleep, a smaller number of sleep episodes, and lower risk of insomnia [36]. Therefore, our findings are comparable with these MR findings [36]. We consider that our current study may have several strengths. First, large-scale daytime napping [12] and AD [13] GWAS datasets contribute to provide ample power. Second, all the participants in both GWAS datasets are of the European descent and contribute to reduce the bias caused by population stratification. Third, the pleiotropy analysis methods MR-Egger intercept test and MR-PRESSO test ensure the MR assumptions. Fourth, five MR methods contribute to ensure the precision.
Meanwhile, our current study may have several limitations. First, the pleiotropy analysis could not completely be excluded using MR-Egger intercept test and MR-PRESSO test [37, 38]. Second, we limit our MR analysis in individuals of European ancestry. However, the causal association may vary in different ancestries. Third, we only selected 34 AD genetic variants from the IGAP (35,274 AD and 59,163 controls) as the potential instruments [13]. In fact, a recent two-stage GWAS identified 75 AD genetic variants with the genome-wide significance p < 5.00E-08 [39]. However, this large-scale dataset is based on 111,326 clinically diagnosed or proxy AD cases and 677,663 controls [39]. Our recent findings and previous studies have showed significant difference and genetic heterogeneity in clinically diagnosed AD GWAS and self-report proxy phenotype GWAX [14, 40]. Therefore, we did not select these 75 AD genetic variants as the potential instruments. Fourth, we just performed the bidirectional MR analysis to investigate the causal association between daytime napping and AD, but not the causal association between daytime napping and cognitive decline or cognitive function. Fifth, we could not analyze the original genotype and phenotype data using advanced AI/ML methods to come into definitive conclusion, as only the GWAS summary results are publicly available. Therefore, future studies are required to further verify our findings.
Taken together, we conducted a bidirectional MR analysis to evaluate the causal association between daytime napping and AD, and only demonstrated the causal effect of AD with daytime napping. Our current findings are consistent with recent evidence from other MR studies that highlight little evidence supporting a causal effect of sleep traits on AD and support the causal effect of AD on sleep traits.
Footnotes
ACKNOWLEDGMENTS
We thank the International Genomics of Alzheimer’s Project (IGAP) and Sleep Disorder Knowledge Portal for providing the GWAS summary results. This work was supported by funding from the National Natural Science Foundation of China (Grant No. 81901181, 82071212, and 12026414), Beijing Natural Science Foundation (Grant No. JQ21022), Beijing Ten Thousand Talents Project (Grant No. 2020A15), and the Natural Science Foundation of Shandong Province (No. ZR2020MH141).